荔枝海绵(Tethya sp.)形态及愈伤过程的观察研究
2017-04-14夏光远王德祥
夏光远,王德祥*
(1.厦门大学 海洋与地球学院 海洋生物制备技术国家地方联合工程实验室,福建 厦门 361102;2.厦门大学 福建省海陆界面生态环境重点实验室,福建 厦门 361102;3.厦门市海湾生态保护与修复重点实验室,福建 厦门 361102)
荔枝海绵(Tethyasp.)形态及愈伤过程的观察研究
夏光远1,2,3,王德祥1,2,3*
(1.厦门大学 海洋与地球学院 海洋生物制备技术国家地方联合工程实验室,福建 厦门 361102;2.厦门大学 福建省海陆界面生态环境重点实验室,福建 厦门 361102;3.厦门市海湾生态保护与修复重点实验室,福建 厦门 361102)
海绵动物具备很强的愈伤再生能力。荔枝海绵是海绵动物中比较特殊的类群,它有皮质层细胞(cortex)和领细胞层细胞(choanoderm)的初步分化。本文对荔枝海绵的形态结构进行观察,并通过切割的方法,有针对性地移除荔枝海绵的特定结构,观察其愈伤过程,以期了解海绵愈伤过程的主要变化及各部分在愈伤过程中的作用。结果发现,愈伤过程发生的主要变化为未受创部分的细胞向受创部分迁移导致的细胞重新排布;其身体结构中的领细胞层和皮质层对其生存愈伤是必要的,而骨针核心对其愈伤影响较小。
荔枝海绵;形态学;愈伤
1 引言
一般认为,在动物进化中海绵动物很早就分离出来,并进化成区别于其他后生动物的一个侧枝,又称“侧生动物”(Parazoa)[1]。其胚胎发育过程中动物极及植物极细胞的后期分化不同于其他所有后生动物,另外海绵动物体内的领细胞(choanocyte)除了与原生动物的领鞭毛虫类相似之外,在绝大多数其他后生动物中不曾发现。海绵动物具有一些独特的身体结构,如骨架(skeleton)。骨架是由分布在中胶层的钙质或硅质的骨针(spicule)和(或)类蛋白质的海绵质纤维(spongin fiber)或称海绵丝彼此连接而组成,骨针以各种不同的方式排列结合,可以形成不同的复杂结构,以适应不同的环境条件,骨针和骨架的结构也是海绵动物重要的分类依据。骨架支持起了海绵特有的适应固着生活的体内水流通道即水沟系(canal system),后者起到了高等动物相类似的呼吸、循环、消化、排泄、生殖等多个系统的功能。海绵有很强的再生(regeneration)能力,机体受损伤后能恢复其失去的部分,并能诱导体细胞发生胚胎化转变(somatic embryogenesis),机体的所有细胞都能参与结构和机能的重新组织,形成新个体。但对于海绵再生的研究多集中于其存活率以及生物量恢复的时间与速率[2],以及海绵养殖过程中进行移植(explant)等操作所产生的损伤的修复过程[3]。
荔枝海绵属(Tethya)是寻常海绵纲(Demospongiae),荔枝海绵目(Tethyida),荔枝海绵科(Tethyidae)的模式属[4]。它在福建近岸均有分布,但数量较少。其繁殖方式分为有性繁殖和无性繁殖,其中无性繁殖主要是出芽生殖(budding)等方式[5—7]。Nickel等针对它独特的身体结构和生物学特性展开过很多研究。其个体结构由皮质层(cortex)、领细胞层(choanoderm)、骨针(style)组成的核心(center)及贯穿个体的大骨针束(megasclere bundles)组成,细胞分化较其他海绵较为彻底[8—9]。与多数海绵相比较,荔枝海绵具有较为不同的生物学特性。一般海绵的成体多为终身固着,除了开闭出入水孔外几乎完全没有运动和主动形变能力,而荔枝海绵则不然,它能在附着基上进行缓慢的移动[10—11];且整体具有规律性的收缩和舒张的能力,短时间体积变化可达70%,这种规律性舒缩与海绵体内的扁平细胞有关,且会受到神经递质类物质的影响,暗示着这类海绵有着类似原始神经系统的结构[12—14]。
如前所述,荔枝海绵的细胞分化较明显,因此其再生能力预测不像其他海绵那样任意切成小块都能形成独立存活的个体。本研究通过主动地破坏其身体结构,来对这类独特的海绵愈伤再生的能力与过程进行研究。了解这类海绵个体的愈伤能力,将增进我们对海绵这个生物类群的特性和多细胞生物身体结构形成与维持的认知。
2 材料与方法
2.1 实验材料
荔枝海绵实验样品25个,采集于福建省漳州市东山湾(23.801 5°N,117.592 8°E),直径在14~31 mm之间不等。采样时小心地将荔枝海绵从附着基上剥离,避免离水,采用PE塑料袋密封并充入医用纯氧的方式带回实验室。将样品放入实验室1 200 L的循环恒温养殖系统暂养,保持海水温度(24±1)℃、盐度(32±1),不间断流水,自然光照,不额外投喂饵料。
2.2 实验方法
2.2.1 海绵骨针标本的制作
骨针标本的制作方法为:取约20 mg海绵样品于1.5 mL离心管中,加5%次氯酸钠溶液,55℃水浴至样品完全消化。离心(500 r/min,2 min),弃去上层清液,用蒸馏水冲洗骨针样品,重复离心清洗3~4次,将骨针样品滴至载玻片上晾干,滴加中性树脂,盖上盖玻片,待中性树脂凝固即完成玻片制作。
2.2.2 荔枝海绵骨架冷冻切片的制作
骨架切片制备在冷冻切片机(莱卡CM1900型)上进行。取待观察的海绵样品置于载玻片上,用包埋剂包埋,放在样品头上充分冷却后徒手切片,样品片厚度约1 mm。切面要通过样品的核心且与创口面垂直,以最大程度体现愈伤过程发生的变化。将切下的样品片置于另一片载玻片上,用纯水冲去包埋剂。吸去多余的水分,滴加二甲苯浸没样品片使样品脱水透明。最后滴加中性树脂,盖上盖玻片,完成制作。
2.2.3 对荔枝海绵皮质层和领细胞层的切割方式
为研究荔枝海绵皮质层、领细胞层及骨针核心在荔枝海绵创伤愈合过程中的作用,设计了两类主要的切割方式。一是仅损伤皮质层,切除部分皮质层(P)时尽量避免对领细胞层(L)的损伤,创伤面在皮质层和领细胞层之间,如图1a~d所示。有7个样本实施图1a切割方式,其中2个样本用于愈伤过程的持续观察,另5个用于骨架切片的连续取样。图1b~d切割方式则各有2个平行样。二是将皮质层、领细胞层和骨针核心同时损伤的切割方式,切割方向平行于大骨针束,避免对大骨针束的损伤。如图1e、1f所示,其中实验e为将荔枝海绵个体等分为两部分,有5个平行样本;实验f将荔枝海绵个体上的一小部分X切下,余下部分为Y,有7个平行样本。
2.2.4 愈伤过程的观察与记录
在实验过程中,每天定时进行观察、拍照,记录实验材料的形态变化。当观察的海绵死亡或愈伤达到领细胞层愈合,皮层愈合等状态时,将实验材料从养殖水体中取出直接放入-22℃冰箱冻存,制备骨针及骨架切片观察。
3 实验结果
3.1 荔枝海绵的形态
与多数海绵没有固定的形态不同,荔枝海绵的形态结构较为固定,主要特征为(图2):整体为球形,最外层是坚韧而富弹性的皮质层,包裹着疏松的领细胞层,个体中心有一个由针状骨针组成的核心,由这个核心辐射出许多同样由针状骨针平行排列组成的大骨针束(megasclere bundles),大骨针束穿透领细胞层到达皮质层,原来基本相互平行的针状骨针或放散成花束状结构(bouquet),在皮质层表面形成一个个疣状突起(verrucose prominence);或者保持互相平行而穿出海绵表面,形成刺状突起(spinous prominence),刺状突起的末端可能有芽体(bud)。皮质层表面的突起数量和大骨针束的数量是紧密相关的,海绵表面的每个疣状或刺状突起都对应着内部的一个大骨针束,二者在空间上的位置相对固定。
荔枝海绵的这种大骨针束贯穿皮质层-领细胞层-核心的结构模式,其细胞的分化程度较高。每个个体包含的4种基本结构,每一种都由不同种类的骨针(图3)、不同的组合方式形成其独特的骨架结构。骨针核心和大骨针束几乎完全由棒针状骨针(style)组成,但是排列方式不同;尺寸小得多的头星状骨针(tylaster)仅分布在皮质层和外层领细胞层之中,至于大小介于前二者之间的球星形骨针(spheraster)或球真星骨针(spheroxyaster),则完全分布于皮质层之中。

图1 荔枝海绵的切割方式Fig.1 The cutting patterns of Tethya sp.a~d中灰色部分为P,其余部分为L;e中切割形成的两部分没有区别;f中灰色部分为X,其余部分为YIn a~d,P represent the gray part,and L represent the rest part;in e,there is no difference between the two parts;in f, X represent the gray part,and Y represent the rest part

图2 荔枝海绵(Tethya sp.)的结构Fig.2 The structure of Tethya sp.

图3 荔枝海绵(Tethya sp.)的骨针Fig.3 The spicule of Tethya sp.

图4 离体的海绵皮质Fig.4 The cortex cut off from the sponge sample

图5 离体皮质的骨架结构变化Fig.5 The change of skeleton structure of cortex cut off from the sponge body

图6 损失小半皮质的荔枝海绵个体愈伤过程Fig.6 The wound healing process of an individual with less than half cortex lost
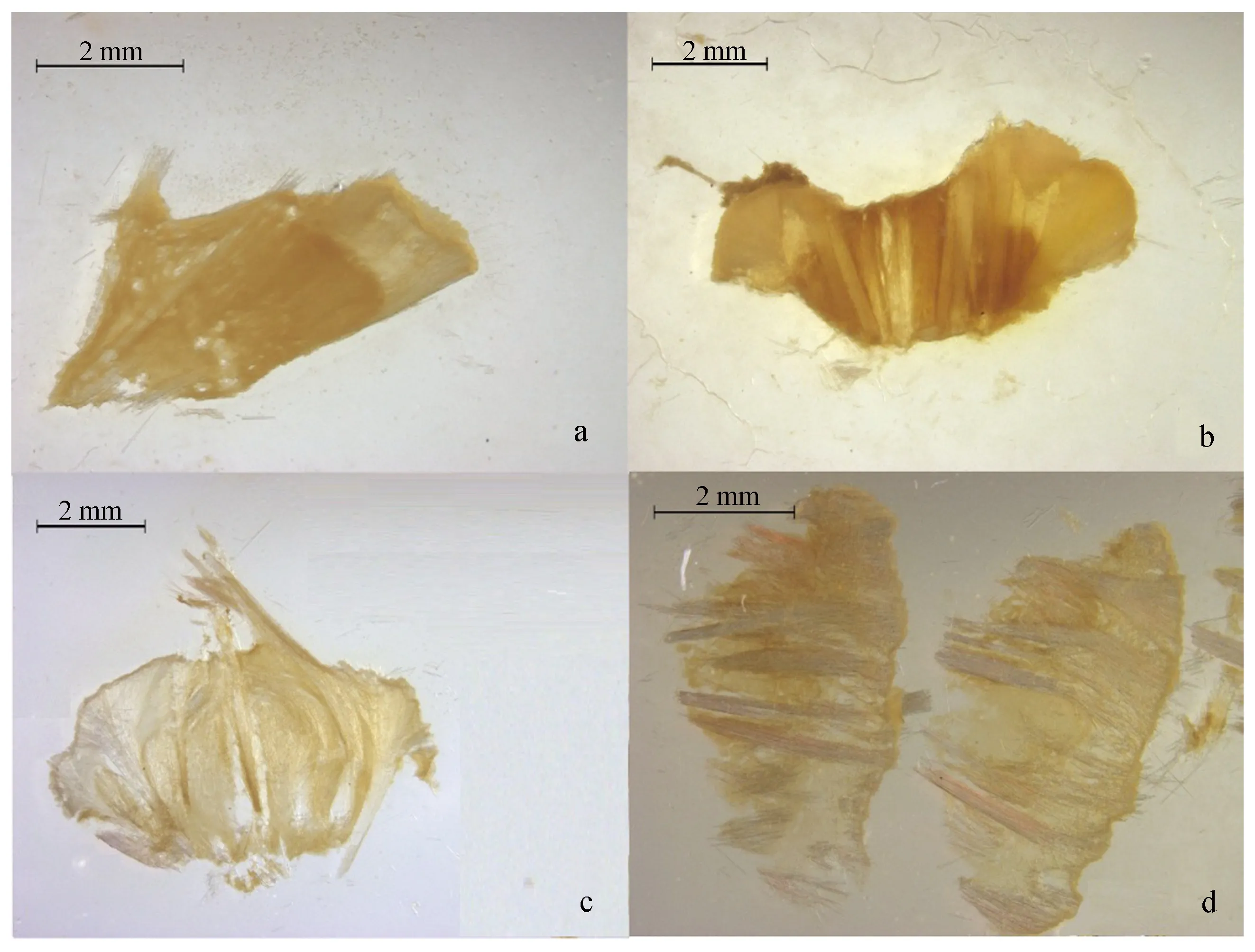
图7 损失小半皮质的个体骨架修复过程Fig.7 The skeleton healing process of the individuals with less than half cortex lost
3.2 皮质层和领细胞层在愈伤过程中的作用
3.2.1 单纯皮质层在切割后发生的变化
a~d切割方式(图1)获得的P部分均为不含领细胞的离体皮质层,共有13个样本,全部在切割48 h之内死亡,表现为离体皮质层进行剧烈的形态变化(图4),表面出现白色霉变,之后腐烂。形态变化大体规律为海绵结构沿大骨针束的方向极度伸长,在原来的表面上形成许多突起,其形状介于正常状态下的刺状突起和疣状突起之间。骨架切片显示(图5),皮质层中的针状骨针沿大骨针束向外移动显得没有规律,排列杂乱,不像形成刺突的骨针那样排列有序。
3.2.2 失去不同量皮质层的个体受创后的愈伤现象
a~d切割方式分离的L部分为失去不同量皮质层的个体。其中,通过a切割方式分离的损失小半(约全部的1/8)皮质层的样本共7个,全部愈伤完成或有愈伤完成的趋势(有几个个体未愈伤完成即用于切片观察);通过b切割方式分离的L部分(损失一半皮质层)共2个,一个愈伤完成,另一个在实验持续的14 d内一直处于愈伤的过程中,但截至实验结束时尚未完成愈伤;通过c切割方式分离的L部分(损失大半皮质层)共2个,在切割后3~5 d都死亡,表现为形态剧烈变化得很不规则,表面覆盖白色霉变,有腐臭气味。通过d切割方式形成的L部分(失去全部皮质层)共2个,一个切割后2 d即死亡,状态与c方案切割的L部分相似。另一个截至实验结束取样分析的第14 d为止也未完成愈伤。以损失小半皮质层的样本为例,生物的个体愈伤情况和骨架修复过程如图6和图7所示,骨针核心受损的个体愈伤过程如图8所示。由于图7中所用切片来自不同个体,各阶段愈伤的时间存在个体差异,故各愈伤阶段不标示切割后时间,下文图9和图11情况相同。
综合生活状态观察以及骨架切片观察的结果,损失少量皮质层的荔枝海绵愈伤过程具体如下:
(1)创伤初期,内部的领细胞层暴露在外,创口显得十分粗糙;而皮质层具有一定收缩能力,在受创时收缩,甚至可能拉扯创口使之扩大;
(2)切割之后1 d内,暴露的领细胞层会收缩,表面变得平滑,此时创口被领细胞层所围绕的大骨针束会突出于领细胞层之外;
(3)此后(切割后约2~3 d内),创口部分突出在外的大骨针束也慢慢收缩,创口整体变得平整。与创口变平整的同时或稍晚些,创口边缘的皮质层开始逐渐向创口中心迁移,创口周围未受损的大骨针束由原来的笔直状态向创口中心弯折,二者同步表现出由四周向中心汇聚的趋势;
(4)切割后3 d至1周间,创口周围迁移来的皮质逐渐完全覆盖暴露的领细胞层,使创口愈合,但总体形状尚未恢复球形;
(5)切割约1周后,个体形态基本恢复。
当原有的皮质层大量或完全损失时,生物难以通过原有皮质的迁移来缩小创口,因而很容易死亡,死亡率随着皮质层损失的程度增大而上升。
3.3 骨针核心在愈伤过程中的作用
综合e、f两种切割方式所形成的3种受损个体愈伤结果,在完成愈伤的个体中,既有骨针核心保持完整的,也包括骨针核心受损的乃至彻底失去骨针核心的。各种受损个体在切割后均有一定概率愈伤形成完整的个体。骨针核心受损的个体,在其外部形态完全恢复正常时,骨针核心仍表现出受损的状态;而失去骨针核心的个体,在愈伤过程中通过大骨针束聚集而再次形成了新的骨针核心。
3.3.1 骨针核心受损的个体愈伤过程
e切割方式使得个体包括骨针核心被平均分成了两部分。5个荔枝海绵整体切割形成10个实验材料,其中来自2个个体的4个实验材料死亡,而6个成功愈合,存活率较高。其愈伤过程为:
(1)因为荔枝海绵皮质层的收缩性导致受创个体的形变,创口会向外凸起,使原呈半球形的海绵略呈锥形,受创的骨针核心位于锥形的尖端。因为切割方向基本没有切断任何大骨针束,所以大骨针束很少暴露,创口十分平滑;
(2)切割后1~5 d之间,观察到创口周围的皮质细胞向创口转移,创口逐渐缩小,而个体内部的大骨针束也表现出了相应的弯折变化,但是海绵块的形状在愈伤过程中保持锥形。
(3)直到皮质层结构完全覆盖创口后(切割5 d以后),骨针核心才慢慢向个体中心转移,个体逐渐恢复球形;
(4)直至切割后15 d实验结束,个体形状基本恢复球形,切片后发现骨针核心的形状和位置都没能恢复,一直保持着受损的半球形,稍偏向创口方向。
个体愈伤情况和骨架修复过程见图8和图9。

图8 骨针核心受损的个体愈伤过程Fig.8 The wound healing process of an individual with the sponge center wounded

图9 骨针核心受损的骨架修复过程Fig.9 The skeleton healing process of an individual with the sponge center wounded
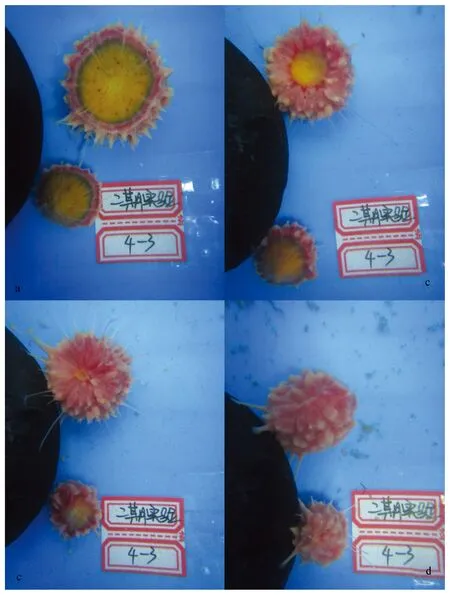
图10 骨针核心未受损的个体和无骨针核心的损伤部分愈伤过程Fig.10 The wound healing process of an individual with the sponge center unwound-ed/lost

图11 骨针核心完全损失的骨架修复过程(创口均在视野上方)Fig.11 The skeleton healing process of an individual with the sponge center lost

图12 可能的受损成分排出体外的过程与被排出部分的切片Fig.12 The process of the possibly damaged part be discharged from main body, and the skeleton section of the discharged part
3.3.2 骨针核心未受损的个体愈伤过程
f切割方式形成的Y部分共7个样本,2个未能愈合而死亡,另5个愈伤完成或有趋势愈伤为新的荔枝海绵个体的趋势(中途被取样用于切片观察),其存活率较高。因为f切割方式的切割方向基本是沿着大骨针束方向,所以创口比较平滑,大骨针束很少暴露,无需经历创口平滑阶段。对于这些样本,失去的部位只占整体的一小部分,对整体形状的影响不很大,其个体愈伤过程(图10较大的样本)及骨架修复过程与损失少量皮质层的个体(如a的切割方式)愈伤过程较为接近,不再重复描述。
3.3.3 无骨针核心的损伤部分愈伤过程
f切割方式形成的X部分共7个样本,2个未能愈伤而死亡(与3.3.2中Y部分死亡的2个样本分别来自同一整体),另5个愈伤完成或有愈伤为新的荔枝海绵个体的趋势,其存活率较高。这一组样本切割后失去了大部分的生物量,其形状呈锥形,且锥形的尖端没有骨针核心。愈伤的过程描述如下:切割后,创口周围的皮质层即开始向创口转移,使得创口缩小直至消失;而锥形的海绵块尖端分散的大骨针束随着创口缩小,逐渐汇聚成一个大的平行的骨针簇。创口消失的同时,骨针簇汇聚的地方即新个体的边缘形成了新的骨针核心。在创口消失之后,新的骨针核心的位置由新个体的表面逐渐移动至中心。其个体愈伤情况和骨架修复过程见图10(较小的样本)和图11。
3.4 在切割中受损的成分的可能去向
切割不一定会造成整个生物个体的死亡,但一定会造成创口附近一些结构成分的损伤。这些受创的成分,如折断的骨针及大骨针束,受损的细胞在愈伤过程中会逐渐形成一些不规则的突起,并最终被排出。这些被排出的部分结构复杂,骨针种类混杂、排列凌乱,同时含有大量不完整的骨针如图12所示。
4 结论与讨论
总结实验结果,荔枝海绵身体各部结构在愈伤中起到的作用如下:(1)由离体的独立皮质层无法存活的现象可知,领细胞层的存在对于个体的存活是必要的;(2)皮质层在愈伤过程中起到迅速缩小伤口、保护内部的作用,其存在对于生存和愈伤也是十分重要的。损失少量皮质层的个体能够愈伤而恢复正常,但其成活率随着受创程度的增大而下降;(3)骨针核心的主要成分是骨针而不是细胞,因此对生物的生存影响较小,其存在、受创与否对荔枝海绵能否愈伤再生无明显影响,它仅仅是生物形态上的中心,而非功能上的中心。
原细胞是海绵体中必不可少的一种细胞,通常认为它能够多向分化,从而转化为多种其他细胞类型,在海绵细胞中扮演类似干细胞的角色,可能是愈伤过程的主要参与者。本实验并未从细胞分化层面对荔枝海绵的愈伤过程进行观察研究,因此不能确定原细胞分化形成新的特化细胞过程在愈伤过程中具体的方式和作用。但在本实验愈伤成功的个体中,愈伤过程中创口周围的皮质层和大骨针束表现出了同步的向创口中心汇聚的趋势。由于皮质层与大骨针束的相对位置是被花束状结构或刺状突起结构所固定的,所以通过大骨针束的弯曲现象可以判定荔枝海绵愈伤过程中覆盖创口的皮质层结构大部分来源于原有的皮质层迁移。保持了接近原生物的各种细胞比例的残体如失去骨针核心的锥形结构,可以愈伤恢复;如果大比例损失某一种细胞,使得生物体各种细胞的相对比例失衡,则会最终导致生物的死亡,如各种离体的单纯皮质层,或损失大量皮质层的领细胞层,即使保留的总生物量较大,最终也都会死亡。如此的实验结果表明,荔枝海绵愈伤过程的早期,较少发生细胞种类的转变,而主要是通过对已有细胞迁移重新排布来完成整体结构的复原。可以推测,在荔枝海绵的愈伤过程中细胞的迁移和分化是同时存在的,在荔枝海绵受损的早期主要是通过各部分细胞的迁移进行伤口的愈合;在愈伤的后期再通过细胞的分化及增殖完成结构的修复。前者在愈伤方面的效果要远比后者明显,因而掩盖了后者的作用效果,即直接在创口区域形成愈伤结构。
综上,我们可以如此概括荔枝海绵的愈伤现象,荔枝海绵愈伤的策略主要是对残存已分化细胞的重新排布,先通过皮质层细胞的迁移和大骨针束的弯折尽快使皮质层结构覆盖创口,减少受创领细胞层的暴露,而后再调整整体的形态恢复正常。切割中受损的成分将在此愈伤过程中被逐渐排出体外。
将荔枝海绵的愈伤与其他多细胞生物进行对比,可发现这种以细胞重新排布为主的愈伤方式是十分独特的,但也是十分有效的。例如,扁形动物也具有极强的细胞再组织以及再生能力,被切割后仍有能力再生为完整的个体或特殊的异形个体,这种能力主要来自于其体内的未分化细胞。切割后,扁形动物通过肌肉的收缩来缩小创口,在创口处形成含大量成体未分化细胞的胚芽组织,再由这些胚芽组织分化形成失去的身体结构,过程伴以大量细胞的有丝分裂与分化[15]。在荔枝海绵的愈伤过程中,皮质层细胞的迁移起到了与肌肉收缩减小创口相似的作用,然而与未分化细胞的分化作用类似的过程却没有在愈伤过程中起到主要作用,而是直接由各类细胞迁移重新排布完成愈伤。从物质和能量消耗的角度来看,直接利用现存细胞重排愈伤比起未分化细胞分化形成新的身体结构要节约得多。前者仅仅是细胞位置的移动,而后者涉及大量的有丝分裂以及蛋白质的合成。在本实验没有投放饵料的营养条件下,采取将现存细胞重新排布的愈伤方法无疑是经济而快速的愈伤方法。
荔枝海绵通过细胞迁移完成愈伤的主要内因可能是海绵体内变形细胞的移动[16],这种动态组织的存在表明了海绵动物在进化过程中介于原生动物细胞群与其他组织细胞更加专门化的后生动物类群之间的地位。而不完整的个体能通过细胞的动态排布完成愈伤,说明它们有一种宏观调控的机制,能感受到自身的受创,进而调控已分化的细胞迁移到合适的位置,来恢复正常的完整身体结构。这种整体调控的分子以及基因机制尚不明了,需要进一步的研究来揭示,以增进我们对多细胞生物身体结构形成与维持的认知。
[1] 刘凌云, 郑光美. 普通动物学[M]. 北京: 高等教育出版社, 2009.
Liu Lingyun, Zheng Guangmei. General Zoology[M]. Beijing: Higher Education Press, 2009.
[2] Duckworth A R. Effect of wound size on the growth and regeneration of two temperate subtidal sponges[J]. Journal of Experimental Marine Biology and Ecology, 2003, 287(2): 139-153.
[3] Duckworth A. Farming sponges to supply bioactive metabolites and bath sponges: a review[J]. Marine Biotechnology, 2009, 11(6): 669-679.
[5] Gaino E, Liaci L S, Sciscioli M, et al. Investigation of the budding process inTethyacitrinaandTethyaaurantium(Porifera, Demospongiae)[J]. Zoomorphology, 2006, 125(2): 87-97.
[6] Cardone F, Gaino E, Corriero G. The budding process inTethyacitrinaSar& Melone (Porifera, Demospongiae) and the incidence of post-buds in sponge population maintenance[J]. Journal of Experimental Marine Biology and Ecology, 2010, 389(1/2): 93-100.
[7] Corriero G, SarM, Vaccaro P. Sexual and asexual reproduction in two species ofTethya(Porifera: Demospongiae) from a Mediterranean coastal lagoon[J]. Marine Biology, 1996, 126(2): 175-181.
[8] Nickel M, Donath T, Schweikert M, et al. Functional morphology ofTethyaspecies (Porifera): 1. Quantitative 3D-analysis ofTethyawilhelmaby synchrotron radiation based X-ray microtomography[J]. Zoomorphology, 2006, 125(4): 209-223.
[9] Nickel M, Bullinger E, Beckmann F. Functional morphology ofTethyaspecies (Porifera): 2. Three-dimensional morphometrics on spicules and skeleton superstructures ofT.minuta[J]. Zoomorphology, 2006, 125(4): 225-239.
[10] Nickel M. Like a 'rolling stone': quantitative analysis of the body movement and skeletal dynamics of the spongeTethyawilhelma[J]. Journal of Experimental Biology, 2006, 209(15): 2839-2846.
[11] Fishelson L. Observations on the moving colonies of the genusTethya(Demospongia, Porifera)[J]. Zoomorphology, 1981, 98(1): 89-99.
[12] Nickel M. Kinetics and rhythm of body contractions in the spongeTethyawilhelma(Porifera: Demospongiae)[J]. Journal of Experimental Biology, 2004, 207(26): 4515-4524.
[13] Ellwanger K, Eich A, Nickel M. GABA and glutamate specifically induce contractions in the spongeTethyawilhelma[J]. Journal of Comparative Physiology A, 2007, 193(1): 1-11.
[14] Ellwanger K, Nickel M. Neuroactive substances specifically modulate rhythmic body contractions in the nerveless metazoonTethyawilhelma(Demospongiae, Porifera)[J]. Frontiers in Zoology, 2006, 3: 7.
[15] Nimeth K T, Egger B, Rieger R, et al. Regeneration inMacrostomumlignano(Platyhelminthes): cellular dynamics in the neoblast stem cell system[J]. Cell and Tissue Research, 2007, 327(3): 637-646.
[16] Bond C. Continuous cell movements rearrange anatomical structures in intact sponges[J]. Journal of Experimental Zoology, 1992, 263(3): 284-302.
Observation on the morphology and wound-healing process ofTethyasp.
Xia Guangyuan1,2,3,Wang Dexiang1,2,3
(1.State-ProvinceJointEngineeringLaboratoryofMarineBioproductsandTechnology,CollegeofOceanandEarthSciences,XiamenUniversity,Xiamen361102,China;2.FujianProvincialKeyLaboratoryforCoastalEcologyandEnvironmentalStudies,XiamenUniversity,Xiamen361102,China;3.XiamenCityKeyLaboratoryofUrbanSeaEcologicalConservationandRestoration(USEeR),Xiamen361102,China)
Sponges of Porifera have powerful regeneration capacity. Among them, the genusTethyais a special group, which has differentiation in cortex and choanoderm. In this paper, the morphology ofTethyasp., and its wound-healing process were studied by removing specific part of some tissues, to understand the transformation ofTethyasp. and the role of different tissues in the healing process. The results showed that the main transformation in the wound-healing process is the cell rearrangement resulted by the cell migration from the unwounded to the wounded part, the cortex and choanoderm are vital to the survival ofTethya; while the spicule center is less importance in the wound-healing process.
Tethyasp.;morphology;wound-healing
10.3969/j.issn.0253-4193.2017.04.009
2016-09-09;
2016-12-15。
国家自然科学基金海洋创新科研基金(J1210050);厦门南方海洋研究中心项目(13GYY002NF07)。
夏光远(1994—),男,辽宁省锦州市人,主要开展海绵动物的生态学研究。E-mail:22320151152142@stu.xmu.edu.cn
*通信作者:王德祥,男,福建省漳浦县人,副教授,主要从事海绵动物分类学及生态学研究。E-mail:dxwang@xmu.edu.cn
S968.9
A
0253-4193(2017)04-0089-12
夏光远,王德祥. 荔枝海绵(Tethyasp.)形态及愈伤过程的观察研究[J].海洋学报,2017,39(4):89—100,
Xia Guangyuan,Wang Dexiang. Observation on the morphology and wound-healing process ofTethyasp. [J]. Haiyang Xuebao,2017,39(4):89—100, doi:10.3969/j.issn.0253-4193.2017.04.009
