赤霉素与小麦生理型雄性不育的关系
2017-04-24巴青松张改生李桂萍张根生周丽娟宋运贤傅兆麟
巴青松,张改生,李桂萍,张根生,周丽娟,宋运贤,陈 楚,傅兆麟
(1.淮北师范大学生命科学学院/资源植物生物学安徽省重点实验室,安徽淮北 235000;s2.西北农林科技大学农学院/陕西省作物杂种优势研究与利用重点实验室,陕西杨凌712100)
赤霉素与小麦生理型雄性不育的关系
巴青松1,张改生2,李桂萍1,张根生1,周丽娟1,宋运贤1,陈 楚1,傅兆麟1
(1.淮北师范大学生命科学学院/资源植物生物学安徽省重点实验室,安徽淮北 235000;s2.西北农林科技大学农学院/陕西省作物杂种优势研究与利用重点实验室,陕西杨凌712100)
为探究赤霉素与小麦生理型雄性不育的关系,以新型小麦化学杀雄剂SQ-1 作为诱导剂,西农1376为材料,构建了西农1376 不育和可育生理系,通过扫描电镜、荧光定量PCR、气相质谱等技术,研究了小孢子花粉形态、TaGAMYB和 CYP709C1基因表达、硬脂酸和赤霉素含量和花粉育性。结果表明,化学杀雄剂SQ-1 诱导的生理型雄性不育系花粉粒畸形,TaGAMYB和 CYP709C1基因表达量显著下降,硬脂酸含量显著上升,赤霉素含量显著低于可育系。赤霉素对雄性不育系的育性有一定逆转效应。
小麦;生理型雄性不育;赤霉素;花粉壁
大量研究表明,植物雄性不育系的内源激素含量与保持系显著不同[1-4]。王华忠等[5]研究认为,油菜细胞质不育系的赤霉素含量显著低于保持系。李英贤[6]研究发现,小麦K型细胞质雄性不育与花药组织赤霉素含量有密切关系。赤霉素的生理功能主要是依靠赤霉素在植物体内的信号转导来实现。GAMYB(Gibberellic acid v-myb avian myeloblastosis viral oncogene homolog)是受GA(Gibberellic acid)诱导的MYB类转录因子,GAMYB蛋白通过结合到GA应答基因的启动子上来诱导下游基因的表达,在植物种子萌发、茎的伸长、开花和花药的发育过程中发挥着极其重要的作用[7]。Fiona等[8]研究发现,随着大麦HvGAMYB基因表达量的上升,花药长度变小,这种现象和过量施用GA的植株表型相似;转基因大麦中,HvGAMYB蛋白比野生大麦高4倍以上可导致雄性不育现象。Zhang等[9]研究认为,黄瓜的CsGAMYB基因只在花药中表达,调控花粉育性。Kaneko等[10]发现水稻OsGAMYB基因缺失可导致花药和花粉发育异常。OsGAMYB能够通过调控脂肪酸羟基化影响花粉壁的形成[11]。由此可见,花药中GAMYB基因能够调控脂肪酸代谢,进而影响花粉壁的形成。
TaGAMYB(AY615200.1)是小麦赤霉素诱导的GAMYB转录因子,此基因异常表达导致小麦育性大幅下降[12]。 CYP709C1是目前发现的小麦中唯一催化18C脂肪酸羟基化酶基因,在小麦花粉发育中,可能受TaGAMYB转录因子调控,能够合成花粉壁前体[13]。然而,赤霉素诱导的TaGAMYB基因在小麦生理型雄性不育中的作用还少见报道。目前,对于赤霉素在雄性不育中的作用,主要集中于不同作物遗传型雄性不育小孢子发育过程中赤霉素的定量分析,对于赤霉素是否参与生理型雄性不育小麦育性转换的调控,以及如何参与调控育性转换都是值得研究的问题。由于单核期是SQ-1诱导的生理型雄性不育的关键时期[14],本研究拟测定不育系单核期赤霉素含量、TaGAMYB和 CYP709C1基因表达水平和脂肪酸含量,分析外源赤霉素对生理型雄性不育小麦育性的影响,以期确定小麦生理型雄性不育与赤霉素的关系,为揭示雄性不育的表达调控机理积累资料。
1 材料与方法
1.1 材料与设计
供试材料为小麦品种西农1376。当西农1376 生长发育至Feeke’s 8.0~8.5 时期,将其分为两组,一组喷施化杀剂SQ-1(5.0 kg·hm-2),诱导产生生理型不育系,简称为1376-CIMS,另一组喷施等量清水为对照。通过DAPI鉴定,在1376-CIMS和西农1376花药发育到单核期进行花药取样,然后迅速用液氮冷冻,并保存于-80 ℃冰箱中。在成熟期通过淀粉碘化钾实验检测其育性,同时,通过扫描电镜观察花粉形态。
1.2 方 法
1.2.1 小麦花药总RNA提取、cDNA合成、TaGAMYB和 CYP709C1基因表达量测定
称取约100 mg花药,用TRNzol-A+试剂(北京天根生化科技有限公司)按说明书方法提取样品总RNA,经核酸蛋白检测仪和电泳检测RNA浓度和质量。用Prim Script RT Reagent Kit 试剂盒(TaKaRa,大连)合成第一链cDNA,按照产品说明书进行反转录试验。根据TaGAMYB(AY615200.1)和 CYP709C1基因的cDNA序列,在其保守区用Primer Premier 5.0软件设计引物。TaGAMYB,E1-F1:5′-AATGGCAC CTTCTCTACTT-3′,E1-R1:5′-GGCGACACA CACTCCGACT-3′; CYP709C1,E1-F1:CGGAG CCAAGCCGACACT,E1-R1:GGCTGCTCAGG TCAATCT。内参为小麦18S rRNA基因,其引物为:18S-F:5′-CGTCCCTGCCCTTTGTACAC-3′,18S-R:5′-AACACTTCACCGGACCATTCA-3′。以花药的cDNA 为模板,小麦18S rRNA 基因为内参进行定量PCR分析。3 次重复。PCR 反应体系20 μL :2 μL cDNA模板,10 μmol·L-1正反向引物各0.5 μL,10 μL SYBR qPCR Mix,用ddH2O 补至20 μL。反应条件为95 ℃ 30 s;95 ℃ 5 s,52 ℃ 30 s,72 ℃ 50 s, 32个循环;72 ℃,10 min。每个样品的相对表达量以目的基因的表达量(目的基因/18S rRNA)除以单核期可育系的表达量(目的基因/18S rRNA)来表示,单核期可育系的相对表达量规定为1。
1.2.2 花药脂肪酸含量的测定
气相色谱条件:RXi-5ms毛细管色谱柱(30 m×0.25 mm,0.25 μm),载气为氦气;程序升温:柱初始温度180 ℃,保温2 min;以5 ℃·min-1速率升至280 ℃,保温5 min;进样口温度280 ℃,柱流量1 mL·min-1,分流比100∶1;进样量1 μL。质谱条件:四极质量选择检测器 GCMS-QP 2010,电子轰击离子源(EI),电子能量70eV,电子倍增器电压1.22 kV,质量扫描范围(m/z)为50至700,离子源温度200 ℃,GC-MS接口温度230 ℃,溶剂峰切除时间2 min,质谱检测起测时间2 min。质谱图计算机检索数据库NIST147。
1.2.3 花药赤霉素含量的测定
参照王宝勤等[15]的方法进行赤霉素的提取和测定。
1.2.4 赤霉素对小麦育性转换的影响
矮壮素是赤霉素合成的抑制剂[16],在小孢子发育到雌雄蕊原基分化期时,对西农1376进行矮壮素处理,处理浓度为 100 μmoL·L-1;同时,对1376-CIMS进行赤霉素处理,处理浓度为0.8 mg·L-1。处理方法采用叶片涂抹法,每个处理涂抹 30 株,处理后挂牌标记。待抽穗后,套袋调查自交结实率。
2 结果与分析
2.1 小麦TaGAMYB和 CYP709C1基因的表达
通过定量PCR分析发现,TaGAMYB和 CYP709C1基因在花药单核期,在1376-CIMS中的相对表达量显著低于西农1376(图1)。
2.2 小麦花药脂肪酸和赤霉素含量
气相质谱分析结果表明,单核期1376-CIMS的硬脂酸含量显著高于西农1376(图3A),表明1376-CIMS单核期的脂肪酸转化受阻,引起花粉壁畸形(图2A),导致雄性不育(图2B)。
单核期1376-CIMS花药中赤霉素的含量显著低于西农1376,只有对照株含量的 75.2%(图3B)。
2.3 赤霉素和矮壮素对小麦生理型不育系育性的调控
在雌雄蕊分化期用赤霉素和矮壮素处理小麦,结果发现,矮壮素处理1376后,其花药中赤霉素含量显著降低(P<0.05),结实率为76%,育性降低。赤霉素处理西农1376-CIMS后,花药中赤霉素含量提高,其结实率可达12%。试验结果证明,赤霉素信号途径参与了小麦育性的转换,赤霉素在花粉发育过程中发挥着重要作用。
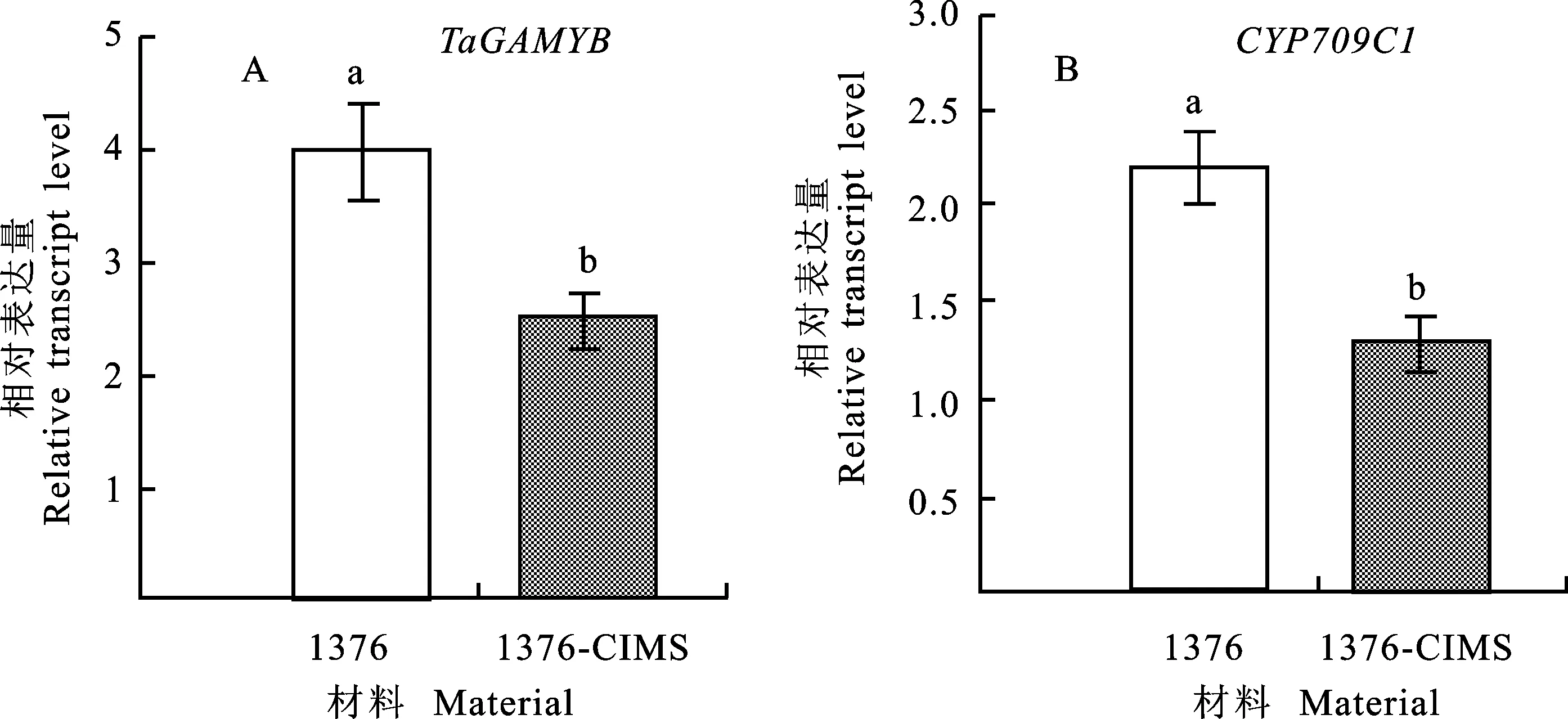
1376:西农1376。图柱上不同字母表示处理间差异在0.05水平显著。下同。
1376:Xinong 1376.Different letters following data mean significant difference between treatments at 0.05 level. The same below.
图1TaGAMYB和 CYP709C1基因在小孢子单核期的相对表达量
Fig.1 Relative expression ofTaGAMYBand CYP709C1 at single nucleus stage of microspore
3 讨 论
本试验中,可育系花药的GA含量显著高于不育系,结果与前人研究相同[17-18]。大麦中的GAMYB受赤霉素诱导,在花药发育、雄蕊生长和花原基诱导等植物生长发育中发挥重要作用[10]。GAMYB表达量降低,引起下游调控育性基因 TDR、CYP703A3、CYP704B2表达量下调,导致绒毡层异常伸长,花粉壁形成受阻,证明GAMYB在水稻花药小孢子发育和花粉壁形成中起关键作用[19-20]。本试验中,不育系的TaGAMYB和 CYP709C1基因表达量显著低于对照,引起脂肪酸积累。这是因为不育系中较低的GA 含量,使GAMYB的活性降低,导致脂肪酸转化受阻[12],从而严重影响花粉壁的正常形成。
关于外源施用赤霉素对雄性育性的转变作用也有许多报道。张建奎等[21]研究认为,外源赤霉素能够提高小麦温光敏核不育系C49S的育性。Sakata等[22]研究发现,外施赤霉素能够显著提高水稻低温诱导的雄性不育系的育性。但过高浓度的赤霉素处理也会引起植物雄性不育[23]。本研究发现,赤霉素合成抑制剂-矮壮素可以降低小麦花粉的可育性,外施赤霉素对小麦生理型雄性不育系的育性有一定的“拯救效应”。这些研究结果说明赤霉素对小麦的育性有一定调控效应,推测其作用机制中可能存在着一种剂量依赖效应。
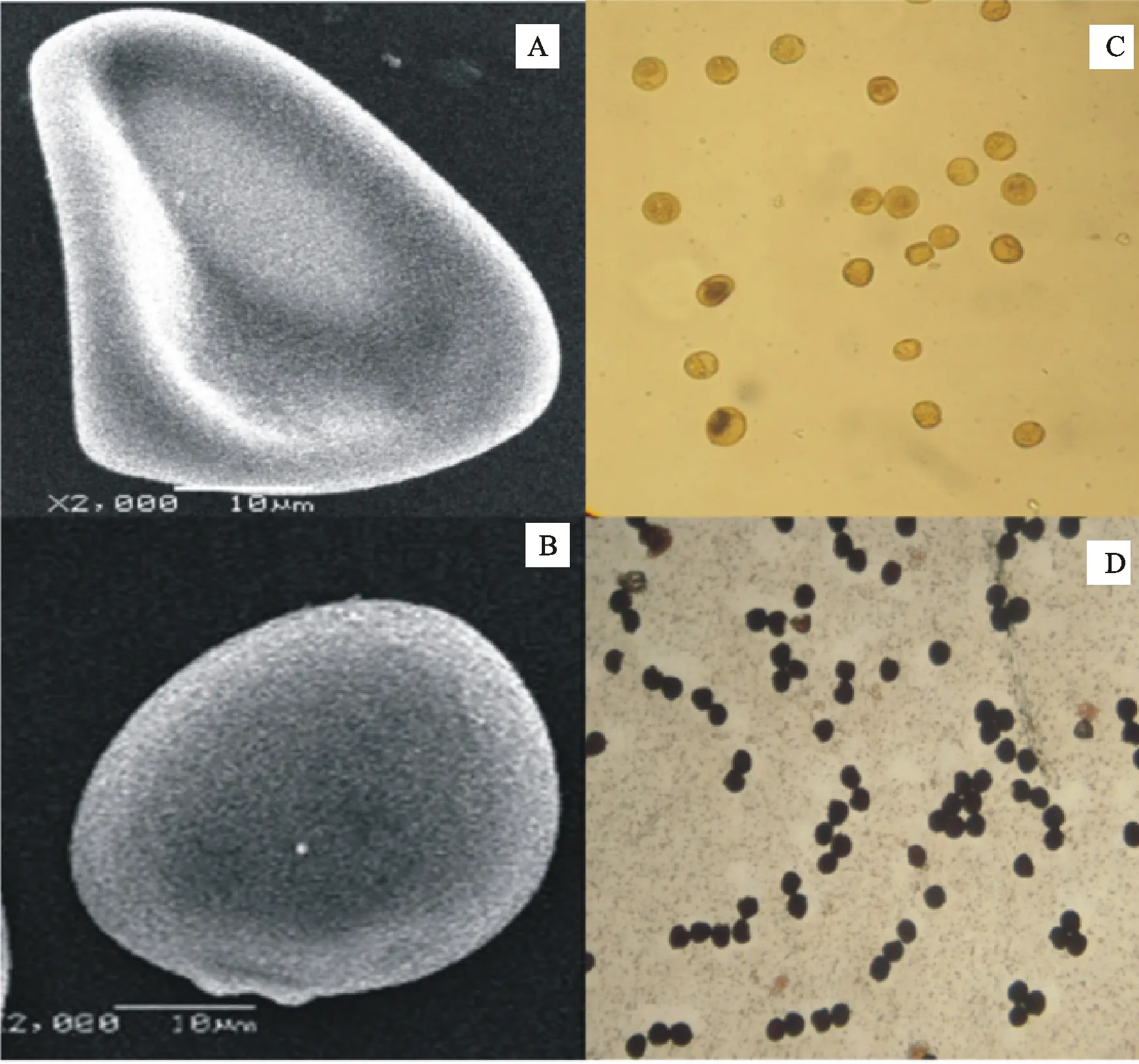
A:不育花粉; B:可育花粉;C:不育花粉淀粉碘染色;D:可育花粉淀粉碘染色。
A:Sterile pollen; B:Fertile pollen; C:Sterile pollen stained with KI; D:Fertile pollen stained with KI.
图2 花粉形态和碘化钾染色
Fig.2 Pollen morphology and potassium iodide dyeing test

图3 西农1376和1376-CIMS的硬脂酸(A)和赤霉素(B)含量
[1] ZHANG J K,ZONG X F,YU G D,etal.Relationship between phytohormones and male sterility in thermo-photo-sensitive genic male sterile(TGMS) wheat [J].Euphytica,2006,150(1):241.
[2] SHUKLA A,SAWHNEY V K.Cytokinins in a genic male sterile line of Brassica napus [J].PhysiologiaPlantarum,2006,85(1):23.
[3]SAWHNEY V K,SHUKLA A.Male sterility in flowering plants:Are plant growth substances involved? [J].AmericanJournalofBotany,1994,81(12):1640.
[4] 张爱民,李英贤,黄铁城.小麦雄性不育与内源激素关系的初步研究[J].农业生物技术学报,1996(1):56.
ZHANG A M,LI Y M,HUANG T C.Relations between endogenous hormones and male sterility in wheat(Triticumaestivum):Preliminary results [J].JournalofAgriculturalBiotechnology,1996(1):56.
[5] 王华忠,吴则东,韩 英,等.甜菜细胞质雄性不育与内源激素含量的关系[J].中国农业科学,2008,41(4):1134-1141.
WANG H Z,WU Z D,HAN Y,etal.Relationships between endogenous hormone contents and cytoplasmic male sterility in sugar beet [J].ScientiaAgriculturaSinica,2008,41(4):1134.
[6] 李英贤.小麦细胞质雄性不育与花药组织内源激素的关系[J].农业生物技术学报,1996(4):307.
LI Y X.Relationship between wheat cytoplasmic male sterility and the content of endogenous hormones in the anther [J].JournalofAgriculturalBiotechnology,1996(4):307.
[7] WOODGER F J,MILLAR A,MURRAY F,etal.The role of GAMYB transcription factors in GA-regulated gene expression [J].JournalofPlantGrowthRegulation,2003,22(2):176.
[8] MURRAY F,KALLA R,JACOBSEN J,etal.A role forHvGAMYBin anther development [J].PlantJournalforCell&MolecularBiology,2003,33(3):481.
[9] ZHANG Y,ZHANG X,LIU B,etal.AGAMYBhomologueCsGAMYB1 regulates sex expression of cucumber via an ethylene-independent pathway [J].JournalofExperimentalBotany,2014,65(12):3201.
[10] KANEKO M,INUKAI Y,UEGUCHI-TANAKA M,etal.Loss-of-function mutations of the riceGAMYBgene impair alpha-amylase expression in aleurone and flower development [J].PlantCell,2004,16(1):33.
[11] AYA K,MATSUOKA M.Gibberellin modulates anther development in rice via the transcriptional regulation of GAMYB [J].PlantCell,2009,21(5):1453.
[12] WANG Y,SUN F,CAO H,etal.TamiR159 directed wheatTaGAMYBcleavage and its involvement in anther development and heat response [J].PlosOne,2012,7(11):1.
[13]KANDEL S,MORANT M,BENVENISTE I,etal.Cloning,functional expression,and characterization of CYP709C1,the first sub-terminal hydroxylase of long chain fatty acid in plants.Induction by chemicals and methyl jasmonate [J].JournalofBiologicalChemistry,2005,280(43):35881.
[14] 盛 英,张改生,李亚鑫,等.小麦生理型雄性不育花药绒毡层和孢粉素变化与 RAFTIN1表达的关系[J].中国农业科学,2011,44(19):3937.
SHENG Y,ZHANG G S,LI Y X,etal.The relationship on anther tapetum,sporopollenin and expression of RAFTIN1 in physiological male sterile wheat [J].ScientiaAgriculturaSinica,2011,44(19):3937.
[15] 王保勤,牛吉山.一种小麦籽粒赤霉素含量测定方法的建立[J].中国农学通报,2007,23(1):334.
WANG B Q,NIU J S.A determination method for wheat grain gibberellins [J].ChineseAgriculturalScienceBulletin,2007,23(1):334.
[16] 黄 涛,董高峰,李耿光,等.赤霉素及其合成抑制剂对沙田柚上胚轴离体再生的影响[J].植物生理学报,2002,38(6):559.
HUANG T,DONG G F,LI G G,etal.Effects of gibberellic acid and its synthetic inhibitors on shoots regeneration from epicotyl ofCitrusgrandis[J].PlantPhysiologyCommunications,2002,38(6):559.
[17] 杨海燕,宗学凤,余国东,等.激素调控对小麦温光敏核不育系育性转换的影响 [J].西南大学学报(自然科学版),2006,28(03):369.
YANG H Y,ZONG X F,YU G D,etal.Regulating effects of hormones on fertility alternation in thermo-photo-sensitive genic male sterile wheat [J].JournalofSouthwestAgriculturalUniversity(NaturalScience),2006,28(3):369.
[18] 张艳玉,张卫东,高庆荣,等.温光敏雄性不育小麦BNS幼穗发育中的内源激素变化 [J].西北植物学报,2013,33(6):1165-1170.
ZHANG Y Y,ZHANG W D,GAO Q R,etal.Dynamic changes of endogenous hormones in thermo-photo-sensitive male sterile wheat line BNS [J].ActaBotanicaBoreali-OccidentaliaSinica,2013,33(6):1165.
[19] 杨正福,张迎信,孙廉平,等.水稻雄性不育突变体gamyb5的鉴定与基因定位 [J].中国水稻科学,2016,30(02):143.
YANG Z F,ZHANG Y X,SUN L P,etal.Identification and gene mapping of male sterile mutant gamyb5 in rice [J].2016,30(02):143.
[20] MORANT M,JQRGENSEN K,SCHALLER H,etal.CYP703 is an ancient cytochrome P450 in land plants catalyzing in-chain hydroxylation of lauric acid to provide building blocks for sporopollenin synthesis in pollen [J].PlantCell,2007,19(5):1473.
[21] 张建奎,冯 丽,罗 丹,等.外源化学物质对小麦温光敏核不育两用系C49S育性的影响 [J].麦类作物学报,2001,21(1):69.
ZHANG J K,FENG L,LUO D,etal.Effect on the fertility of thermo-photo-sensitive genic male-sterile wheat C49S using different chemcals [J].JournalofTriticeaeCrops,2000,21(1):69.
[22] SAKATA T,ODA S,TSUNAGA Y,etal.Reduction of gibberellin by low temperature disrupts pollen development in rice [J].PlantPhysiology,2014,164(4):2011.
[23] COLOMBO N,FAVRET E A.The effect of gibberellic acid on male fertility in bread wheat [J].Euphytica,1995,91(3):297.
Relationship between Gibberellin and Chemical-Induced Male Sterility in Wheat
BA Qingsong1,ZHANG Gaisheng2,LI Guiping1,ZHANG Gensheng1,ZHOU Lijuan1,SONG Yunxian1,CHEN Chu1,FU Zhaolin1
(1.Anhui Key Laboratory of Plant Resources and Biology/College of Life Science,Huaibei Normal University,Huaibei,Anhui 235000,China; 2.Key Laboratory of Crop Heterosis of Shaanxi Province/College of Agronomy,Northwest A&F University,Yangling,Shaanxi 712100,China)
The study was conducted in order to reveal the relationship between gibberellin and chemical-induced male sterility in wheat(TriticumaestivumL). Wheat variety Xinong 1376 was explored to develop physiological male sterile line through spraying SQ-1(a new chemical agent) on wheat plants at Feeke’s 8.5 stage,with the plants sprayed with same volume water as the control. Pollen morphology,TaGAMYBand CYP709C1 expression,octadecanoic acid content,and gibberellin content were analyzed via techniques such as scanning electron microscope,fluorescence quantitative PCR,gas chromatography mass spectrum at mononuclear stage. The results indicated that pollen deformity of chemical-induced male sterility generated,and expression patterns ofTaGAMYBand CYP709C1 were down-regulated significantly,and instead,octadecanoic acid content rise sharply. Meanwhile,gibberellin content was significantly lower than that of fertile line.Gibberellin have a reverse effect on sterility of male sterile lines.
Wheat; Chemical-induced male sterility; Gibberellin; Pollen wall
时间:2017-03-07
2016-08-05
2016-09-0
安徽省自然科学基金项目(1708085QC62); 安徽省高等学校省级自然科学研究项目(KJ2014B19,KJ2016A624,KJ2013A232);淮北师范大学博士科研启动基金项目(15600971,12601033)
E-mail:baqs1234@163.com
S512.1;S330
A
1009-1041(2017)03-0344-05
网络出版地址:http://kns.cnki.net/kcms/detail/61.1359.S.20170307.1637.020.html
