水稻根系形态与氮素吸收累积的相关性分析
2017-04-14陈晨龚海青张敬智郜红建
陈晨,龚海青,张敬智,郜红建
(安徽农业大学资源与环境学院,安徽合肥 230036)
水稻根系形态与氮素吸收累积的相关性分析
陈晨,龚海青,张敬智,郜红建*
(安徽农业大学资源与环境学院,安徽合肥 230036)
【目的】氮肥过量施用,不仅造成氮肥大量流失,还增加了农业生产成本,对生态环境带来了巨大的威胁。水稻根系形态作为影响养分吸收和利用的主要因素之一,明确其与氮素吸收累积的相关性是提高氮素利用效率、降低环境污染的有效途径。【方法】利用营养液培养方法,研究了 55 个水稻品种在 NH4+-N 和 NO3--N 供应条件下苗期植株生物量、氮含量和氮素累积量及其与根系形态指标的相关性。【结果】在 NH4+-N 培养下,水稻营养指 标 与 根系形态指标的相关性高于其在 NO3--N 培养下的相 关 性 。在相同供氮水平下,供应 NH4+-N的水稻苗期平均生物量为 55.77 mg/plant,比供应 NO3--N 的量高 4.94 mg/plant;水稻苗期平均氮含量为 4.22%,比供应 NO3--N 的高 0.72%;水稻苗期平均氮累积量为 1.91 mg/plant,比供应 NO3--N 的苗期平均氮累积量高 0.67 mg/plant。在 NH4+-N 和 NO3--N 两种氮素形态培养条件下,水稻根系形态指标品种间根尖数变异系数最大,平均根系直径变异系数最小。总根体积、总根面积、总根长、分枝数四个形态指标与植株生物量、植株氮含量、植株氮累积量相关性最为显著,且相关系数 (r) 呈总根体积 > 总根面积 > 总根长 > 分枝数的规律。在 NH4+-N 培养下的水稻营养指标与根系形态指标的相关性要高于其在 NO3--N 培养下的相关性。【结论】水稻苗期总根体积、总根面积、总根长、分枝数可作为水稻氮高效评价的重要指标。
水稻;根系形态;生物量;氮累积量;相关性分析
根系在植物养分吸收和生长发育过程中起着极为重要的作用。水稻根系既是吸收养分和水分的重要器官,也是很多物质同化、转化或合成的场所,还是与地上部进行物质交流的代谢器官,其生长情况会直接影响整个水稻的生长发育、营养水平和产量水平[1]。根系发达、根生物量高、根系体积大、根系吸收表面积大是氮素高效利用型水稻品种的根系特征[2]。水稻利用氮的形态主要有 NH4+-N 和 NO3--N,氮素形态会在很大程度上影响水稻的生长和对氮的吸收,水稻苗期根内缺少硝酸还原酶,生育初期吸收硝态氮的速率较低;而根部细胞色素氧化酶占优势,吸收的铵态氮较多,所以水稻苗期吸收铵态氮的效率高于硝态氮。随着水稻生长发育,体内黄酶逐渐占优势,硝酸还原酶活性的提高,吸收硝态氮的 速 率 逐 渐 加 快[3]。 我 国 水 稻 平 均 氮 肥 使 用 量 为190~200 kg/hm2,氮肥施用过量,既造成肥料资源浪 费 , 又 造 成 环 境 污 染[4]。 因 此 , 提 高 氮 素 利 用 效率,对于减少氮肥施用量和保护环境具有极其重要的意义。有关提高水稻氮素利用效率的途径和方法等方面已有一些研究。朱兆良认为,对化学肥料本身进行改性,开发更适合作物生长的新型肥料可以有效地提高作物的氮素利用效率[5]。改进施肥和管理措施是提高肥料利用效率,减少氮素损失的有效途径。筛选氮高效品种,即通过水稻营养遗传改良以培育出能高效吸收利用氮素的高效品种,也是行之有效的提高氮效率的手段[6]。
根系形态学特征与植物养分效率有着密切的关系[7]。戢林等[8]探讨水稻苗期至抽穗期不定根、粗分枝根和细分枝根的长度、表面积和体积等形态指标与氮吸收的关系,指出水稻产量和氮利用效率呈现极显著的基因型差异,低氮水平下,氮高、低利用效率基因型间的根系形态指标差异显著。樊剑波[9]研究表明,根干重、总根长、根系表面积、根冠比等指标与氮素积累量呈很好的相关关系,在水稻生产中根系形态参数是决定其高效吸收和利用氮素营养的决定性因素。已有的研究多关注对根生长分布特征的定性描述,对根系表面积、根系直径、根尖数等形态参数缺少有效的分析手段,有关水稻根系形态与养分吸收利用之间的关系尚不清楚。研究水稻根系形态与氮素吸收利用的关系,揭示水稻高效吸收利用氮素的根系形态学特征,对于筛选氮高效水稻品种,提高水稻氮素利用效率具有重要意义。
本研究采用营养液培养方法,利用根系专用扫描仪 (Epson perfection V700 photo,Japan) 和根系形态专用分析软件 (Win RHIZO_Pro V2007d, Regent Instrument Inc. Canada) 研究了 55 个水稻品种在相同供氮水平,不同供氮形态 (NH4+-N 和 NO3
--N) 条件下苗期植株生物量、氮含量和氮素累积量及其与总根长、总根面积、总根体积、平均根系直径、根尖数、分枝数、交叉数等根系形态指标的相关性。通过对根系形态指标进行变异特征分析,拟合植株生物量、氮含量、氮累积量与根系形态指标之间的最优回归模型,为氮高效种质的筛选提供理论依据。
1 材料与方法
1.1 试验材料与设计
长江中下游地区主推的 55 个水稻品种/系 (编号1~55) 分别由湖南省水稻研究所、安徽农业大学、安徽省农业科学院和南京农业大学提供 (表 1)。
选择饱满一致的水稻种子,用 10% 的双氧水消毒 30 min 后,自来水冲洗 2~3 遍,再用蒸馏水清洗干净,放入底部垫有浸润滤纸的发芽盒。水稻种子置于 28℃ 恒温培养箱中避光催芽,待出现胚芽鞘后,移至安科发芽纸上培养。将发芽种子沿宽边均匀放置在发芽纸 (规格 38#,10 英寸 × 15 英寸)上,卷成圆筒状后用亚克力支撑架固定,垂直放入蒸馏水中,光照培养 2 天后,依次在国际水稻所 1/4和 1/2 倍标准浓度的水稻专用营养液中分别培养 4天,1 倍标准浓度的水稻专用营养液中培养 7 天,用蒸馏水冲洗水稻根系后,移入相同氮浓度的 NH4+-N和 NO3--N 营养液中培养 10 天。国际水稻所水稻专用营养液的组成 (mg/L):NH4NO3114.3、NaH2PO4· 2H2O 46.8、K2SO4174、MgSO4·7H2O 393.6、CaCl2111、MnCl2·4H2O 1.98、 (NH4)6Mo7O24·4H2O 0.124、H3BO30.62、ZnSO4·7H2O 0.0574、CuSO4·5H2O 0.1、FeSO4·7H2O 19.5、柠檬酸 30。含NH4+-N 和 NO3

表1 供试水稻品种及编号Table 1 The selected rice cultivars and numbering
--N的营养液分别用 (NH4)2SO4和 KNO3替换 NH4NO3,含 NH4
+-N 营 养 液 中 加 入 5.89 mg/L 的 双 氰 胺(C2H4N4) 抑制可能发生的硝化作用,同时设置只加入5.89 mg/L 的双氰胺 (C2H4N4) 不含 NH4+-N 的营养液为对照。培养液每 2 天更换一次,pH 用 1 mol/L HCl或 1 mol/L NaOH 调节至 5.5 ± 0.1。每个水稻品种 3次重复,每次试验重复培养 6 株水稻幼苗。
1.2 测定项目及方法
水稻培养至六叶期时,分别收获植株的茎叶和根部样品。茎叶和根部样品于 105℃ 下杀青 30 min后,置 75℃ 下烘干至恒重,称量茎叶干重与根部干重,计算整株生物量、氮含量、氮累积量。水稻茎叶和根部样品经 H2SO4-H2O2消煮,凯氏定氮法测定植株氮含量。茎叶氮累积量、根部氮累积量和整株氮累积量分别为茎叶、根部、整株干重与其氮含量之积。
收获时,每个水稻品种从每次重复试验中选择 1株水稻用于根系形态分析试验。新鲜水稻根系先用蒸馏水冲洗干净表面附着物,用根系形态专用扫描仪 (Epson perfection V700 photo,Japan) 数字化扫描,然后用根系形态分析专用软件 (WinRHIZO_Pro V2007d,Regent Instrument Inc. Canada) 分析总根长、总根面积、总根体积、平均根系直径、根尖数、分枝数、交叉数等根系形态参数,根系形态参数为 3 次重复试验的平均值。
1.3 数据分析方法
数据采用 Excel 2010、Origin 9 和 SPSS 19.0 软件进行统计分析,采用 F 检验分析不同参数之间的显著性。根系形态指标采用根系分析系统软件WinRHIZO_Pro V2007d (Regent Instrument Inc., Canada) 进行定量分析。采用逐步回归分析建立NH4+-N 和 NO3--N 培养条件下水稻干重、氮含量、氮累积量与根系形态参数之间的最优回归方程。
2 结果与分析
2.1 不同形态氮素培养下水稻苗期各性状分析
水稻在 NH4+-N 和 NO3--N 培养条件下的苗期植株生物量差异显著。在 NH4+-N 培养下,水稻生物量在11.31~150.77 mg/plant之间,平均为 55.77 mg/plant,变异系数为 0.73,其中以两优 766 的生物量最高,为 150.77 mg/plant,以丰两优品种的生物量最低,为11.31 mg/plant。在 NO3--N 培养下,水稻生物量在11.73~141.27 mg/plant之间,均值为 50.83 mg/plant,变异系数为 0.73,其中以甬优 9 号的生物量最高为141.27 mg/plant,以隆香优 130 品种的生物量最低,为 11.73 mg/plant。在相同供氮水平下,不同水稻品种在 NH4
+-N 培 养条件下的平均生 物 量 高于其在NO3
--N 培养条件下平均生物量 (图 1 A)。水 稻 植 株 在 NH4+-N 培 养 条 件 下 的 氮 含 量 在1.96%~9.46% 之间,均值为 4.22%,变异系数为 0.44,其中以湘早籼 24 号的含氮量最高,为 9.46%,以镇稻 14 号品种的含氮量最低,为 1.96%。水稻植株在NO3
--N 培养条件下的氮含量在 1.49%~8.13% 之间,均值为 3.50%,变异系数为 0.61,其中以丰源优 299的含氮量最高,为 8.13%,以镇 9424 品种的含氮量最低,为 1.49%。在相同供氮水平下,不同水稻品种在 NH4+-N 培养条件下的平均含氮量比其在 NO3--N培养条件下平均含氮量高 0.72% (图 1 B)。
水 稻 植 株 在 NH4+-N 培 养 下 的 氮 累 积 量 在0.36~4.78 mg/plant之间,均值为 1.91 mg/plant,变异系数为 0.61。其中以甬优 9 号的氮累积量最高,为 4.78 mg/plant,以镇稻 14 号品种的氮累积量最低,为 0.36 mg/plant。在 NO3--N 培养下,水稻氮累积量在 0.47~3.13 mg/plant之间,均值为 1.24 mg/plant,变异系数为 0.47,其中以广两优 3905 的氮累积量最高为 3.13 mg/plant,以镇稻 14 号品种的氮累积量最低,为 0.47 mg/plant。在相同供氮水平下,不同水稻品种在 NH4+-N 培养条件下的平均氮累积量高于其在NO3
--N 培养条件下平均氮累积量 (图 1 C)。
2.2 不同形态氮素培养条件下水稻苗期根系形态变异特征
根系形态是品种间差异的重要特征,变异系数越大,水稻品种间的差异性越大。NH4+-N 和 NO3
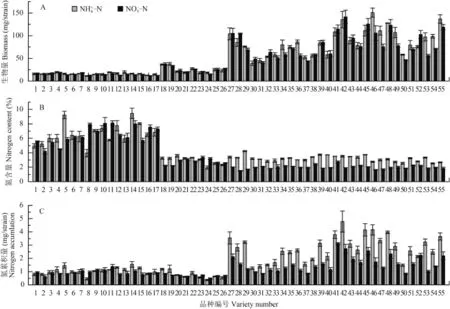
图1 在 NH4+-N 和 NO3--N 培养条件下水稻苗期生物量 (A)、氮含量 (B) 和氮累积量 (C)Fig. 1 The biomass (A) , nitrogen content (B) , nitrogen accumulation (C) of rice plants at the seedling stage cultured in NH4+-NNO3--N solution
--N供应条件下,水稻根系形态指标均表现出一定的变异性,变异系数变幅相差较大。在 NH4+-N 培养条件下,水稻根系形态指标变异系数范围为 0.19~0.87,后者为前者的 4.58 倍,变异系数呈根尖数 > 总根长 >交叉数 > 分枝数 > 总根面积 > 总根体积 > 平均根系直径的趋势,其中平均根系直径变异系数最小,为 0.19;而根尖数变异系数最大,为 0.87。在 NO3--N培养条件下,水稻根系形态指标变异系数呈根尖数 >交叉数 > 分枝数 > 总根长 > 总根面积 > 总根体积 >平均根系直径的趋势,其变异范围为 0.21~1.02,后者是前者的 4.86 倍;其中平均根系直径变异系数最小,为 0.21,根尖数的变异系数最大,为 1.02 (表 2)。

表2 不同形态氮素培养条件下水稻苗期根系形态变异特征Table 2 Variation characteristics of root morphology of rice under different nitrogen conditions
2.3 不同形态氮素培养下水稻营养指标与形态指标相关性分析
在 NH4+-N 培养下,水稻植株生物量与平均根系直径呈极显著负相关关系 (表 3),而与总根长、总根面积、总根体积、根尖数、分枝数、交叉数呈极显著正相关关系 (0.76~0.95)。水稻植株氮含量与总根长、总根面积、总根体积、根尖数、分枝数呈现极显著负相关关系 (-0.31~-0.46),而与平均根系直径呈现正相关关系,但相关系数仅为 0.06。植株氮累积量除与平均根系直径呈显著负相关外 (r = -0.27),与其他根系形态指标皆呈现极显著正相关关系(0.71~0.89)。
在 NO3
--N 培养条件下,水稻植株生物量与平均根系直径呈极显著负相关关系 (表 4),而与总根长、总根面积、总根体积、根尖数、分枝数、交叉数呈极显著正向关 (0.62~0.89)。水稻植株氮含量与总根长、总根面积、总根体积、根尖数、分枝数、交叉数呈现极显著负相关关系 (-0.34~-0.56),而与平均最优回归模型分别为:
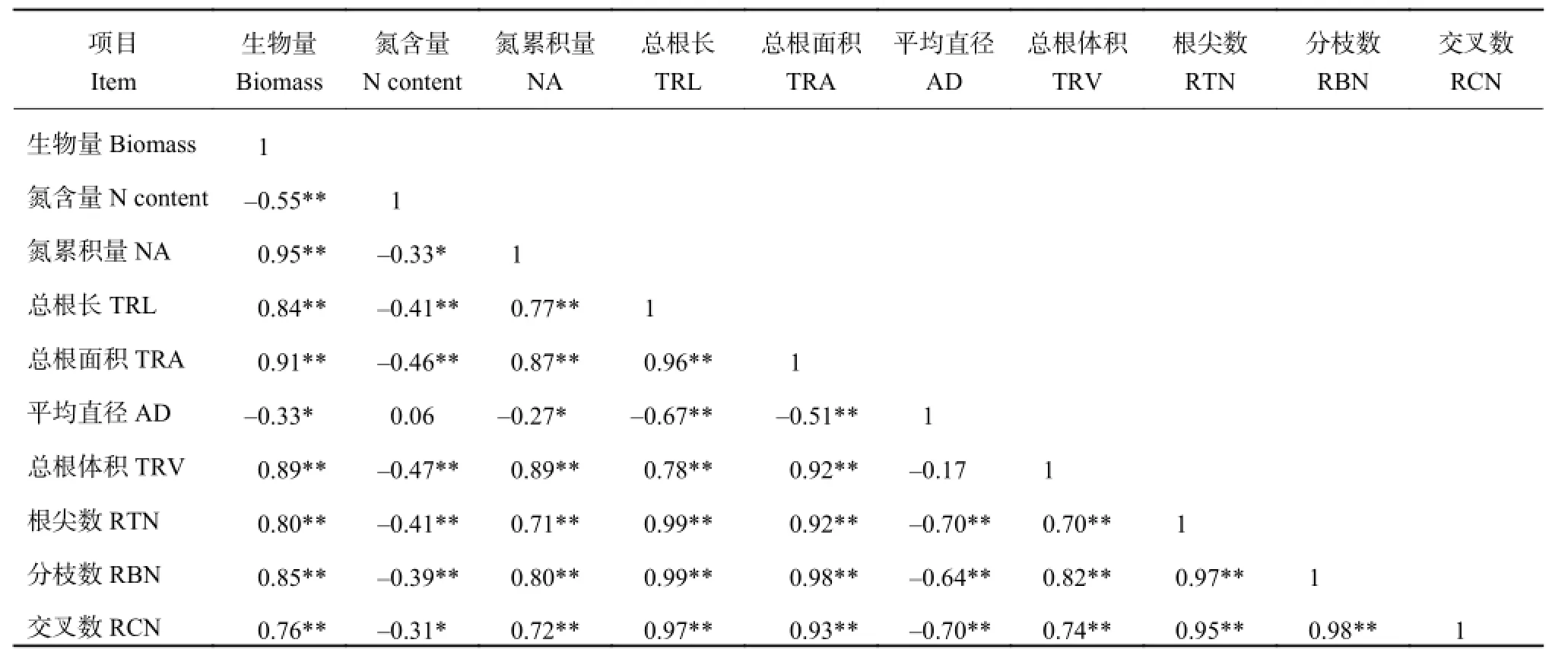
表3 NH4+-N 培养下整株水稻营养指标与根系形态指标相关性分析Table 3 Correlations between the nutritional indices and root morphological indices in the NH4+-N solution
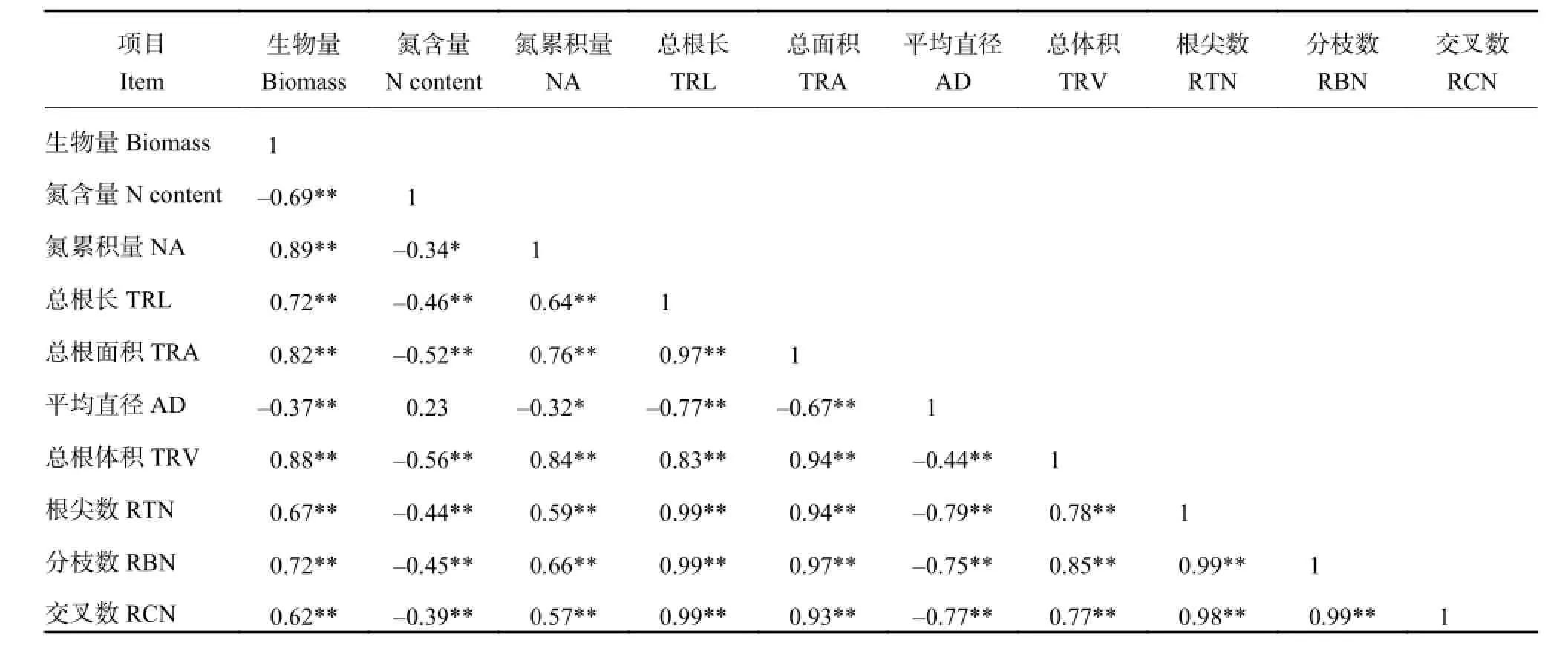
表4 NO3--N 培养下整株水稻营养指标与根系形态指标相关性分析Table 4 Correlation between the nutritional indices and root morphological indices in the NO3--N solution
Y1= - 4 × 10-3+ 3.9E-5X1+ 2 × 10-3X2- 6.74E-6X7
Y2= 1.16 - 7.91 × 10-2X4
Y3= - 0.16+3 × 10-3X1+ 0.07X2
在 NO3--N 培养条件下,水稻植株生物量 (Y1)、氮含量 (Y2) 和氮累积量 (Y3) 与根系形态指标之间的最优回归模型分别为:
Y4= 5 × 10-3+ 1.05 × 10-2X4
Y5= 1.04 - 0.77X4
Y6= 0.51+1.72X4
其中,在 NH4+-N 培养条件下,水稻不同品种根系形态指标中以总根长、总根面积、交叉数对植株生物量的影响最大;以总根体积对植株氮含量的影根系直径呈现正相关关系。植株氮累积量除与平均根系直径呈显著负相关外 (r = -0.32),与其余形态指标皆呈现极显著正相关关系 (0.64~0.84)。
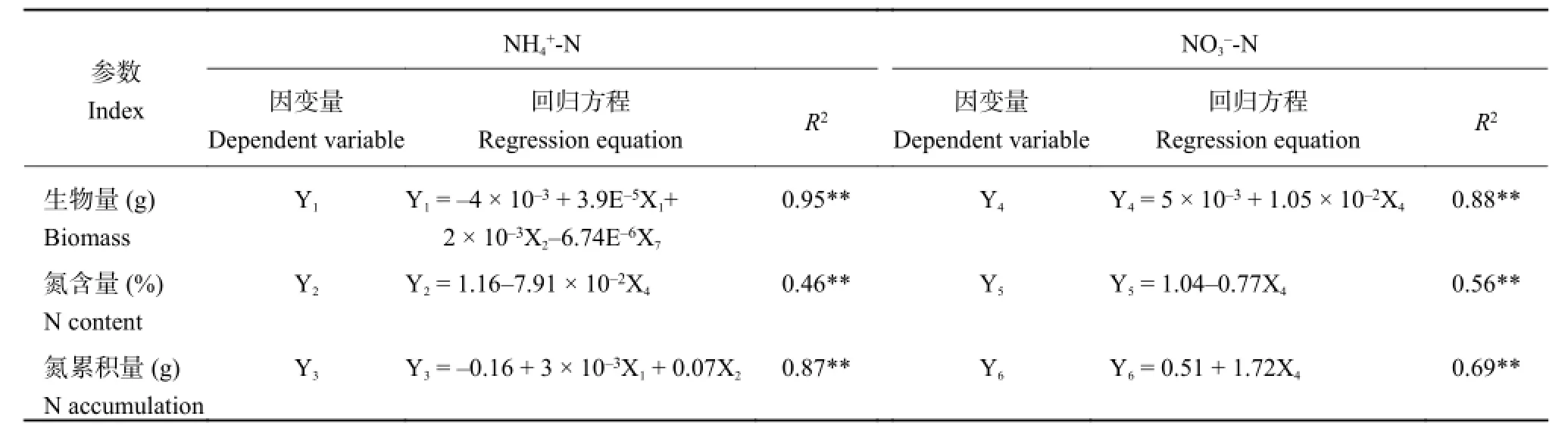
表5 不同形态氮素培养下水稻植株营养指标与形态指标多元线性回归分析Table 5 Multiple linear regression of rice nutritional indices and morphological indices under different nitrogen solution
2.4 不同形态氮素培养下水稻营养指标与形态指标多元线性回归分析
不同形态氮素培养条件下水稻植株生物量、氮含量和氮累积量与根系形态指标多元线性逐步回归方 程 中 ,Xi对 Yi的 决 定 系 数 皆 极 显 著 相 关 (表 5),其在 NH4
+-N 培养条件下的决定系数 (R2) 范围为0.46~0.95,高于其在 NO3--N 培养下的决定系数 (R2)范围 0.56~0.88。
在 NH4+-N 培养条件下,水稻植株生物量 (Y1)、氮含量 (Y2) 和氮累积量 (Y3) 与根系形态指标之间的响最大;以总根长、总根面积对植株氮累积量的影响最大,且均达到了极显著水平。而在 NO3--N 培养条件下,水稻不同类型形态指标均以总根体积对植株生物量、氮含量、氮累积量的影响最大,且均达到极显著水平。
3 讨论
根系是水稻吸收氮素的主要器官,根长、根系表面积等参数在水稻获取氮素方面起着重要作用,是筛选水稻不同氮效率品种的重要指标[10]。水稻高效氮素吸收与其根系形态关系极为密切,良好的根系形态是水稻氮高效的重要基础。水稻氮高效型品种的根系特征包括发根力强、根系发达;根生物量、根系体积、根系吸收表面积和活性吸收面积大;根系氧化力高[11],即氮高效型水稻品种可通过发育良好的根系形态来提高根系对氮素的吸收和利用能力,从而提高地上部氮素含量和氮素积累量[12]。氮高效型水稻根长、根表面积、根体积和根密度显著大于氮低效型[2,10]。杨肖娥等[13]研究指出,水稻根系对氮素的吸收能力较强主要与其根系的形态和生理特性有关。水稻根系长、体积大、分布密度和有效吸收面积大,有利于根系与耕层及底层土壤接触,提高其对土壤中氮的吸收能力。根长和根表面积可能是影响水稻根系吸收氮的重要根系指标,是根系遗传性状改良的重要参数[14]。本研究表明,水稻根系形态中的总根体积、总根面积、总根长、分枝数四个形态指标与植株生物量、植株氮含量、植株氮累积量的相关性最为显著,可作为水稻苗期氮高效评价的重要指标。
根系吸氮效率不仅与根系形态及品种的遗传特性 有 关 , 同 时 也 决 定 于 氮 素 形 态[15-19]。 土 壤 中 的 氮95%~98% 以上呈有机态存在,水溶性和交换性氮含量较少。土壤中的无机氮主要为 NH4+-N 和 NO3--N。淹水条件下硝化作用被强烈抑制,使 NH4+-N 成为水
稻田的主要氮素形态[20-26]。何文寿等[27]和 Cao 等[28]的研究表明,在 NH4+-N 培养条件下能提高硝态氮敏感型水稻体内氮同化酶 (硝酸还原酶和谷氨酰胺合成酶) 的活性,这更利于水稻对氮的吸收[27-28],而 NO3--N吸收的受阻可能与硝酸还原酶的活性受到抑制有关[29-30]。石英等[31]研究也表明,在淹水条件下,水稻呼吸代谢酶以细胞色素氧化酶为主,有利于水稻对铵态氮的吸收。与供应硝态氮 (NO3--N) 相比,供应铵态氮 (NH4
+-N) 可通过提高水稻伤流液、韧皮部汁液的游离氨基酸和 K+含量,显著增强其渗透调节能力 , 从 而 提 高 了 对 铵 态 氮 的 吸 收[32-33]。 本 研 究 也 证实,单供 NH4+-N 的根系的吸氮量要高于单供 NO3--N,说明 NH4+-N 培养下的水稻具有优良的氮吸收累积能力,能更好的被水稻吸收,这与水稻是喜铵植物的结论是一致的。
赵满兴等[34]提出氮高效品种应同时具有较高的氮素吸收能力、较强的氮素转运能力和再转移能力。权 太 勇 等[35]研 究 认 为 水 稻 各 生 育 期 氮 素 吸 收 比 例 不同,移栽至抽穗期占 24%~32%,幼穗形成期至齐穗期占 57%~69%,齐穗期到成熟期占 5.7%~10%。可见水稻苗期的氮素利用效率只是整个生育期的氮素效率的一部分。裴雪霞等[36]研究发现,水培条件下作物苗期相对植株干重与大田条件下相对籽粒产量之间呈极显著的正相关。这说明水稻苗期的根系形态和氮素营养特征对整个生育期的氮素利用效率和籽粒产量均产生重要影响。本研究初步研究了不同氮素培养下水稻苗期的营养指标及其与形态指标的相关性,以期为水稻氮高效品种筛选的根系形态学研究提供科学参考,研究结果尚需在田间试验条件下进行验证,以提高试验结果的可靠性。
4 结论
在 NH4+-N 培养条件下,水稻营养指标与根系形态指标的相关性要高于其在 NO3--N 培养条件下的。在两种不同氮素 (NH4+-N 和 NO3--N) 培养下,55 个供试水稻品种的植株干重、植株氮累积量与总根长、总根面积、总根体积、总根体积、根尖数、分枝数、交叉数等指标存在显著性差异,其中以总根体积、总根面积、总根长、分枝数四个指标的差异性最为显著。水稻根系形态中的总根体积、总根面积、总根长、分枝数四个形态指标与植株生物量、植株氮含量、植株氮累积量的相关性最为显著,可作为水稻苗期氮高效评价的重要指标。
[1]杨建昌. 水 稻 根 系形态生理与产量 、 品质形成及养分吸 收 利 用的关系[J]. 中国农业科学, 2011, 44(1): 36-46. Yang J C. Relationships of rice root morphology and physiology with the formation of grain yield and quality and the nutrient absorption and utilization [J]. Scientia Agricultura Sinica, 2011, 44(1): 36-46.
[2]魏海燕, 张洪程, 张胜飞, 等. 不同氮利用效率水稻基 因型的根系形态与生理指标研究[J]. 作物学报, 2008, 34(3): 429-436. Wei H Y, Zhang H C, Zhang S F, et al. Root morphological and physiological characteristics in rice genotypes with different N use efficiencies [J]. Acta Agronomica Sinica, 2008, 34(3): 429-436.
[3]李勇, 周毅, 郭世伟, 沈其荣. 铵态氮和硝态氮营养对 水、旱稻根系形态及水分吸收的影响[J]. 中国水稻科学, 2007, 21(3): 294-298. Li Y, Zhou Y, Guo S W, Shen Q R. Effects of different N forms on root morphology and water absorption of lowland and upland rice plants [J]. Chinese Journal of Rice Science, 2007, 21(3): 294-298.
[4]巨 晓棠, 谷 保静. 我国 农田氮肥施 用现状、问题 及趋势[J]. 植 物营养与肥料学报, 2014, 20(4): 783-795. Ju X T, Gu B J. Status-quo, problem and trend of nitrogen fertilization in China [J]. Journal of Plant Nutrition and Fertilizer, 2014, 20(4): 783-795.
[5]朱 兆 良. 稻 田 节 氮 的 水 肥 综 合 管 理 技 术 的 研 究[J]. 土 壤, 1991, 23(5): 241-245. Zhu Z L. Study on the integrated management of water and fertilizer in paddy field of nitrogen saving technology [J]. Soils, 1991, 23(5):241-245.
[6]张福锁, 王激清, 张卫峰, 等. 中国主要粮食作物肥料利用率现状与提高途径[J]. 土壤学报, 2008, 45(5): 915-924. Zhang F S, Wang J Q, Zhang W F, et al. Current situation and improvement of fertilizer utilization rate of main grain crops in China [J]. Acta Pedologica Sinica, 2008, 45(5): 915-924.
[7]李宝珍, 王松伟, 冯慧敏, 等. 氮素供应形态对水稻根系形态和磷吸收的影响[J]. 中国水稻科学, 2008, 22(5): 665-668. LI B Z, Wang S W, Feng H M, et al. Effects of nitrogen forms on root morphology and phosphate uptake in rice [J]. Chinese Journal of Rice Science, 2008, 22(5): 665-668.
[8]戢林, 李廷轩, 张锡洲, 等. 氮高效利用基因型水稻根系形态和活力特征[J]. 中国农业科学, 2012, 45(23): 4770-4781. Ji L, Li T X, Zhang X Z, et al. Root morphological and activity characteristics of rice genotype with high nitrogen utilization efficiency[J]. Scientia Agricultura Sinica, 2012, 45(23): 4770-4781.
[9]樊 剑 波. 不同氮效率基因型水 稻 氮 素吸收和根系特征研 究[D]. 南京: 南京农业大学博士学位论文, 2008. Fan J B. Difference in nitrogen uptake and root morphology of rice cultivars with different nitrogen use efficiency [D]. Nanjing: PhD Dissertation, Nanjing Agricultural University, 2008.
[10]樊剑波, 沈其荣, 谭炯壮, 等. 不同氮效率水稻品种根系生理生态指标的差异[J]. 生态学报, 2009, 29(6): 3052-3058. Fan J B, Shen Q R, Tan J Z, et al. Difference of root physiological and ecological indices in rice cultivars with different N use efficiency [J]. Acta Ecologica Sinica, 2009, 29(6): 3052-3058.
[11]程建峰, 戴廷波, 荆奇, 等. 不同水稻基因型的根系形态生理特性与高效氮素吸收 [J]. 土壤学报, 2007, 44(2): 266-272. Cheng J F, Dai T B, Jin Q, et al. Root morphological and physiological characteristics of different rice genotypes and high efficiency of nitrogen uptake [J]. Acta Pedologica Sinica, 2007, 44(2): 266-272.
[12]Ladha J K, Kirk G J D, Bennett J, et al. Opportunities for increased nitrogen use efficiency from improved lowland rice germplasm [J]. Field Crops Research, 1998, 56(1-2): 41-71.
[13]杨肖娥, 孙羲. 不同水稻品种对低氮反应的差异及其机制的研究[J]. 土壤学报, 1992, 29(1): 73-79. Yang X E, Sun X. Studies on the differences and mechanism of different rice varieties on the response to low nitrogen [J]. Acta Pedologica Sinica, 1992, 29(1): 73-79.
[14]赵学强, 施卫明. 水稻根系生长对不同氮形态响应的动态变化[J].土壤, 2007, 39(5): 766-771. Zhao X Q, Shi W M. Dynamic changes of response of rice root growth to different nitrogen forms [J]. Soils, 2007, 39(5): 766-771.
[15]张辰明, 徐烨红, 赵海娟, 等. 不同氮形态对水稻苗期氮素吸收和根系生长的影响[J]. 南京农业大学学报, 2011, 34(3): 72-76. Zhang C M, Xu Y H, Zhao H J, et al. Effects of different nitrogen forms on nitrogen uptake and root growth of rice at the seedling stage [J]. Journal of Nanjing Agricultural University, 2011, 34(3): 72-76.
[16]李素梅, 施卫明. 不同氮形态对两种基因型水稻根系形态及氮吸收效率的影响[J]. 土壤, 2007, 39(4): 589-593. Li S M, Shi W M. Effects of different nitrogen forms on root morphology and nitrogen uptake efficiency of two rice genotypes [J]. Soils, 2007, 39(4): 589-593.
[17]Radin J W. Differential regulation of nitrate reductase induction in roots and shoots of cotton plants [J]. Plant Physiology, 1975, 55(2):178-182.
[18]Zhang H, Xue Y G, Wang Z Q, et al. Morphological and physiological traits of roots and their relationships with shoot growth in “super” rice [J]. Field Crops Research, 2009, 113(1): 31-40.
[19]Himmelbauer M, Loiskandl A, Kastanek A. Estimating length average diameter and surface area of roots using two different image analyses systems [J]. Plant and Soil, 2004, 260(1): 111-120.
[20]Courtois B, Ahmadi N, Khowaja F, et al. Rice root genetic architecture: meta-analysis from a drought QTL database [J]. Rice, 2009, 2(2): 115-128.
[21]Galkovskyi T, Mileyko Y, Bucksch A, et al. Gia roots: software for the high throughput analysis of plant root system architecture [J]. BMC Plant Biology, 2011, 12(1): 1-12.
[22]Kiry J G D, Du L V. Changes in rice root architecture, porosity, and oxygen and proton release under phosphorus deficiency [J]. New Phytologist, 1997, 135(2): 191-200.
[23]Pang W, Crow W T, Luc J E, et al. Comparison of water displacement and WinRHIZO software for plant root parameter assessment [J]. Plant Disease, 2011, 95(10): 1308-1310.
[24]Satoshi O, Milton O V, Manabu I, et al. Root system architecture variation in response to different NH4+concentrations and its association with nitrogen-deficient tolerance traits in rice [J]. Acta Physiology Plant, 2014, 32(9): 355-364.
[25]Dai J L, Duan L S, Dong H Z. Improved nutrient uptake enhances cotton growth and salinity tolerance in saline media [J]. Journal of Plant Nutrition, 2014, 37(8): 1269-1286.
[26]Darrah P R, Jones D L, Kirk G J D, et al. Modelling the rhizosphere:a review of methods for ‘upscaling’ to the whole-plant scale [J]. European Journal of Soil Science, 2006, 57(1): 13-25.
[27]何文寿, 李生秀, 李辉桃. 水稻对铵态氮和硝态氮吸收特性的研究[J]. 中国水稻科学, 1998, (4): 249-252. He W S, Li S X, Li H T. Characteristics of ammonium and nitrate uptake in rice [J]. Chinese Journal of Rice Science, 1998, (4):249-252.
[28]Cao Y, Fan X R, Sun S B, et al. Effect of nitrate on activities and transcript levels of nitrate reductase and glutamine synthetase in rice [J]. Pedosphere, 2008, 18(5): 664-673.
[29]Radin J W. Differential regulation of nitrate reductase induction in roots and shoots of cotton plants [J]. Plant Physiology, 1975, 55(2):178-182.
[30]Mackown C T, Jackson W A, Volk R J. Restricted nitrate influx and reduction in corn seedlings exposed to nitrate and ammonium nutrition [J]. Plant Physiology, 1982, 62(9): 353-359.
[31]石英, 沈其荣, 茆泽圣, 等. 旱作水稻根际土壤铵态氮和硝态氮的时空变异[J]. 中国农业科学, 2002, 35(3): 520-524. Shi Y, Shen Q R, Mao Z S, et al. Time and horizontal spatial variations of NH4+-N and NO3--N of rhizospheric soil with rice cultivation on upland condition mulched with half-decomposed rice straw [J]. Scientia Agricultura Sinica, 2002, 35(3): 520-524.
[32]Li Y, Gao Y X, Ding L, et al. Ammonium enhances the tolerance of rice seedlings (Oryza sativa L.) to drought condition [J].Agricultural Water Management, 2009, 96(12): 1746-1750.
[33]栗海俊, 李勇, 杨秀霞, 等. 不同形态氮素营养和水分条件对苗期水稻生长及渗透调节能力的影响[J]. 中国水稻科学, 2010, 24(4):403-409. Su H J, Li Y, Yang X X, et al. Effects of different nitrogen forms and water stress on the growth and osmotic adjustment of rice seedlings [J]. Chinese Journal of Rice Sciences, 2010, 24(4): 403-409.
[34]赵满兴, 周建斌, 杨绒, 等. 不同施氮量对旱地不同品种冬小麦氮素累积、运输和分配的影响[J]. 植物营养与肥料学报, 2006, 12(2):143-149. Zhao M X, Zhou J B, Yang R, et al. Characteristics of nitrogen accumulation, distribution and translocation in winter wheat on dryland [J]. Plant Nutrition and Fertilizer Science, 2006, 12(2):143-149.
[35]权太勇, 金妍姬, 韩云哲. 水稻不同群体的氮素吸收特性[J]. 延边大学农学学报, 2000, 6(2): 86-90. Quan T Y, Jin Y J, Han Y Z. Nitrogen uptake characteristics of different rice populations [J]. Journal of Agricultural Science Yanbian University, 2000, 6(2): 86-90.
[36]裴雪霞, 王姣爱, 党建友, 等. 耐低氮小麦基因型筛选指标的研究[J]. 植物营养与肥料学报, 2007, 13(1): 93-98. Pei X X, Wang J A, Dang J Y, et al. An approach to the screening index for low nitrogen tolerant wheat genotype [J]. Plant Nutrition and Fertilizer Science, 2007, 13(1): 93-98.
Correlation between root morphology and nitrogen uptake of rice
CHEN Chen, GONG Hai-qing, ZHANG Jing-zhi, GAO Hong-jian*
( College of Resources and Environment, Anhui Agricultural University, Hefei 230036, China )
【Objectives】Excessive application of nitrogen fertilizers lead to nitrogen loss and the cost increase of agricultural production. Root morphology is one of main factors that affect absorption and utilization of nutrients by rice. In this study, we investigated the relationship between root morphology and nitrogen uptake and accumulation of rice.【Methods】Hydroponic experiments were conducted with 55 rice cultivars to examine relationships between the plant nitrogen uptake and several indices of root morphology using both NH4+-N and NO3--N solution.【Results】The correlation coefficients between the nitrogen uptake and root morphology indices when N was supplied as NH4+-N were higher than those when supplied as NO3--N. After the six leaf stage of rice, under the same solution concentration of nitrogen, the average biomass of rice seedlings was 55.77 mg per plant in the NH4+-N solution, which was increased by 4.94 mg per plant compared with the NO3--N solution. The average nitrogen content of rice seedlings was 4.22% when grown in the NH4+-N solution, which was increased by 0.72%, and the average accumulation of nitrogen in the seedlings was 1.91 mg per plant when supplied with the NH4+-N, which was increased by 0.67 mg per plant compared with the NO3--N solution. There were significant differences in the root morphology indices between plants supplied with the NH4+-N and NO3--N under the same concentration. The higher the coefficient of variation of root morphology was, the higher the greater difference between rice cultivars was. The root branch number was most highly correlated with the root morphology indices, whereas the mean root diameter was the least correlated. These differences reflected that there was a largervariation in root branching values, and less variation in mean root diameters among the 55 rice cultivars. The total root volume, total root area, total root length and branching number were significantly correlated with biomass, nitrogen content, and nitrogen accumulation in rice plants at the seedling stage, and the correlation coefficients were in order: the total root volume > the total root area > total root length > the number of branches. The correlations between the nitrogen uptake and the indices of root morphology when plants were supplied with NH4+-N were higher than those when supplied with NO3--N.【Conclusions】The total root volume, total root area, total root length, and the number of branches may be important indicators for evaluating the nitrogen use efficiency of rice at the seedling stage.
rice; root morphology; biomass; nitrogen accumulation; correlation analysis
2016-04-12 接受日期:2016-06-23
国家自然科学基金(31328020);国家科技支撑计划项目(2013BAD07B08,2012BAD04B09)资助。
陈晨(1992—),女,安徽天长市人,硕士,主要从事植物养分高效吸收方面的研究。E-mail:chenchenahau@163.com
* 通信作者 E-mail:gaohongjian2@163.com
