川渝黔稻瘟病菌毒性的区域分化研究
2017-04-13张姝钟雪莲乔贵艳沈丽周天云彭云良
张姝,钟雪莲,乔贵艳,沈丽,周天云,彭云良*
(1.四川省农业科学院植物保护研究所/农业部西南作物有害生物综合治理重点实验室,四川成都610066;2.四川省农业厅植物保护站,四川成都610042;3.重庆市种子管理站,重庆401121)
川渝黔稻瘟病菌毒性的区域分化研究
张姝1,钟雪莲1,乔贵艳1,沈丽2,周天云3,彭云良1*
(1.四川省农业科学院植物保护研究所/农业部西南作物有害生物综合治理重点实验室,四川成都610066;2.四川省农业厅植物保护站,四川成都610042;3.重庆市种子管理站,重庆401121)
四川、重庆和贵州是我国稻瘟病(Magnaporthe oryzae)重要发生区之一,分析这些地区稻瘟病菌群体的毒性差异,有助于延长抗性品种使用寿命,减轻稻瘟病发生危害。为了解不同地区稻瘟病菌的毒性差异,2012-2013年在四川、重庆21个县(区)的稻瘟病常发地种植以丽江新团黑谷为背景的24个水稻抗稻瘟病单基因系,按国际水稻所0~9级标准调查各地水稻抗稻瘟单基因系的发病情况。2014-2015年采集四川省北部10个、贵州省全境15个和山东省2个县(区)共47块病田穗颈瘟标样,浸湿后接种上述近等基因系及16个水稻品种并以薄膜覆盖隔离40 d后调查。四川、重庆21个病圃的调查结果表明在不同地点表现抗病的频率最高的基因为Pi2(38.10%)。四川、贵州等地标样接种鉴定结果表明Pikh、Piks和PiKm对来自贵州28块病田的稻瘟菌的抗性频率分别为51.85%、48.15%和48.15%,PiKm、Pi2、Pi9、Pikh、和Piks对来自四川省北部18块病田的病菌的抗性频率分别为70.00%、60.00%、50.00%、50.00%和45.00%,其中Pi2对四川北部病菌的抗性频率显著高于贵州病菌(P=0.0066)。四川稻瘟病菌生理小种鉴别寄主成恢448、内恢99-14和RHR-1对贵州病田来源病菌的抗性频率分别为74.07%、70.37%和70.37%,对四川北部病田来源病菌的抗性频率则分别为90.00%、70.00%和90.00%,均显著高于IR24、明恢63、多恢1号对两地病菌的抗性频率。10个杂交稻生产品种对四川北部和贵州省病田稻瘟病菌的抗性频率分别在90.00%~100.00%和81.48%~92.59%之间波动。对不同病圃和病田的稻瘟病菌毒性的聚类分析结果表明,稻瘟病菌的毒性差异虽然有随着地理距离扩大而加大的趋势,但相同县乃至同一地点不同田块间病菌毒性也可能有显著差异,而不同县(区)的稻瘟病菌则可能因对24个单基因系同时具有毒性而须借助其他水稻品种加以区分。
稻瘟病菌;抗稻瘟病单基因系;抗性频率;菌株毒性;聚类分析
由子囊菌[Magnaporthe oryzae(Hebert)Barr.]引起的稻瘟病是全世界范围内水稻生产上最具毁灭性的病害之一,一般流行年份病害引起减产20%~30%,重病田块甚至绝收[1]。选育和利用抗病品种,是控制稻瘟病最经济、有效和安全的策略,但是由于稻瘟病菌群体毒性组成复杂并不断演化,抗病品种大面积种植3~5年后即丧失抗病作用,已成为抗稻瘟病品种选育和利用中的瓶颈问题。因此,对稻瘟病菌群体毒性的时空变化进行监测,对于合理布局抗病品种,延长抗病品种使用寿命,具有重要意义。据不完全统计,四川、重庆和贵州3省(市)水稻面积超过400万hm2,水稻叶瘟、穗瘟发生面积在50~100万之间波动,是我国稻瘟病严重发生区域之一,水稻抗稻瘟病品种丧失抗性在四川、重庆和贵州屡有发生[2-7]。除少数品种外,多数品种很少普遍表现抗病[8-9]。本研究在四川、重庆和贵州多点设置病圃,种植以丽江新团黑谷为背景的抗稻瘟病单基因系,并采集不同地点穗颈瘟标样在室内直接产孢病接种上述近等基因系和其他水稻生产品种,以分析不同地点稻瘟病菌群体的毒性差异。通过对四川、重庆和贵州地区稻瘟病菌群体间菌株毒性差异变化的研究,以期为进一步设置病菌毒性监测圃和大面积生产上品种合理布局提供依据。
1 材料与方法
1.1 水稻抗稻瘟病单基因系和品种
24个以丽江新团黑谷为背景的抗稻瘟病单基因新系(表1)系国际水稻研究所(IRRI)和日本专家合作选育而成[10],由美国Ohio州立大学王国亮教授提供,四川省农业科学院植物保护研究所保存和繁殖。水稻稻瘟病菌生理小种鉴别寄主IR24、明恢63、多恢1号、成恢448、内恢99-14和RHR-1[5]由四川省农业科学院植物保护研究所收集、繁殖和保存。10个抗稻瘟病生产品种商品种子则收集于四川省成都、南充和内江等地种子市场,并依张雪梅等已报道方法[8]在四川邛崃设置病圃中表现抗病。
1.2 病圃设置
2012-2013年间在四川、重庆稻瘟病各县(市) (表1)稻瘟病常发区设立病圃,将各单基因系播种,播种时间较当地大面积生产播种时间推迟7~14 d,于5月上、中旬将秧苗按照16.5 cm×23.1 cm规格移栽于大田病圃中,每个品种设3次重复,每个重复移栽3行,每行10株,品种呈随机区组排列。按照常规方法防治水稻虫害、杂草和其他病害,但不施用防治稻瘟病的药剂。
1.3 标样采集
2014-2015年8-9月水稻黄熟期前,到四川位于四川省北部的安县、盐亭、南部、蓬安、营山、渠县、大竹、达川、宣汉、巴州、仪陇等10个县(区)、贵州省遵义、黔西、金沙、赫章、兴仁、兴义、安龙、长顺、花溪、贵定、都匀、麻江、沿河、天柱等14个县(区)、以及山东济南市槐荫区、济宁市任城区的稻瘟病发生区采集穗颈瘟标样,各采样点选取1块或不同品种的多块病田,共计47块病田进行采样;采样时沿病田对角线和稻田四周采取120~150穗病穗,去除谷粒后剪取10~12 cm发病穗颈,风干后放入含变色硅胶的容器内,室温下干燥保存备用。
1.4 室内播种、育苗及诱发接种
10月底将抗稻瘟病单基因系、恢复系和生产品种用甲基纤维素钠溶液固定、排列于双层草纸间,播种于室内水泥池内湿润育秧苗床表面,各品种播种5~10粒,重复2次,2小区四周播种丽江新团黑谷作诱发行。出苗后0.4~1.0 cm时喷施50%百菌清600倍稀释液一次预防立枯病,待幼苗生长至2叶1心期时按10 g/m2施用尿素1次,并将保存的穗颈瘟标样用自来水浸泡4 h,捞出沥干后撒放在两小区之上,架设竹拱,覆盖薄膜将不同地点和田块来源标样隔离,用细土将薄膜四周压实以防交叉侵染直至调查。
1.5 病害调查
田间病圃在7月上、中旬进行,各重复调查100片叶片,按国际水稻所0~9级标准记载叶瘟级别[11]。室内试验调查在接种后40 d、丽江新团黑谷株发病率达到100%时进行,调查各单基因系和品种的株发病率和最高叶瘟级别。
1.6 数据处理
将各单基因系和品种各重复最高叶瘟级别进行归总,将各重复中叶瘟最高级别在2级以下者记为抗病(R),叶瘟级别最高达3级记为中感(MS),3级以上者记为感病(S),分析各单基因系和品种中表现抗病和感病的频率。将MS、S赋值1,R反应赋值为0后,利用Statistica软件进行聚类分析。
2 结果与分析
2.1 水稻抗稻瘟基因在四川和重庆田间抗性表现
24个抗稻瘟病单基因系、丽江新团黑谷在四川省和重庆市21个病圃中对稻瘟病的抗感表现如表1。不同单基因系在各地的抗性表现具有显著差异,其中IRBLb(Pib)、IRBLt-K59(Pit)、IRBL12-M[Pi12 (t)]、IRBL19-M[Pi19(t)]及对照丽江新团黑谷在各个病圃均表现感病,表现抗病的频率为0.00;在多数病圃表现感病、抗性频率较丽江新团黑谷无显著差异的有IRBLa-A(Pia)、IRBLi-F5(Pii)、IRBLks-F5(Pi ks)、IRBLk-Ka(Pik)、IRBLkp-K60(Pikp)、IRBLz-Fu(Piz)、IRBLta-K1(Pita)、IRBLsh-B(Pish)、IRBL1-CL(Pi1)、IRBL3-CP4(Pi3)、IRBL5-M[Pi5 (t)]、IRBL7-M[Pi7(t)]、IRBL20-IR24(Pi20)和IRBL11-Zh(Pi11)等14个单基因系。在4个以上病圃表现抗病、抗病频率显著高于感病对照的有分别含PiKh、Pi zt、Pi9、Pita2、Pikm、和Pi2等基因的6个单基因系,其中Pi2表现抗病的频率最高,达38.10 %。从表1可看出,除在各地均表现感病和部分仅在上述病圃其中一个病圃表现抗病的抗稻瘟病单基因系外,大部分单基因系在不同病圃的抗性表现均有差异。
2.2 四川和重庆不同病圃稻瘟病菌群体毒性的聚类分析
由表1可见,除四川省蒲江、仪陇、重庆市万县、南川和巴南病圃中稻瘟病菌对所有抗稻瘟单基因系均能致病外,不同地点稻瘟病菌群体对各单基因系的毒性不同。聚类分析结果表明(图1),在欧氏距离1.0 h,至少对23个单基因系致病的四川省雅安、蒲江、安县、仪陇、达县、通江、巴州、重庆市万县、南川、巴南、梁平、垫江和合川等13个病圃聚合为一类,其余病圃均单独成为独特类型,其中历年稻瘟病发生面积较小的四川射洪、重庆江津病圃稻瘟病菌毒性最弱,与其他病圃差异最大。
2.3 水稻抗稻瘟基因、抗性恢复系和生产品种对四川北部和贵州病田稻瘟病菌的抗性
将采自贵州省27块稻瘟病田和四川省北部18块病田的穗颈瘟标样充分浸湿后,接种24个抗稻瘟病单基因系、6个四川稻瘟病菌鉴别寄主和10个前期在邛崃病圃表现抗病的杂交稻组合,薄膜覆盖隔离,24 h后标样上即检测到分生孢子,7 d后透过薄膜开始观察到病斑,40 d后揭膜进行调查结果如表2。在抗病单基因系中,含有Pia、Pit、Pib、Pii、Pish、Pi1、Pi3、Pi19(t)和Pi20等单基因系对来自贵州或四川病田的稻瘟病菌的抗性频率与丽江新团黑谷无显著差异,Piks、Pikh、PiKm对贵州病田稻瘟病菌抗性频率分别为48.15%、51.85%和48.15%,显著高于除Pi(7)、Pi11外的其他单基因系。在对来自四川省北部病田的病菌抗性方面,以PiKm的抗性频率(70.00%)最高,而Piks、Pikh、Pi2和Pi9的抗性频率分别为45.00%、50.00%、60.00%和50.00 %。通过t测验发现,Pi2对四川北部病田稻瘟病菌的抗性频率显著高于对贵州病菌的抗性频率(P= 0.0066)。6个四川水稻抗性恢复系中,成恢448、内恢99-14和RHR-1对贵州病田来源病菌的抗性频率分别为74.07%、70.37%和70.37%,显著高于IR24、明恢63、多恢1号和除含Piks、Pikh、PiKm外的单基因系。成恢448、内恢99-14和RHR-1对四川北部病田稻瘟病菌的抗性频率分别为90.00%、70.00%和90.00%,也显著高于IR24、明恢63、多恢1号和除含Piks、Pikh、Pi2和Pi9外的单基因系。在前期已鉴定表现抗病的10个四川杂交稻生产品种中,对贵州省病田稻瘟病菌的抗性频率分别在81.48%~92.59%,显著高于所有抗稻瘟病单基因系。10个抗性生产品种对四川北部病田稻瘟病菌的抗性频率在90.00%~100.00%,显著高于除PiKm之外的抗稻瘟单基因系。3个抗性恢复系和10个生产品种对四川北部和贵州来源病菌的抗性频率无显著差异(P>0.0594)。
2.4 四川北部和贵州病田稻瘟病菌群体毒性的聚类分析
除了来自四川蓬安、贵州赫章和贵定3块病田穗颈瘟标样的稻瘟病菌对所有单基因系均能致病外,各病田稻瘟病菌群体对24个单基因系的毒性均不完全相同。如加上6个四川稻瘟病菌生理小种鉴别品种,则可以将来自四川、贵州45块病田,以及山东济南和济宁各一块病田的稻瘟病菌群体毒性完全区分开。将这47块田病菌对24个单基因系以及6
个鉴别品种的毒性进行聚类分析,发现各病田之间欧氏距离均>1.0,而在欧氏距离=2.0时,47块田病菌毒性可以聚合成23个类群,其中第一类群含有来自四川省北部的8块病田和贵州省不同区域的15块病田(图2),其余类群则均为单成员类群。由图2可以看出,来自相同县(区)的不同田块可以在不同的类群,其中来自遵义市龙坪镇上水村不同品种的遵义-1、遵义-2、遵义市新民镇镇中心村不同品种的遵义-4、遵义-5和遵义-6、沿河县中界乡东流村不同品种的沿河-4、沿河-5、沿河-6均可分布在不同类群中。山东济南和济宁两块病田的病菌毒性与四川、贵州病田病菌毒性差异较大,在欧氏距离≥3.0时才能聚合到一起。
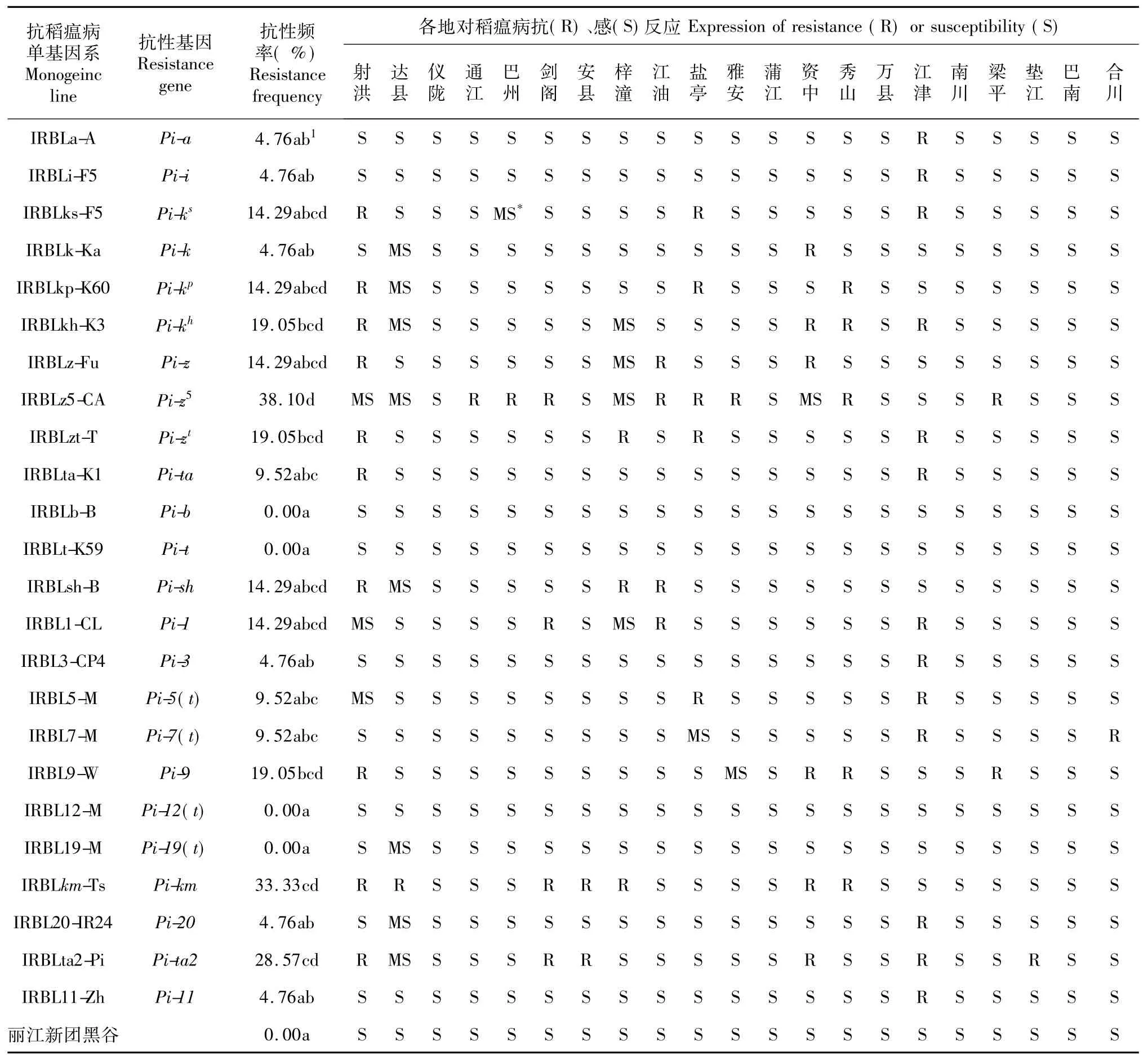
表1 抗稻瘟病单基因系在四川和重庆各地病圃的抗性表现Table 1 Expression of resistance of24 monogenic lines of rice for blast resistance to Magnaportheoryzae atdifferent disease nurseries in Sichuan and Chongqing
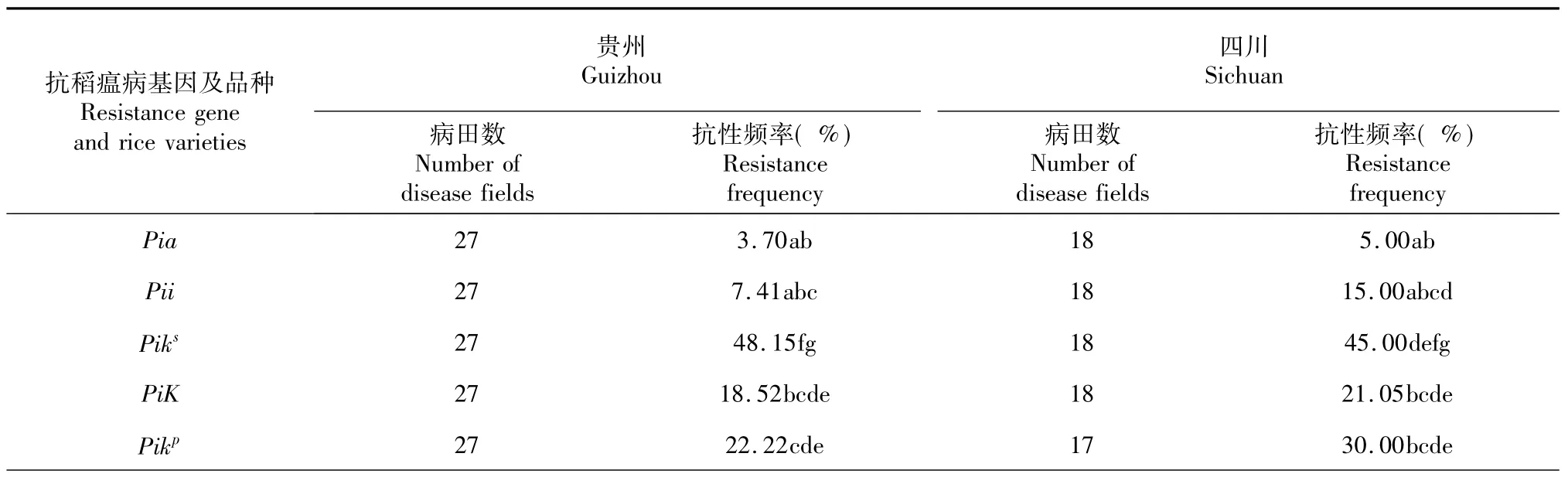
表2 接种贵州和四川不同来源穗颈瘟标样后抗稻瘟病单基因系和品种的抗性表现Table 2 Expression of resistance ofmonogenic lines carrying different resistance genes and rice varieties to Magnaporthe oryzae after inoculation of ear blast specimens of different origins from Guizhou and Sichuan provinces
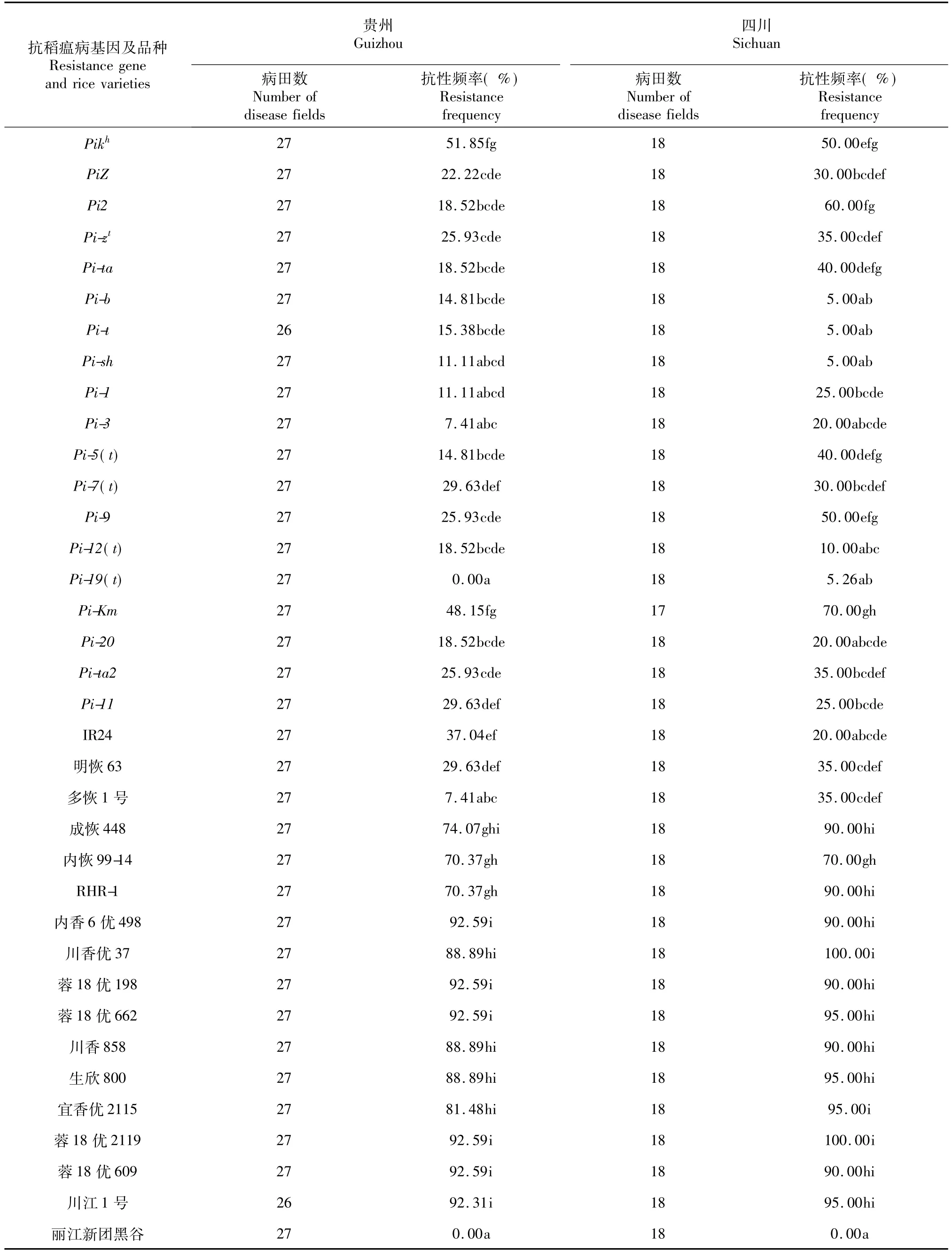
续表2 Continued table 2
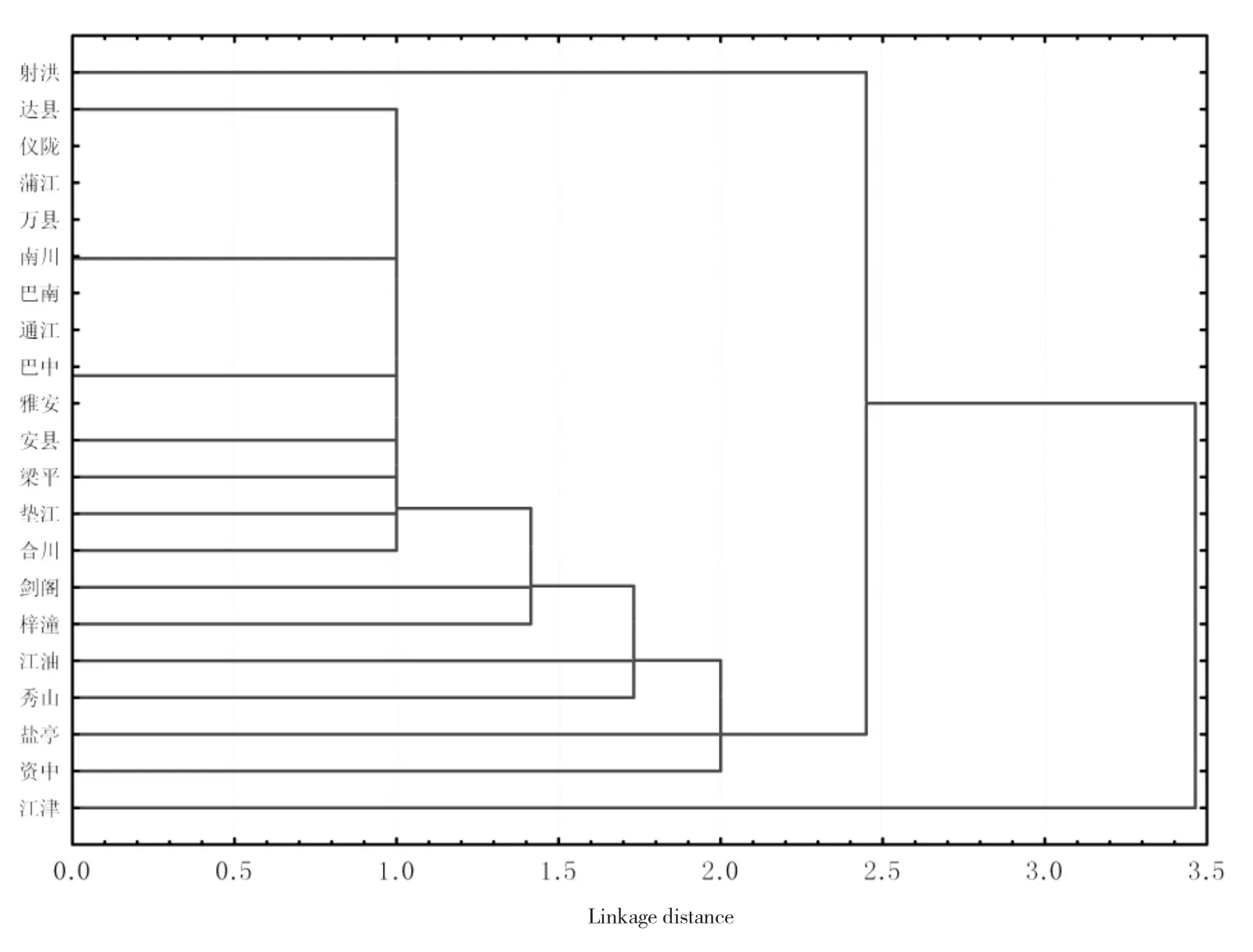
图1 四川和重庆不同县(区)病圃稻瘟病菌对24个抗稻瘟病单基因系毒性的聚类分析Fig.1 Cluster analysis of virulence of Magnaporthe oryzae to 24 monogenic lines of rice for blast ressitance
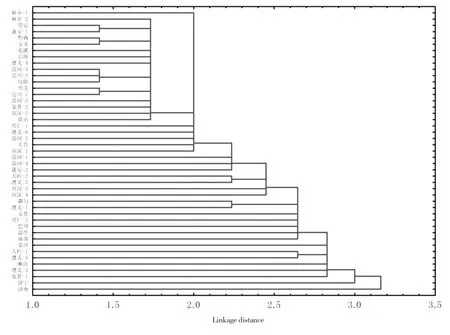
图2 四川北部、贵州省和山东47块病田病菌对30个水稻抗稻瘟病单基因系和品种毒性的聚类分析Fig.2 Cluster analysis of virulence of Magnaportheoryzaeoryzae from 47 paddy fields located in Shandong,Guizhou and northern part of Sichuan province to30 monogenic lines and varieties for blast resistance
3 讨论
众多研究表明,稻瘟病菌群体毒性随时空而变化[1]。例如2011年至2014年间江苏省各稻区稻瘟病菌群体毒性的变化[12],而黑龙江各稻作区土地辽阔稻瘟病菌的毒性分化各不相同[13]。本研究中24个抗稻瘟病单基因系在不同地点的病圃中的发病结果,以及采自不同地区病田穗颈瘟标样所产分生孢子对各单基因系的致病结果,都表明来自四川、重庆和贵州不同地区的稻瘟病菌群体均可能对24个单基因系同时具有毒性,而加入生产品种可以更好地区分不同田块之间的病菌毒性差异。各单基因系在不同地区的抗性表现表明,随着地理距离的进一步扩大,不同区域稻瘟病菌的毒性差异更加明显。总体上Pi2基因在四川省北部的抗性表现优于贵州、而西南地区与山东病田的稻瘟病菌毒性迥异。
不同地区乃至不同田块之间稻瘟病菌毒性的差异,不仅反映了不同地点既往和当前种植水稻品种抗性基因对病菌毒性的筛选作用[14],还为抗性品种筛选病圃的地点设置带来了困难。四川和重庆不同病圃的研究结果表明,稻瘟病常发区的毒性强于非常发区的射洪和江津,在稻瘟病常发区域设置病圃有利于高抗品种的选育。但常发区之间稻瘟病菌毒性仍有差异,因此利用含有不同抗性基因的水稻品种作为指示品种,选择发病品种最多的地点作为抗性品种选育过程中的抗性鉴定圃很有必要。但是有研究发现水稻品种的抗病表型具有明显的地域特征[15],本研究数据也表明,在部分病田表现抗病的生产品种,并不必然在其他病田表现抗病。
4 结论
不同地区稻瘟病菌群体毒性差异较大[16],因此各地在进行抗性品种布局时,应针对本区域的稻瘟病菌群体筛选合适的抗病品种。将不同地区尤其是不同省份之间稻瘟病菌引入本地田间病圃而随气流传播,将不利于抗性品种布局。本研究采集不同地区稻瘟病颈瘟标样在室内产孢、接种不同品种,简化了接种技术,节约了试验时间和成本,研究此筛选结果与品种在生产上的实际表现之间的关系并进一步将此技术规范和推广,将有利于基层农技部门乃至农户筛选抗病品种。
[1]Ou SH.Rice Diseases[M].2nded,Slough,UK:Commonwealth Agricultural Bureau,1985,380.
[2]彭云良,陈国华,杨晓蓉,等.稻瘟病菌对杂交水稻致病菌系组成及其来源研究[J].植物保护学报,1995,22:247-250.
[3]Peng Y L,Chen G H,He M.A possible mechanism for the breakdown of resistance of hybrid rice to blast disease[J].Med.Fac.Landbouww.Univ.Gent,1998,63:947-956.
[4]姬红丽,廖华明,龚爱清,等.四川稻瘟病菌群体致病性变异新趋势[J].西南农业学报,2005,18(3):286-290.
[5]JiH,Shen L,Xiang Y,etal.Chapter23:Studieson the Complementary Differential Varietiesand Local Physiologic RacesofMagnaporthe grisea in Sichuan Province[A].In:Guoliang Wang and Babara Valent.Advances in Genetics,Genomics and Control of Rice Blast Disease[M].Netherlands:Springer,2008:229-238.
[6]金星,谈笑凤.贵州主要水稻品种对稻瘟病的抗性监测研究[J].耕作与栽培,2009(2):57-59.
[7]王旭伟,张召荣,徐兴慧,等.重庆地区2004年稻瘟病局部成灾致因浅析[J].植物医生,2005,18(2):10.
[8]张雪梅,冯慧,白玉连,等.水稻抗稻瘟病基因和杂交[J].稻品种在四川的抗性评价[J].西南农业学报,2012,25(4): 1266-1272.
[9]刘晋,桑维均,金星.贵州部分水稻主栽品种对稻瘟病的抗性评价[J].种子,2009,28(2):64-67.
[10]Hiroshi T,Marry JT Y,Erbon L A,et al.Development ofmonogenic lines of rice for blast resistance[J].Breeding Science,2000,50(3):229-234.
[11]中华人民共和国农业行业标准.水稻抗稻瘟病鉴定技术规范NY/T[S].
[12]王伟舵,于俊杰,聂亚锋,等.2011-2014年江苏省稻瘟病菌种群动态及毒力变化[J].江苏农业学报,2015,31(2):285-289.
[13]马军韬,张国民,辛爱华,等.水稻品种对黑龙江省部分稻瘟病病菌抗病性分析[J].中国植保导刊,2014,34(11):7-11.
[14]杨璨,王玉翠,吴伟怀,等.稻瘟病菌群体遗传结构及致病型的研究进展[J].安徽农业学报,2015,43(13):111-114.
[15]马军韬,张国民,辛爱华,等.哈尔滨地区抗瘟基因抗性分析及水稻品种抗性评价与利用[J].植物保护学报,2015,42(2): 160-168.
[16]史明乐,刘志恒,陈毓苓,等.苏辽粳稻区稻瘟病菌群体多样性比较[J].植物病理学报,2015,45(2):158-166.
(责任编辑 陈虹)
Difference in Virulence of Magnaporthe oryzae from Sichuan,Chongqing and Guizhou
ZHANG Shu1,ZHONG Xue-lian1,QIAO Gui-yan1,SHEN Li2,ZHOU Tian-yun3,PENG Yun-liang1*
(1.Institute of Plant Protection,Sichuan Academy of Agricultural Sciences/MOA Key Laboratory for Integrated Pest Management on Crops in Southwest China,Sichuan Chengdu 610066,China;2.Plant Protection Station,Sichuan Provincial Department of Agriculture,Sichuan Chengdu 610042,China;3.Chongqing Municipal Station of Seed Management,Chongqing 401121,China)
Sichuan,Chongqing and Guizhou are among the most important areas for rice blast disease(Magnaporthe oryzae)to take place and cause losses.To investigate the difference in the virulence of blast fungus between the different regions,itwill be benefit for prolonging the use of resistance cultivars.In 2012 and 2013,24 monogenic linesof rice for blast resistance of the common background of Lijiangxintuan Heigu(LTH)were planted at the locations of frequent epidemic of blast disease in 21 counties of Sichuan province and Chongqing city.In 2014 and 2015,panicle blast specimenswere collected from totally 47 paddy fields from 15 counties in whole Guizhou provice,10 counties in the northern part of Sichuan Province and 2 counties in Shandong provincesand immersed in water for4 hoursbefore they were inoculated on the arrays of16 varieties and the 24 monogenic lines of rice.After themulching and complete segregation by plastic membrane for 40 days,to investigate the rice blast incidences of16 varieties and the24 monogenic lines,The sesult of experiment in 2012 and 2013 indicate that resistance gene Pi2 was then found to express the resistance in 38.10%of the disease nurseries.The rice blast identification results of panicle blast specimens which collecting form Sichan,Guizhou and other place indicate that resistance genes Pikh,Piks,and PiKm were found to be resistant to blast fungus respectively from 51.85%,48.15%and 48.15%of the disease fields in Guizhou,PiKm,Pi2,Pi9,Pikh,and Pikswere resistant to theblast fungus respectively from 70.00%,60.00%,50.00%,50.00%and 45.00%of the disease fields in Sichuan.The frequency for Pi2 to be resistant to the blast fungus from fields in northern part of Sichuan was significantly higher than that in Guizhou(P=0.0066).The frequencies for 3 differential host varieties i.e.Chenghui448,Neihui99-14 and RHR-1,to be resistant to blast fungus from the fields in Guizhou and northern part of Sichuan were respectively 74.07%,70.37%and 70.37%vs.90.00%,70.00%and 90.00%,which were significantly higher than that for other three differential host varieties,i.e.IR24,Minghui63 and DuohuiNo.1.The frequencies for 10 hybrid rice varieties to express the resistance to blast fungus from the fields in Guizhou and northern partof Sichuan varies respectively between 81.48%-92.59%and 90.00%-100.00%.While the cluster analysis of the virulence of blast fungus in the disease nurseries and paddy fieldshad indicated the bigger difference in virulence roughly in accordance to the geographic distance,the virulence of blast fungus from the different fields in same county and even the same sampling sites could be significantly different.On the other hand,the blast fungus from different counties and different provinces could be virulent to all of the 24 monogenic lines and the further differentiation of the virulence relied on other rice varieties.
Magnaporthe oryzae;Monogenic lines of rice for blast resistance;Resistance frequency;Virulence of isolate;Cluster analysis
S435.111
A
1001-4829(2017)2-0359-07
10.16213/j.cnki.scjas.2017.2.020
2016-03-20
公益性(农业)行业专项(201203014);国家水稻产业技术体系专项经费(CARS-01-25);四川省育种攻关项目(YZGG2016-25)
张姝(1988-),女,四川新都人,硕士,从事水稻病虫综合防控研究,E-mail:becky_zs@126.com,*为通讯作者:彭云良,E-mail:pengyunliang@aliyun.com。
