抗寒锻炼期果园气温对库尔勒香梨不同越冬期抗寒性的影响
2017-04-13艾克来木艾合买提阿不来提买买提巴特尔巴克阿丽腾吐尔孙哈力萨吉旦阿卜杜克日木祖力克艳麻那甫
艾克来木·艾合买提,阿不来提·买买提,巴特尔·巴克*,阿丽腾·吐尔孙哈力,萨吉旦·阿卜杜克日木,祖力克艳·麻那甫
(1.新疆农业大学草业与环境科学学院,新疆乌鲁木齐830052;2.和田地区林管站,新疆和田848000)
抗寒锻炼期果园气温对库尔勒香梨不同越冬期抗寒性的影响
艾克来木·艾合买提1,阿不来提·买买提2,巴特尔·巴克1*,阿丽腾·吐尔孙哈力1,萨吉旦·阿卜杜克日木1,祖力克艳·麻那甫1
(1.新疆农业大学草业与环境科学学院,新疆乌鲁木齐830052;2.和田地区林管站,新疆和田848000)
为了解释抗寒锻炼期果园气温变化对库尔勒香梨不同越冬期抗寒性的影响,连续测定了2年不同越冬期间库尔勒香梨树干阳阴面形成层、一年生枝条和花芽等器官和组织的SOD、CAT、POD酶活性、可溶性蛋白含量、相对电导率、果园气温等生理指标和环境因子;采用隶属函数综合评价2年不同越冬期抗寒性,分析抗寒锻炼期气温与不同越冬期抗寒性变化之间的关系。抗寒锻炼期间气温变化一定程度上影响不同越冬期库尔勒香梨表现出的抗寒性,抗寒锻炼期气温波动性小的年份不同越冬期表现出的抗寒性强于气温波动性大的年份;不同生理指标与气温均呈负相关,其中气温与保护酶呈极显著性负相关,生理指标中可溶性蛋白和POD对库尔勒香梨抗寒性的贡献率比其它生理指标大;总体上库尔勒香梨属于抗寒性品种。
库尔勒香梨;抗寒锻炼;果园气温;隶属函数;抗寒性;生理指标
低温是影响库尔勒香梨(Pyrus brestschneideri Rehd.)生长和产量关键的环境因素之一,每年都造成一定程度的冻害,导致很大的经济损失。2000年以来库尔勒香梨经历过2次较大范围的严重冻害。2007/2008年冬季库尔勒市最低温度达到-23.9℃,远离市区的乡村则达到-26.3℃,果园气温会低于-26.3℃,遭受面积达1×104hm2,减产5×104t,直接经济损失3亿元人民币[1]。2010/2011年也有很严重的冻害,经初步调查受害面积大约1.3× 104hm2(巴州香梨研究中心)。本研究果园平均气温比市区平均气温低于3~5℃。
在温带和寒带多年生植物在冬季,通过将活跃的生长器官和组织进入休眠状态来增强抵御和适应寒冷胁迫[2]。Janská等发现紫花苜蓿的抗寒过程包含秋季低温驯化与越冬耐寒适应两个方面[3],大量研究表明,与未经历低温驯化相比,经历低温驯化的植物抗寒性大幅提高[4]。
低温驯化或抗寒锻炼是多年生植物在秋末冬初感受逐渐下降的外界温度,并且受光周期的诱导,经由细胞信号转导、基因表达调控等一系列过程,引发植物体内发生一系列生理生化变化,从而提高对冬季低温的抵抗能力的过程[5]。前人关于甜樱桃[6],蓝莓[7],樟子松[8],紫苜蓿[9],阿月浑子[10]、小麦[11]等树和植物的研究发现抗寒锻炼与植物抗寒性密切相关,植物经过低温驯化后,增加了细胞膜不饱和脂肪酸的含量,从而提高了膜的抗冻性,同时低温驯化提高植物的抗氧化能力,提高细胞内可溶性物质等的含量,增强抗冻能力[12]。
本研究将库尔勒香梨的越冬期分为5个月份,分别为11、12、1、2和3月份,其中11月份作为抗寒锻炼期。
气温是影响植物抗寒锻炼或低温驯化主要的环境因子之一,前人对人工和自然低温胁破条件下,低温驯化与抗寒性之间关系的研究较多,但关于抗寒锻炼期间气温变化对香梨抗寒性影响的报道鲜见,本文比较两年抗寒锻炼期间气温变化状况,分析它们对库尔勒香梨不同越冬期抗寒性的影响,解释香梨对低温的适应和抵抗机制,为减少冻害造成的损失提供参考。
1 材料与方法
1.1 实验材料
实验从2013年11月至2014年4月、2014年11月至2015年4月在库尔勒市阿瓦提乡梨园(41.69°N,86.10°E)进行,实验材料为23年生,健康,长势相似的库尔勒香梨树阳阴面树干形成层(离地面60 cm),一年生枝条和花芽,每个样品有5个重复。
1.2 研究方法
1.2.1 野外取样和预处理香梨阳阴面树干形成层,一年生枝条和花芽等器官进行采样,采样日期别为,2013年12月4日、2014年1月5日、2014年2月3日、2014年3月5日和2014年12月2日、2014年12月31日、2015年1月20日、2015年2月11日和2015年3月16日。
采样后立即用纱布和锡箔纸包裹放入液氮罐送回实验室转入-80℃的低温冰箱冷藏,取出样品时先把样品放在-40℃的低温冰箱,自然升温到0℃以后,按照要求在实验室分别测定阳阴面形成层,枝条形成层和花芽(去除鳞片)等器官和组织的超氧化物歧化酶(SOD)、过氧化氢酶(CAT)、过氧化物酶(POD)、相对电导率、可溶性蛋白含量等与抗寒性相关的主要生理指标,测定每个指标有5个重复。
1.2.2 气象因子的监测与处理实验进行期间两年连续监测测定越冬期果园2 m高处的空气温度(型号:AV-ST),每15 min记录一次数据,监测数据采用美国Avelon公司生产的AR5型数据采集器存储,根据所监测的15 min的气象因子分别计算每小时、日值、月值等。
1.2.3 生理参数的测定POD活性的测定:愈创木酚法[13]。CAT活性的测定:紫外吸收法[14]。SOD活性的测定:氮蓝四唑法[14]。可溶性蛋白含量:采用考马斯亮蓝G-250染色法[14]。相对电导率测定采用电导仪法[15]。
1.2.4 抗寒性综合评价方法应用Fuzzy数学中隶属函数法[16-17]进行综合评判,算式为:

式中:U(Xijk)为第i个器官第j个温度阶段第k项指标的隶属度,且U(Xijk)∈[0,1];Xijk表示第i个器官第j个温度阶段第k个指标测定值;Xmax、Xmin为所有参试种中第k项指标的最大值和最小值。
1.2.5 数据处理与分析采用SPSS20、Excel 2013等软件进行数据分析,采用Pearson相关系数来分析相关性。
2 结果与分析
2.1 越冬期间果园气温的变化
2013/2014年越冬期间最低平均气温为-14.00℃(表1,出现在1月份,极端最低气温为-28.30℃,出现日期为12月30日,极端低温出现日-20℃的气温持续时间为18 h,-26℃极端低温持续时间是6 h;1月7日又一次出现极端低温,-20℃的气温持续时间为15 h,-26℃极端低温持续时间为6 h,远远超过前人报道中的对香梨造成冻害的阈值。最低气温出现时间主要集中在0:00-10:00 (乌鲁木齐时间),12月30日极端低温出现时间为2:00。整个越冬期连续出现-20℃低温日数为26 d,其中-25℃低温连续持续时间6 h上的日数为9 d。
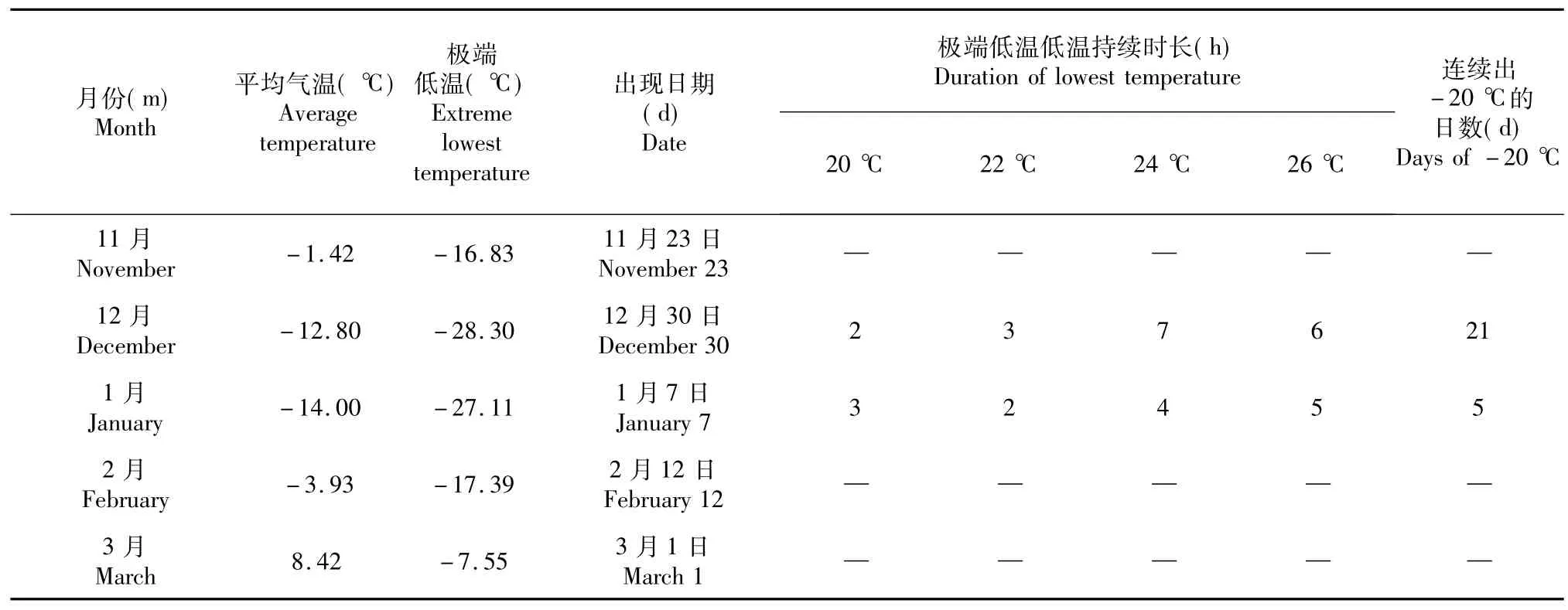
表1 2013/2014年越冬期间果园气温的变化Table 1 Changes of orchard air temperature during over-wintering periods in 2013/2014
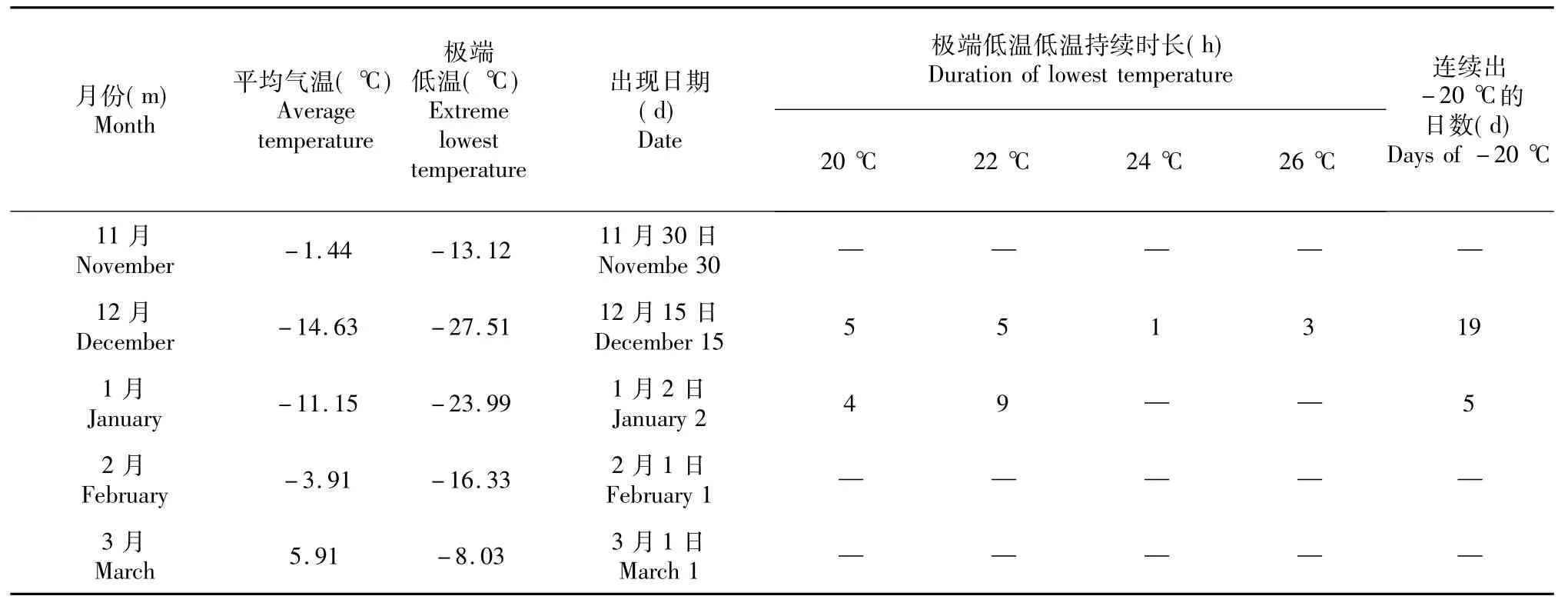
表2 2014/2015年越冬期间果园气温的变化Table 2 Changes of orchard temperature during over-wintering periods in 2014/2015
2014/2015年越冬期最低平均气温为-14.63 (表2),出现在12月份,极端最低气温为-27.51℃,出现日期为12月15日,极端低温出现日-20℃的气温持续时间为14 h,-26℃极端低温持续时间为3 h,极端低温出现日期比2013/2014年提前15 d。最低气温出现时间主要集中在3:00-10:00 (乌鲁木齐时间),12月15日极端低温出现时间为9:00。整个越冬期连续出现-20℃低温日数为21 d,其中-25℃低温连续持续时间6 h上的日数为3 d,出现日期分别为12月15日、12月16日和12月17日。
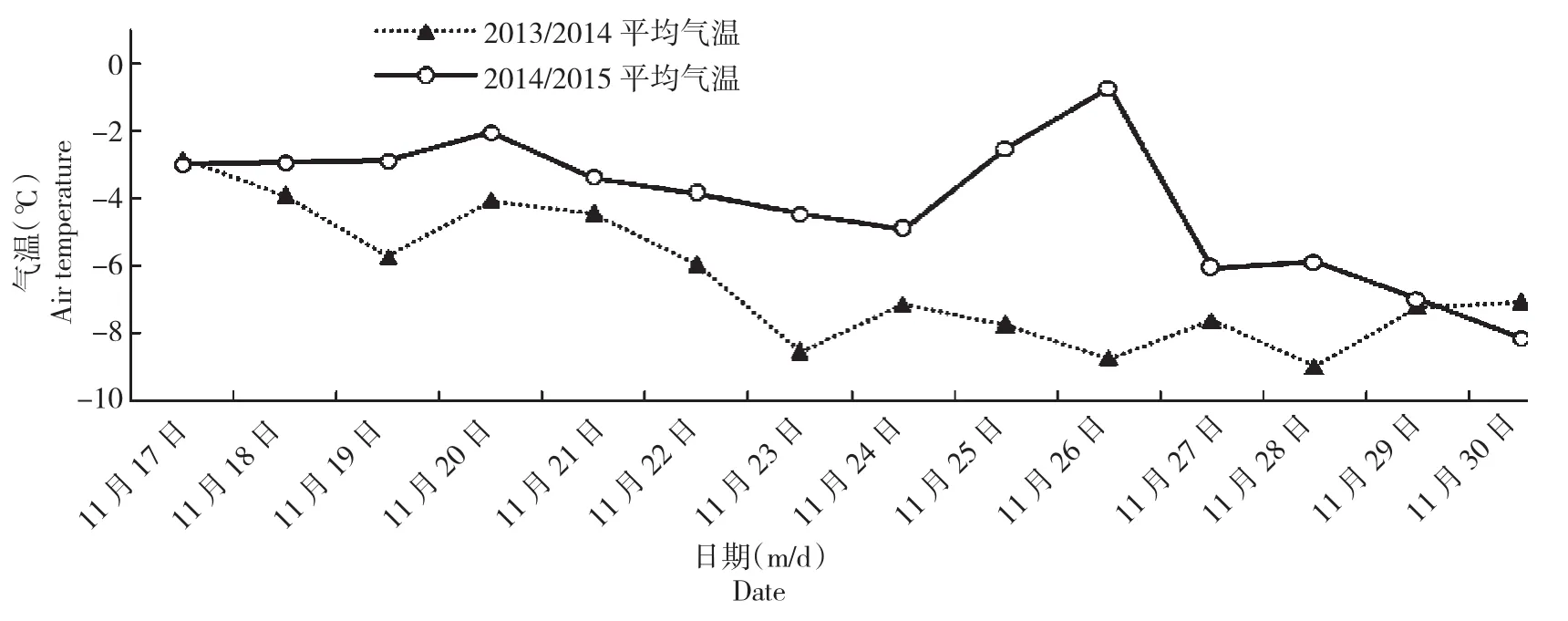
图1 抗寒锻炼期平均气温的变化Fig.1 Changes of average air temperature during cold hardening periods
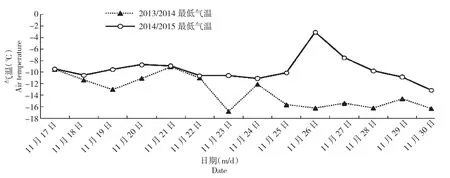
图2 抗寒锻炼期最低气温的变化Fig.2 Changes of the lowest air temperature during cold hardening periods
根据以上2年越冬期间平均气温、极端低温以及低温持续时间等气象因子的变化可以断定2013/ 2014年香梨受到的低温逆境强度比2014/2015年大。
2.2 抗寒锻炼期果园气温的变化
图1~2可以看出,2014/2015年抗寒锻炼期平均气温和极端低温变化的波动性较大,平均气温比2013/2014年高于3℃左右,11月26日出现-0.69℃的反常气温,比11月27日高于6℃,短时间内这种大幅度的气温变化会加强融冻交替,可能增加细胞膜机械损伤程度,影响细胞膜的正常代谢功能; 2014/2015年抗寒锻炼期没出现短时间内气温急剧上升或者急剧降低的天气。
2.3 气温和生理指标之间的相关性分析
气温与测定的生理指标均呈负相关(表3),其中与SOD、CAT、POD等保护酶之间的相关性达到极显著水平;不同生理指标之间SOD与CAT、POD分别呈极显著性正相关,SOD与可溶性蛋白含量呈显著性正相关;CAT与POD、可溶性蛋白含量呈显著性正相关;POD与可溶性蛋白含量呈极显著性正相关;保护酶和可溶性蛋白与相对电导率均呈负相关,但相关性没达到显著水平。
2.4 隶属函数综合评价2年不同越冬期间抗寒性
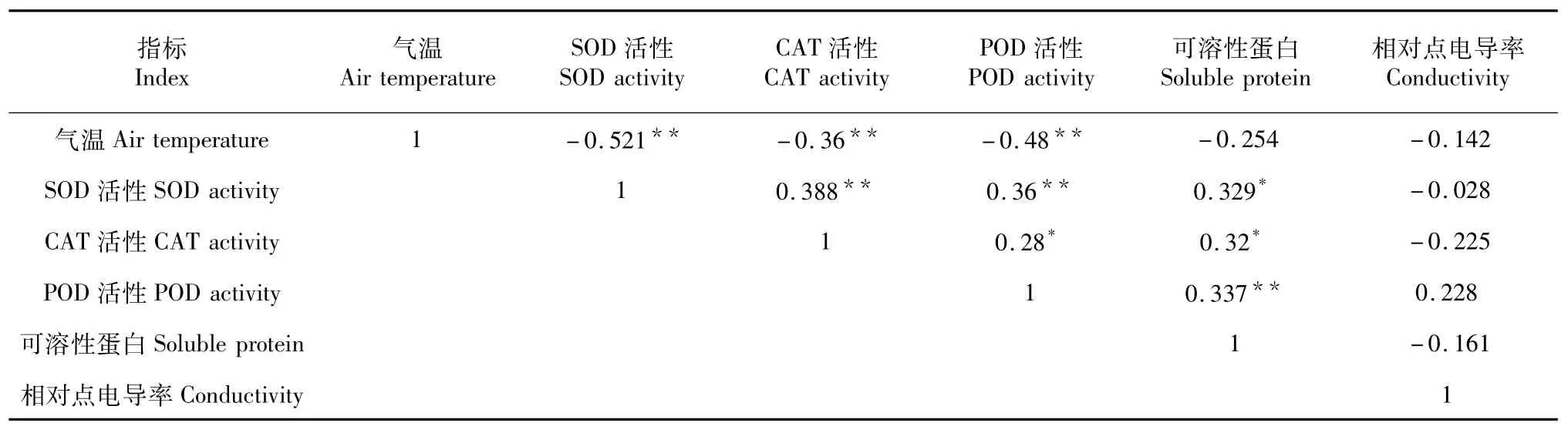
表3 2014/2015年越冬期间气温与不同生理指标相关性Table 3 Correlation between different physiological index and air temperature during over-wintering periods in 2014/2015
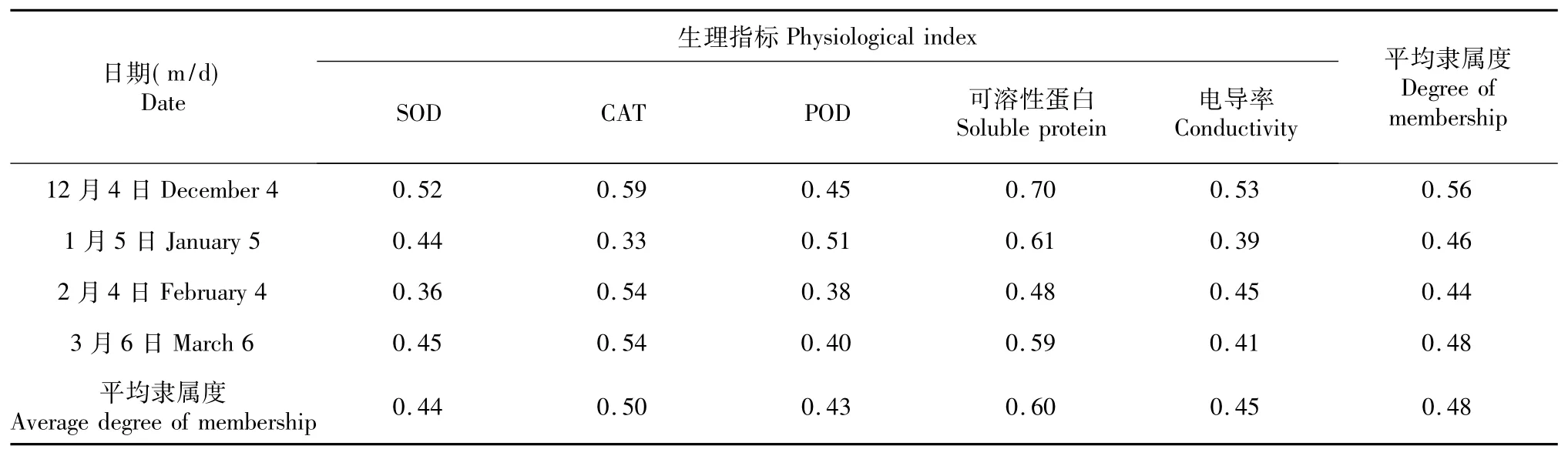
表4 2013/2014年不同越冬期香梨抗寒性的变化Table 4 Changes of cold resistance of fragrant pear during different over-wintering periods in 2013/2014
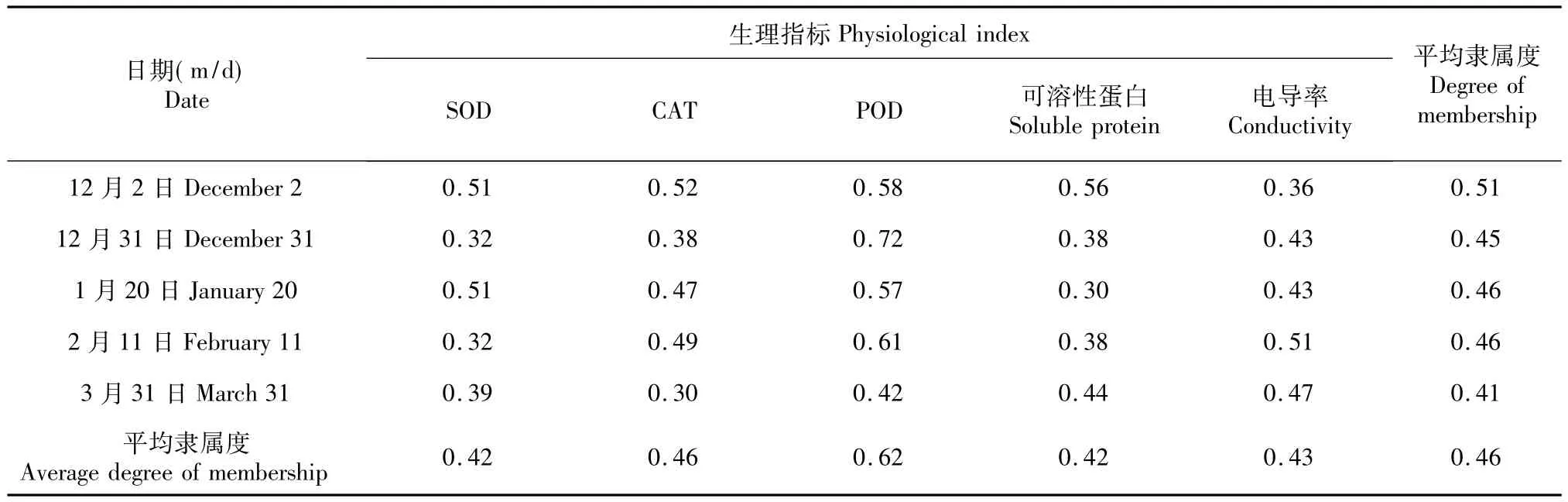
表5 2014/2015年不同越冬期香梨抗寒性的变化Table 5 Changes of cold resistance of fragrant pear during different over-wintering periods in 2014/2015
前人运用Fuzzy数学隶属函数综合评判法,对核桃[18-19]、扁桃[20]、草莓[21]等果树和植物的抗寒性进行综合评价,取得了与实际符合的结果。采用隶属函数法评价抗寒性时,隶属函数值越大抗寒性越强,反之抗性越弱,根据平均隶属度库尔勒香梨属于抗寒性品种。
从表4可以看出,2013/2104年在整个越冬期库尔勒香梨在12月份表现出最强的抗寒性;2月份抗寒性最弱,这可能是2月份虽然气温比12月份和1月份高,但气温日较差值比12月份和1月份大,引起的融冻交替程度大,会进一步加大细胞膜受到的机械损伤程度,从而导致大量渗透调节物质外渗,细胞失去了结构和功能的整体性从而减弱抗寒性的原因。本实验整个越冬期不同器官在2月份相对点电导率是最大,可说明2月份细胞膜所受的伤害程度最大以及融冻交替引起的机械损伤是伤害细胞膜和影响抗寒性的主要原因之一。越冬期不同阶段各生理指标中隶属值最大的是可溶性蛋白,对机体增强抗寒性的贡献率最大,可说明可溶性蛋白是增大细胞渗透势,有利于提高机体抗寒性的渗透调节物质。
2014/2015年不同越冬期香梨抗寒性变化与2013/2014稍微不同(表5),12月份抗寒性最强,3月份最低,这可能是3月份温度回升的原因;虽然2月份抗寒性较强,但还是比12月份抗寒性弱。本实验不同越冬期2月份相对电导率最大,而POD隶属值也最高,这可能是细胞受到较大损伤时POD表现出较高的酶活性,对机体起到保护作用的原因。
比较2年库尔勒香梨所表现出的抗寒性,按整个越冬期来说,2013/2014年抗寒性强于2014/2015年的抗寒性;按不同越冬期来比较,2013/2014极端低温出现后香梨表现出的抗寒性稍微高于2014/ 2015年极端低温出现后香梨表现出的抗寒性。本实验抗寒锻炼期气温变化对不同越冬期抗寒性有明显的影响。
3 讨论
抗寒性是库尔勒香梨对低温环境长期适应而形成的一种遗传特性。香梨感受到低温之后,体内发生一系列酶促反应和酶合成反应等生理生化变化,为安全越冬做抗寒准备,因此,在抗寒锻炼期或者低温驯化期间气温变化状况直接影响进入抗寒锻炼的质量。
2010/2011年在库尔勒地区出现暖冬,引起了抗寒锻炼延迟并提早解除抗寒锻炼的现象,除此之外,在抗寒锻炼期间气温变化幅度大,严重影响抗寒锻炼的正常进行,导致抗寒性降低,在极端低温出现后造成面积大约1.3×104hm2梨园冻死(巴州香梨研究中心),这与1996年Repo等研究具有相似的结果[22]。
Bélanger等(2002)发现,冬季在气候变化,全球变暖背景下的气象因子的变化增加了多年生作物受到的低温伤害程度[23]。暖冬的出现絮乱了香梨抗寒锻炼生理生化反应和越冬准备的正常进行及极端低温出现时间和持续时间,增大短时间内气温突然变化的概率,适应和抵制低温伤害是香梨在几百万年进化的结果,因此很难适应短时间内突然的天气变化。Crawford等(2000)发现,突然天气变化的情况下,植物遭受的危害更不稳定,例如:冷冻-融化都可能导致植物死亡[25]。
本实验2014/2015年,抗寒锻炼气温变化波动性大,抗寒锻炼期出现较高的气温,这些现象是暖冬出现的表现,它不仅通过直接影响抗寒锻炼质量来影响香梨抗寒性,还可以通过影响细胞糖代谢影响机体抗寒性,这可能是2014/2015年越冬期低温伤害强度比2013/2014年低,表现出的抗寒性比2013/2014年弱的原因。前人研究表明,在冬季较温暖的气温条件,促进植物呼吸,从此增加糖类的消耗,导致细胞渗透调节物质含量降低,细胞亲和力下降,大量渗透调节物质外渗,树木抗寒性的下降[24]。
植物经历低温驯化或者抗寒锻炼后,抗寒性明显增加,本实验2013/2014、2014/2015年整个越冬期间,12月份表现出最强的抗寒性,与前人研究结果类似。根据本实验结果可以总结,库尔勒香梨进入抗寒锻炼期间的气温变化状况是直接影响香梨抗寒性的重要的环境因素。
本实验越冬期间,气温变化与SOD、CAT、POD、可溶性氮含量和相对电导率等生理指标均呈负相关,气温与抗氧化酶呈极显著正相关;POD和可溶性蛋白含量对抗寒性的贡献率比其它生理指标大。
4 结论
越冬期库尔勒香梨抗寒性最强的时期是12月份;抗寒锻炼期间气温变化幅度影响不同越冬期库尔勒香梨表现出的抗寒性。与香梨抗寒性相关的生理指标与气温呈负相关,生理指标中可溶性蛋白和POD对库尔勒香梨抗寒性的贡献率比其它生理指标大。库尔勒香梨属于抗寒性品种。
[1]巴特尔·巴克,克热木·伊力,匡玉疆,艾合买提·阿不都热依木.库尔勒香梨历年冬季低温评价及严重冻害成因分析[J].新疆农业大学学报,2008,31(6):17-20.
[2]Wisniewski M,Norelli J,Bassett C,et al.Ectopic expression of a novel peach(Prunus persica)cbf transcription factor in apple(Malus ×domestica)results in short-day induced dormancy and increased cold hardiness[J].Planta,2011,233(5):971-983.
[3]JanskáA,Marík P,ZelenkováS,et al.Cold stress and acclimation-What is important for metabolic adjustment[J].Plant Biology,2010,12(3):395-405.
[4]Castonguay Y,Bertrand A,Michaud R,etal.Cold-induced biochemical and molecular changes in alfalfa populations selectively improved for freezing tolerance[J].Crop Science,2011,51(5):2132-2144.
[5]Ensminger I,Busch F,Huner N P A.Photo stasis and cold acclimation:Sensing low temperature through photosynthesis[J].Physiologia Plantarum,2006,126(1):28-44.
[6]Melbar.Salazar-Gutiérrez,Bernardo Chaves.Variation in cold hardiness of sweet cherry flower buds through different phenological stages[J].Scientia Horticulturae,2014,172(3):161-167.
[7]Takashi Kikuchi,Kiyoshi Masuda.Class II chitinase accumulated in the bark tissue involveswith the cold hardiness of shoot stems in high bush blueberry[J].Scientia Horticulturae,2009,120(2):230-236.
[8]李臻,孙彪彪,张钢,等.抗寒锻炼期间樟子松抗寒性与生理指标的变化[J].河北农业大学学报,2013(1):46-49.
[9]刘志英,李西良,李峰,等.紫花苜蓿秋眠性对低温驯化过程与越冬耐寒适应的作用机理[J].植物生态学报,2015(6):635-648.
[10]Pakkish Zahra,RahemiMajid,Baghizadeh Amin.Seasonal Changes of Peroxidase,Polyphenol Oxidase Enzyme Activity and Phenol Content During and after Rest in Pistachio(Pista ciavera L.)Flower Buds[J].World Applied Sciences Journal,2009,6(9):1193-1199.
[11]王晓楠,付连双,李卓夫,等.低温驯化及封冻阶段不同冬小麦品种叶绿素荧光参数的比较[J].麦类作物学报,2009(1):83-88.
[12]Castonguay Y,DubéM P,Cloutier J,etal.Molecular physiology and breeding at the crossroads of cold hardiness improvement[J].Physiologia Plantarum,2013,147(1):64-74.
[13]郝再彬,苍晶,徐仲.植物生理实验技术[M].哈尔滨:哈尔滨出版社,2002:204-205.
[14]李合生.植物生理生化试验原理与技术[M].北京:高等教育出版社,2000:165-169.
[15]冯献宾,董倩,李旭新,等.黄连木和黄山栾树的抗寒性[J].应用生态学报,2011,22(5):1141-1146.
[16]LIYi-Bing,Yang SQ,Ren G X,et al.Changes analysis in physiological properties of several gramineous grass species and cold-resistance comparison on under cold stress[J].Acta Ecologica Sinica,2009,29(3):1341-1347.
[17]Pan X Y,Cao Q D,Wang G X.Evaluation of lipid peroxidation for use inselection of cold hardiness cultivarsofalmond[J].Acta Ecologica Sinica,2002,22(11):1902-1911.
[18]相昆,张美勇,徐颖,等.不同核桃品种耐寒特性综合评价[J].应用生态学报,2011,22(9):2325-2330.
[19]刘杜玲,张博勇,孙红梅,等.早实核桃不同品种抗寒性综合评价[J].园艺学报,2015,34(3):545-553.
[20]李斌,刘立强,罗淑萍,等.扁桃花芽的抗寒性测定与综合评价[J].经济林研究,2012,30(3):16-21.
[21]杨凤翔,金芳,颜霞.不同草莓品种抗寒性综合评价[J].果树学报,2010(3):368-372.
[22]Repo T,Hanninnen H,Kellomaki S.The effects of long-term elevation of air temperature and CO2on the frost hardiness of Scots Pine[J].Plant Cell and Environment,1996,19(2):209-216.
[23]Bélanger G,Rochette P,Castonguay Y,et al.Climate change and winter survival of perennial forage crops in eastern Canada[J].Agronomy Journal,2007,61(5):963-975.
[24]Crawford R M M.Ecological hazards of oceanic environment[J].New Phytologist,2000,147(2):257-281.
[25]Gren E,Nilsson T,Sundblad L G.Relationship between respiratory depletion of sugars and loss of cold hardiness in coniferous of different sensitivities of spruce and pine[J].Plant Cell and Environment,1997,20(2):247-253.
(责任编辑 陈虹)
Effect of Orchard Tem perature on Cold Resistance of Korla Fragrant Pear during Cold Hardening Period
Akrem Ahmet1,Ablet Mamat2,Batur Bake1*,Altun Tursunhali1,Sajida Abdukarim1,Zulkiye Manaf1
(1.College of Grassland and Environmental Science,Xinjiang Agricultural University,Xinjiang Urumqi830052,China;2.Forest Management Station,Hotan Prefecture,Xinjiang Hotan 848000,China)
To explain the effect of temperature changes during cold resistance period on the cold hardiness of Korla fragrant pear’s different over-wintering period,the SOD,CAT,POD activity,soluble protein content,relative conductivity of Korla Fragrant Pear’s trunk cambium,annual branch and bud for two yearswere continuously determined,and the air temperature of the orchard were simultaneouslymeasured;Its cold resistance of different over-wintering period by using fuzzymathematicsmethod was evaluated,and the relation between air temperature changes and its cold resistance over thewinterwere analyzed.The result showed that the air temperature changes of the cold hardening period affected the cold resistance of Korla fragrant pear during differentwintering periods,the yearwhich had the smaller temperature fluctuation range during the cold hardening period expressed stronger cold resistance than the yearwhich had the bigger temperature fluctuation range; Different physiological indexeswere negatively correlated with air temperature changes,ofwhich air temperaturewas very negatively correlated with anti-oxidantenzymes,and the POD and soluble protein contenthad the biggest contribution rate for the cold resistance;Overall Korla fragrant pear belonged to cold resistance varieties.
Korla fragrant pear;Cold hardening;Orchard temperature;Fuzzymathematicsmethod;Cold resistance;Physiological index
S661.2
A
1001-4829(2017)2-0309-06
10.16213/j.cnki.scjas.2017.2.012
2016-03-10
国家自然科学基金(31260289);新疆维吾尔自治区土壤学重点学科
艾克来木·艾合买提(1990-),男,维吾尔族,新疆图木舒克人,硕士研究生,研究方向为干旱区资源与环境,E-mail:akeram27@163.com,*为通讯作者:巴特尔·巴克,教授,研究方向干旱区生态与环境,E-mail:bateerbake@163.com。
