Sema4D表达在慢性病毒持续感染中的作用机制研究进展
2017-04-17张久聪郑晓凤杨永林汪泳
张久聪,郑晓凤,杨永林,汪泳
1.兰州军区 兰州总医院消化科,甘肃 兰州 730050;2.兰州大学 第二医院消化科,甘肃 兰州 730030
Sema4D表达在慢性病毒持续感染中的作用机制研究进展
张久聪1,郑晓凤2,杨永林1,汪泳1
1.兰州军区 兰州总医院消化科,甘肃 兰州 730050;2.兰州大学 第二医院消化科,甘肃 兰州 730030
Sema4D又称CD100是近年新发现的重要的免疫调节分子,Sema4D与其受体plexin-B1和CD72结合,通过多种信号转导途径,在神经系统的轴突导向,肿瘤的侵袭转移和免疫系统中T、B细胞的活化和免疫调节中发挥关键作用。最新研究表明,Sema4D在慢性病毒持续感染中发挥重要作用,Sema4D对维持CD8+T细胞的功能尤为重要,Sema4D对控制感染慢性化至关重要。Sema4D作为监测T细胞免疫功能的关键标志分子,可能参与慢性病毒感染性疾病T细胞功能低下。我们就Sema4D表达在慢性病毒持续感染中的作用机制方面的研究进展做简要综述。
Sema4D;慢性病毒感染;免疫调节;研究进展
Semaphorins家族是一类以结构中具有Sema区域为共同特征的蛋白,最初作为轴突导向因子影响中枢神经系统的发育而被发现。近年发现,Semaphorins家族在广泛的器官系统中起着形态发生和内环境稳态的关键调控作用[1]。Semapho⁃rin 4D(Sema4D,又称CD100)与其受体plexin-B1和CD72结合,通过多种信号转导途径,在神经系统的轴突导向,肿瘤的侵袭转移和免疫系统中T、B细胞的活化和免疫调节中发挥关键作用[2-3]。最新研究表明,Sema4D在慢性病毒持续感染中发挥重要作用,Sema4D对维持CD8+T细胞的功能尤为重要,Sema4D对控制感染慢性化至关重要。Se⁃ma4D作为监测T细胞免疫功能的关键标志分子,可能参与慢性病毒感染性疾病T细胞功能低下。以下就Sema4D表达在慢性病毒持续感染中的作用机制方面的最新研究进展做简要综述。
1 Sema4D及其受体
Sema4D是一个相对分子质量为150×103的跨膜蛋白。其分子结构自N端始依次为Sema结构域、半胱氨酸富集区、免疫球蛋白样结构域、富赖氨酸延长区、疏水跨膜区及胞液尾[2-3]。在人类和小鼠,Sema4D都以2种形式存在,即膜结合型和可溶型。单核细胞表面的Sema4D与Sema4D单克隆抗体交联后可诱导膜结合型Sema4D被蛋白酶酶解成一个相对分子质量为120×103的保留有生物活性的可溶性Sema4D,另外活化的T、B淋巴细胞也可以释放可溶性Sema4D。Sema4D分布较广,在人类多种组织中都有表达,包括淋巴器官(脾脏、淋巴结和胸腺)和非淋巴器官(脑、肾和心脏);在小鼠,Sema4D主要分布在胚胎的神经系统、肺脏、肾脏及胸腺。在淋巴器官,Sema4D高表达于静止T细胞,静止B细胞、抗原提呈细胞(an⁃ tigen presenting cell,APC,包括树突状细胞)、粒细胞、内皮细胞和上皮细胞表面也存在着Sema4D的表达[3-4]。
Sema4D主要有2个受体,一个是CD72,另一个是plexin-B1,分别在淋巴组织和非淋巴组织优势表达。CD72是Sema4D在淋巴组织的主要功能受体,对B细胞的发育和分化起负性调控作用。Sema4D与CD72结合可以消除CD72的抑制作用,从而使B细胞活化,并提高B细胞的聚集和生存能力及APC的活化和成熟能力[5-6]。plexin-B1是一个钙依赖型膜结合蛋白,在细胞和组织中以单个蛋白形式存在。研究证实,plexin-B1与Sema4D结合可以抑制单核细胞、内皮细胞及树突细胞的迁移。
2 Sema4D在免疫应答中发挥重要作用
Sema4D在免疫系统中发挥重要的生理功能。Sema4D与其受体CD72结合后可抑制CD72胞浆区ITIM的磷酸化及其募集活化SHP-1的作用,阻断CD72介导的抑制作用,对T、B细胞的活化发挥正调节作用。
2.1 Sema4D参与B细胞的活化和存活

图1 Sema4D参与B细胞介导的免疫应答(A、B)及参与T细胞活化与分化过程(C)(根据文献[4]修改)
Sema4D和CD72分别优势表达于T细胞和B细胞,CD40分子强烈刺激诱导生发中心中的B细胞表达Sema4D,而Sema4D通过CD72增强CD40分子依赖的B细胞活化[7-8]。外源性转染Sema4D后能促使人B细胞聚集和存活,而Sema4D缺陷的小鼠存在异常的B细胞及抗体反应。研究发现,年轻的Sema4D缺陷小鼠中CD5+B-1细胞减少而其他亚型的B细胞数量都正常;随着变异小鼠的长大,CD21hiCD23lo边缘区B细胞却逐渐增加。深入研究发现,边缘区B细胞数量的增加常在B细胞受体(BCR)信号缺陷的小鼠中发现,而在缺乏抑制性受体(如CD72)的小鼠中B-1细胞数量增多。因而在Sema4D缺陷的小鼠中较高的BCR信号阈可能促使边缘区B细胞的发展和存活,但阻止了B-1细胞的发展。Sema4D-CD72相互作用参与T细胞在小结间区与B细胞的相互作用,在Se⁃ma4D缺陷的小鼠,用T细胞依赖性抗原免疫后,表现出缺陷的抗体亲和力成熟和抗原特异性生发中心B细胞的弱增殖[9-10]。Sema4D和CD72的相互作用,通过B细胞间的相互作用促进了生发中心的B细胞总数扩增;通过来源于辅助性T细胞的增强的生存信号,协助高亲和力性B细胞进行选择。
2.2 Sema4D在T细胞免疫应答过程中的作用
Sema4D主要由T细胞产生[1],对Sema4D缺陷小鼠的研究表明,Sema4D对T细胞介导的免疫有重要作用。用蛋白抗原对Sema4D缺陷小鼠进行免疫接种后,引起淋巴结CD4+T细胞受抗原再刺激后出现严重受损的增殖反应能力和生成细胞因子的能力。转基因小鼠过表达可溶性Sema4D增强了T细胞应答;然而,Sema4D缺陷的T细胞对CD3单克隆抗体或有丝分裂原如刀豆素的应答正常则表明Sema4D并非通过T细胞自主的方式发挥作用。此外,可溶性重组Sema4D并不影响T细胞激活,研究认为Sema4D是通过提高树突状细胞的活性及提高B细胞的增殖来实现T细胞的分化[11-13]。
2.3 Sema4D对树突状细胞的作用
CD72特异性单克隆抗体能刺激Sema4D产生对树突状细胞几乎所有的效应,表明CD72是树突状细胞中Sema4D的主要受体。重组Sema4D使树突状细胞表面的CD80、CD86和MHCⅡ类分子表达增加,免疫原性增强,可能由CD40分子诱导所引起。由于细胞免疫缺陷,Sema4D缺陷小鼠不会发生由髓磷脂少突胶质细胞糖蛋白肽所诱导的实验性自身免疫性脑脊髓炎,树突状细胞表现出共刺激分子缺陷,IL-12产生障碍。加入外源可溶性Sema4D可使Sema4D缺陷小鼠恢复正常功能,并进一步增强正常树突状细胞的功能[13-14]。因此,Sema4D可以促进树突状细胞的成熟,使抗原特异性T细胞活化并进一步分化。
3 Sema4D在病毒感染性疾病中的作用
在一般急性病毒性感染的患者体内可以发现多特异性的CD4+和CD8+T细胞反应,而在许多慢性病毒感染者体内病毒特异性CD4+和CD8+T细胞存在数量和功能上的障碍,T细胞的应答呈进行性减弱,这种现象将一直伴随着感染的慢性化而恶化,T细胞应答的恶化从早期细胞增殖能力、细胞毒性和产生IL-2能力的下降,逐步到大多数暴露于病毒抗原的T细胞发生功能障碍,随着疾病的进展,甚至连产生IFN-γ的功能也逐步丧失,最终导致T细胞功能衰竭(T cell exhaus⁃tion),无法清除病毒[15-16]。最新研究表明,Sema4D在多种慢性病毒持续感染中发挥重要作用。
3.1 Sema4D在人免疫缺陷病毒慢性持续感染中的作用
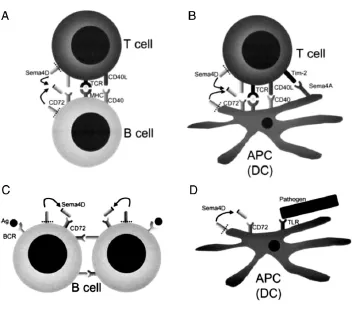
图2 Sema4D在免疫应答不同阶段的作用(根据文献[11]修改)
目前有关Sema4D在病毒感染中的研究主要集中在人免疫缺陷病毒(human immunodeficien⁃cy virus,HIV)感染后的免疫应答。Eriksson等[17]发现,Sema4D对维持CD8+T细胞的功能尤为重要,Sema4D对控制HIV慢性化发挥了重要作用。研究发现:①HIV感染者Sema4D+CD8+T细胞、Se⁃ma4D+CD4+T细胞的比例较健康对照显著降低;②HIV感染早期总CD8+T细胞Sema4D的表达即被破坏,这种状态持续至感染慢性期,而且高效抗逆转录病毒治疗(highly active antiretroviral therapy,HAART)只能部分恢复Sema4D的表达,只有“精英控制者”可以维持正常的Sema4D分泌水平;③HAART能够部分恢复机体初始Sema4D+CD8+T细胞的数量,增加Sema4D+CD8+Tcm细胞的数量;④Sema4D的表达与HIV特异性T细胞应答的幅度相关;⑤Sema4D的丢失与机体免疫活化状态及病毒载量相关;⑥Sema4D表达下调与CD8+T细胞功能低下相关;⑦Sema4D-CD8+T细胞应对抗原刺激的功能不同;⑧病毒复制和促炎性细胞因子不能诱导CD8+T细胞表面Sema4D的丢失。研究表明,Sema4D在HIV感染慢性化中对维持CD8+T细胞正常功能发挥重要作用,Sema4D作为监测T细胞免疫功能的关键标志分子,可能在感染T细胞功能低下方面发挥重要作用[17-18]。
3.2 Sema4D在汉滩病毒感染中的作用
汉坦病毒(Hantavirus)归属布尼亚病毒科,是一种有包膜分节段的负链RNA病毒。汉坦病毒中的汉滩病毒(Hantaan virus,HTNV)可以导致人类致命的肾综合征出血热(hemorrhagic fever with renal syndrome,HFRS)。Liu等[19]研究认为,HFRS感染中可溶性Sema4D水平的增加与疾病的严重程度有关,可溶性Sema4D的表达可能是疾病进展的直接后果。研究发现:①HFRS感染急性期患者血清中的可溶性Sema4D水平较健康者显著增加,而在感染恢复期表达降低;②血小板计数、白细胞计数、血清肌酐水平及血尿素氮水平与可溶性Sema4D的表达紧密相关,是导致其水平升高的独立因素;③相对B细胞和单核细胞,Sema4D在T细胞呈高表达;④HFRS急性期感染时Sema4D+CD4+T细胞,Sema4D+CD8+T细胞,Sema4D+B细胞及单核细胞的表达均有不同程度下降,并且在感染恢复期几乎恢复正常。研究认为,Sema4D在促进B细胞增殖、增加免疫球蛋白产量以及增强T细胞-APC应答中发挥重要作用。3.3 Sema4D在丙型肝炎病毒感染中的作用
丙型肝炎病毒(hepatitis C virus,HCV)为单股正链RNA病毒,属黄病毒科嗜肝病毒属,球形,有包膜。HCV的慢性持续感染增加了终末期肝病发生的风险。HCV的治疗主要依靠IFN-α和利巴韦林,IFN-α除了直接抑制病毒复制,还具有免疫调节功能,但其具体机制尚不清楚。He等[20]研究20例慢性HCV感染患者接受治疗前后B细胞表面Sema4D和CD72的表达,发现HCV感染和IFN-α能够上调Sema4D的表达,而对于发生持续病毒学应答的患者则会降至正常水平。IFN-α诱导的B细胞Sema4D的表达与HCV RNA滴度呈负相关,提示Sema4D的上调可能与机体对HCV感染抑制作用的增强相关。
4 结语
Sema4D是惟一在免疫系统中发现的Sema⁃phorins家族成员,它不但在神经系统中对轴突导向发挥作用,还对T、B细胞的活化及免疫应答起重要的调节作用。最新研究表明,Sema4D在慢性病毒持续感染中发挥重要作用,Sema4D对维持CD8+T细胞的功能尤为重要。Sema4D作为监测T细胞免疫功能的关键标志分子,可能参与慢性病毒感染性疾病T细胞功能低下。进一步探索Se⁃ma4D在慢性病毒持续感染中的作用,将为控制感染慢性化,揭示病毒感染慢性化发病机制、病毒与宿主细胞相互作用机制提供新的线索。
[1]Kikutani H,Kumanogoh A.Semaphorins in interac⁃tions between T cells and antigen-presenting cells[J]. Nat Rev Immunol,2003,3(2):159-167.
[2]Kumanogoh A,Kikutani H.Immune semaphorins:a new area of semaphorin research[J].J Cell Sci,2003, 116(Pt 17):3463-3470.
[3]Nkyimbeng-Takwi E,Chapoval S P.Biology and func⁃tion of neuroimmune semaphorins 4A and 4D[J].Im⁃munol Res,2011,50(1):10-21.
[4]Suzuki K,Kumanogoh A,Kikutani H.Semaphorins and their receptors in immune cell interactions[J].NatImmunol,2008,9(1):17-23.
[5]Wu H J,Bondada S.CD72,a coreceptor with both positive and negative effects on B lymphocyte develop⁃ment and function[J].J Clin Immunol,2009,29(1):12-21.
[6]Kumanogoh A,Kikutani H.The CD100-CD72 interac⁃tion:a novel mechanism of immune regulation[J]. Trends Immunol,2001,22(12):670-676.
[7]Kumanogoh A,Shikina T,Watanabe C,et al.Require⁃ment for CD100-CD72 interactions in fine-tuning of B-cell antigen receptor signaling and homeostatic maintenance of the B-cell compartment[J].Int Immu⁃nol,2005,17(10):1277-1282.
[8]Deaglio S,Vaisitti T,Bergui L,et al.CD38 and CD100 lead a network of surface receptors relaying positive signals for B-CLL growth and survival[J]. Blood,2005,105(8):3042-3050.
[9]Granziero L,Circosta P,Scielzo C,et al.CD100/Plex⁃in-B1 interactions sustain proliferation and survival of normal and leukemic CD5+B lymphocytes[J].Blood, 2003,101(5):1962-1969.
[10]Watanabe C,Kumanogoh A,Shi W,et al.Enhanced immune responses in transgenic mice expressing a truncated form of the lymphocyte semaphorin CD100 [J].J Immunol,2001,167(8):4321-4328.
[11]Kumanogoh A,Kikutani H.Roles of the semaphorin family in immune regulation[J].Adv Immunol,2003,81: 173-198.
[12]Suzuki K,Kumanogoh A,Kikutani H.CD100/Sema4D, a lymphocyte semaphorin involved in the regulation of humoral and cellular immune responses[J].Cytokine Growth Factor Rev,2003,14(1):17-24.
[13]Kumanogoh A,Suzuki K,Ch'ng E,et al.Requirement for the lymphocyte semaphorin,CD100,in the induc⁃tion of antigen-specific T cells and the maturation of dendritic cells[J].J Immunol,2002,169(3):1175-1181.
[14]Li M,O'Sullivan K M,Jones L K,et al.CD100 en⁃hances dendritic cell and CD4+cell activation leading to pathogenetic humoral responses and immune com⁃plex glomerulonephritis[J]. J Immunol, 2006,177(5): 3406-3412.
[15]Shin H,Wherry E J.CD8 T cell dysfunction during chronic viral infection[J].Curr Opin Immunol,2007,19 (4):408-415.
[16]Yi J S,Cox M A,Zajac A J.T-cell exhaustion:char⁃acteristics,causes and conversion[J].Immunology,2010, 129(4):474-481.
[17]Eriksson E M,Milush J M,Ho E L,et al.Expansion of CD8+T cells lacking Sema4D/CD100 during HIV-1 infection identifies a subset of T cells with de⁃creased functional capacity[J].Blood,2012,119(3):745-755.
[18]Mizrahi S,Markel G,Porgador A,et al.CD100 on NK cells enhance IFNgamma secretion and killing of target cells expressing CD72[J].PLoS One,2007,2(9): e818.
[19]Liu B,Ma Y,Yi J,et al.Elevated plasma soluble Se⁃ma4D/CD100 levels are associated with disease severi⁃ty in patients of hemorrhagic fever with renal syn⁃drome[J].PLoS One,2013,8(9):e73958.
[20]He Y,Guo Y,Zhou Y,et al.CD100 up-regulation in⁃duced by interferon-α on B cells is related to hepati⁃tis C virus infection[J].PLoS One,2014,9(12):e113338.
Advances of Sema4D Expression in Chronic Viral Infections
ZHANG Jiu-Cong1*,ZHENG Xiao-Feng2,YANG Yong-Lin1,WANG Yong1
1.Department of Gastroenterology,Lanzhou General Hospital of Lanzhou Military Command,Lanzhou 730050; 2.Department of Gastroenterology,Second Clinical Medical College of Lanzhou University,Lanzhou 730030;China
Sema4D,also known as CD100,was the first discovered immune semaphonrin.Sema4D binding its re⁃ceptor plexin-B1 and CD72 plays a key role in the axon guidance of nervous system,tumor metastasis and activa⁃tion and immune regulation of T,B cells in the immune system through a variety of signal transduction pathways. To date,accumulating evidences have indicated that Sema4D has a pivotal function during chronic viral infection and Sema4D is particularly crucial in the control of chronic infection by maintaining CD8+T cell functions.Se⁃ma4D as a key marker of T cell immune function may involve in the T cell exhaustion in chronic viral infec⁃tions.The following is a brief overview of the recent advance in the role of Sema4D expression in chronic viral in⁃fections.
Sema4D;chronic viral infection;immunomodulation;research progress
R392.1
A
1009-0002(2017)02-0202-05
10.3969/j.issn.1009-0002.2017.02.027
2016-09-18
国家自然科学基金(81500454);兰州军区医药卫生科研项目(CLZ15JA02)
张久聪(1985-),男,医学博士,主治医师
张久聪,(E-mail)zhangjiucong@163.com
*Corresponding author,E-mail:zhangjiucong@163.com
