新麦草种质资源表型多样性的主成分和聚类分析
2017-04-12郝林峰李志勇李鸿雁
郝林峰,李志勇,李 俊,黄 帆,李鸿雁,﹡
(1.包头市家畜改良工作站,内蒙古包头 014010;2.中国农业科学院草原研究所,内蒙古呼和浩特 010010)
新麦草种质资源表型多样性的主成分和聚类分析
郝林峰1,李志勇2,李 俊2,黄 帆2,李鸿雁2,﹡
(1.包头市家畜改良工作站,内蒙古包头 014010;2.中国农业科学院草原研究所,内蒙古呼和浩特 010010)
对来自新疆、陕西、美国、哈萨克、利比亚玻利维亚和吉尔吉斯的11份新麦草野生材料的茎干节数、叶舌长度、叶片长度、叶片宽度、小穗数、小穗长、小花数、颖长度、颖果长度和株高等10个表型性状采用方差分析、主成分分析、聚类分析等方法进行了遗传多样性分析,以期筛选出优良种质,为新麦草育种提供可靠的依据。结果表明:1)新麦草种质间表型性状变异程度较高,变异系数范围为14.6%-68.0%,其中小花数最大,叶片宽度最小;2)10个表型性状可归为3个主成分因子,累计贡献率达到83.34%,最大程度上反映了11份新麦草的形态特征,其中小穗长、小花数、颖长度和植株高度是影响新麦草表型性状变异的主要性状;3)10个表型性状间存在显著或极显著的正相关和负相关;4)采用欧氏距离系统聚类法将11份新麦草材料分为3大类。本研究主要揭示了新麦草的形态遗传多样性,可为新麦草资源的收集、鉴定、评价、保存和资源创新利用奠定理论基础。
新麦草;表型性状;聚类分析
新麦草(Psathyrostachysjuncea(Fisch)Nevski)是禾本科小麦族新麦草属的重要物种。它是一种多年生疏丛型牧草、异花授粉的二倍体(2n=2x=14)禾草〔1〕,原产于中亚和西伯利亚,在我国的天山、阿尔泰山和青藏高原等地有分布〔2〕。新麦草分蘖多、叶量大、抗旱、耐牧、耐盐碱,是一种优良的放牧刈割兼用型牧草,对于我国北部干旱地区的退耕和生态重建有重要的价值,为草地畜牧业和草地生态建设中发挥了重要作用〔2-3〕。我国新麦草属现4个种,分别为华山新麦草、新麦草、毛穗新麦草和单花新麦草,育成品种有山丹新麦草和紫泥泉新麦草〔3〕。种质资源是遗传育种研究的基础,了解种质资源遗传变异等背景,能够加快育种的进程〔4〕。目前关于新麦草野生种质资源遗传变异方面的研究还较少,已报道的野生种质和栽培品种遗传分析中供试的种质较少,代表性不强〔5-9〕。近年来,新麦草的研究集中在合理施肥提高草产量及肥料配比对新麦草人工草地产草量动态关系、放牧刈割对新麦草产量影响、遗传学研究、组织培养技术、杂交育种及转基因育种方面的研究〔10-17〕。而表型性状遗传多样性的研究少见报道,本研究对野生新麦草的主要表型性状进行方差分析、聚类分析、主成分分析和相关分析。本研究的结果能够为新麦草资源的收集、鉴定、评价、保存和资源创新利用奠定理论基础。
1 材料与方法
1.1 试验材料
供试材料由中国农业科学院草原研究所国家种质牧草中期库提供(见表1),试验于2014年在呼和浩特市土默特左旗沙尔沁乡国家多年生牧草圃中进行,该圃为半干旱大陆性气候,年平均降水量400mm左右,无霜期大约为130d左右,≥10℃活动积温在2700℃以上,海拔1065m。土地有机质平均含量为1.5%,土壤类型主要为灰褐土和草甸土,土壤盐碱化程度比较高,适宜抗旱性、抗寒性和耐盐碱性的牧草适宜生长。
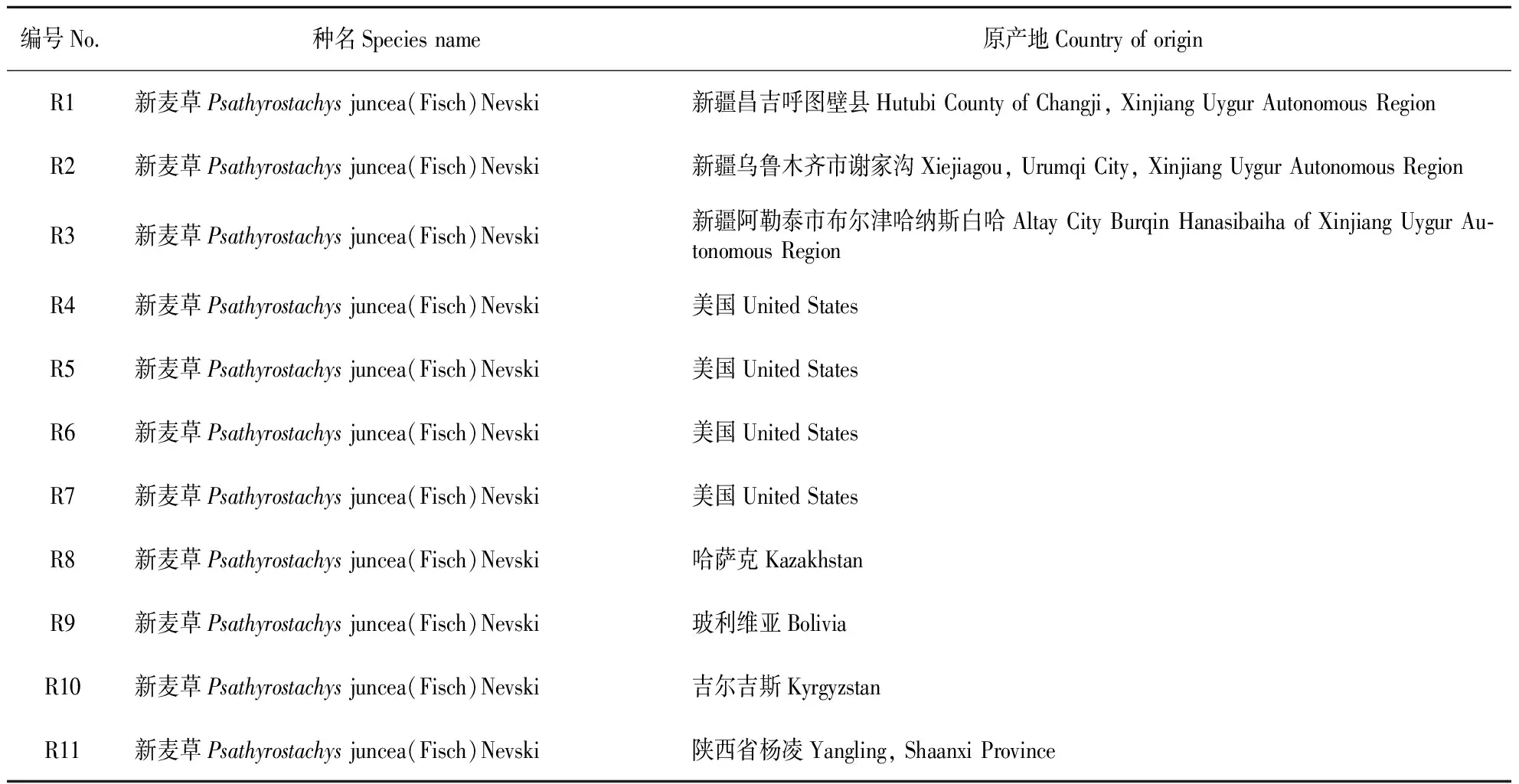
表1 供试材料及原产地
1.2 试验地概况与试验设计
2014年4月份在温室播种育苗,6月份移苗定植在资源圃中,试验地施底肥1次,生育期浇水4-5次,中耕除草3-4次。试验按随机区组设计,3次重复,小区面积20m2,每小区之间相隔60cm。
1.3 性状调查的项目与方法
在鉴定新麦草材料表型性状的同时,也对其物候期等进行了调查记载(见表2)。开花期在田间对每份材料随机取10个单株进行编号挂牌,对这些筛选单株的茎干节数、叶舌长度、叶片长度、叶片宽度、小穗数、小穗长、小花数、颖长度、颖果长度和株高等性状进行观察记载,3次重复,采集2年的数据进行统计分析。
1.4 数据处理
观测记载的数据在Excle软件中进行整理,计算各性状的最大值、最小值、平均值、标准差和变异系数采用SPSS 11.5软件对11份新麦草种质材料的10个表型性状进行聚类分析、主成分分析和相关分析。
2 结果与分析
2.1 不同材料表型性状方差分析
11份新麦草种质材料的10个表型性状进行方差分析(见表2),结果表明,材料间存在较大差异,材料间各性状也存在较大差异。小花数的变异系数最大,为68.0%,变异幅度2.33~10.90个;其次小穗数的变异系数为51.0%,变异幅度15.60~91.00个;叶舌长度的变异系数为46.5%,变异幅度为0.43~1.80mm;叶片长度的变异系数31.3%,变异幅度为15.90~36.70cm。材料间每个性状中都存在着个体间的差异,且不同材料、不同性状之间的差异的程度也不相同〔18〕。采用SPSS 11.5软件进行单因素方差分析,F检验表明(见表2),10个表型性状在材料间的茎干节数、叶片宽度、小穗长、小花数和颖果长度达到显著水平(P<0.05),其余表现差异不显著。茎干节数在10个性状指标中的F值最大,为2.818,10个性状指标的差异度主要有茎干节数、叶片宽度、小穗长、小花数和颖果长度等。
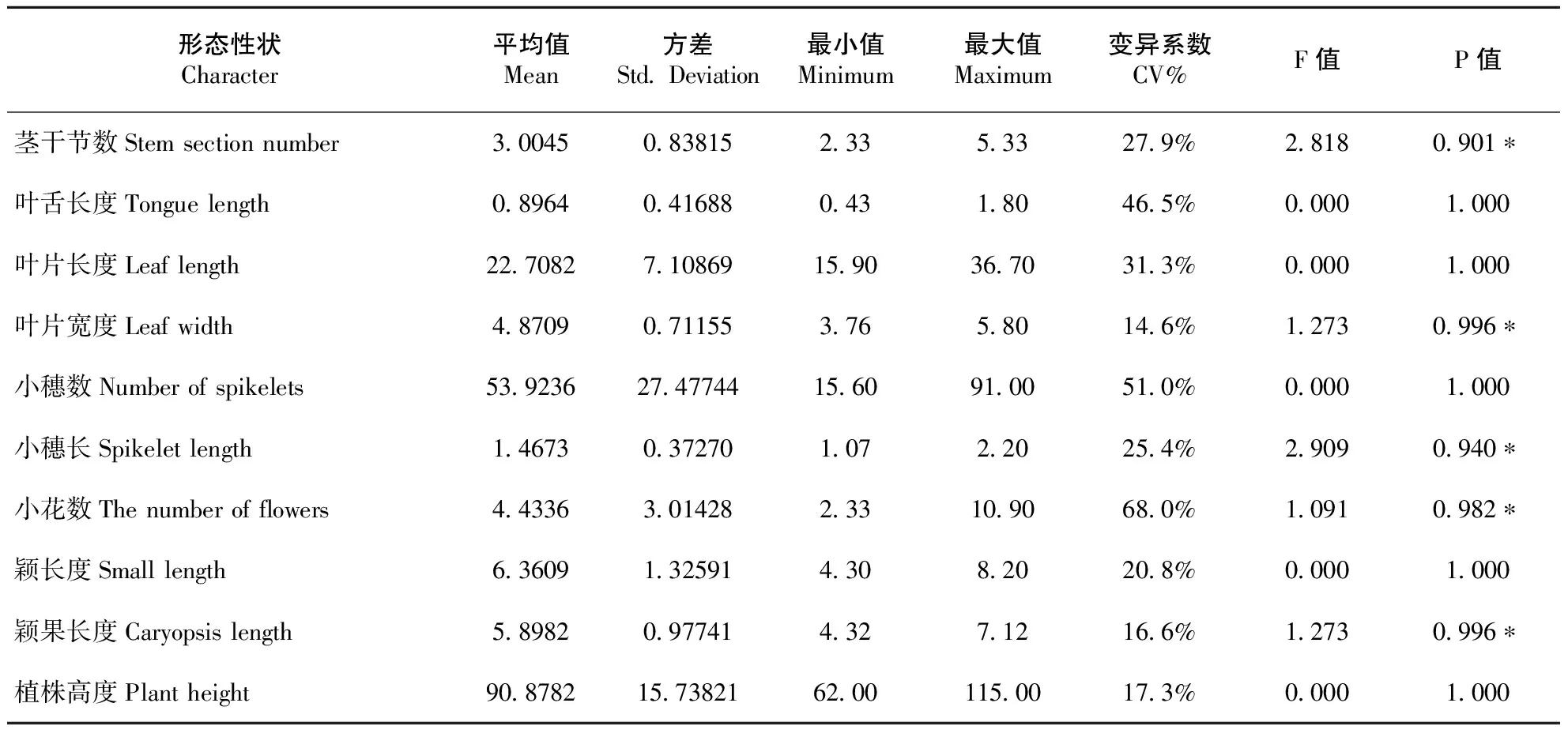
表2 新麦草各性状的平均值、标准差和F值
2.2 不同种质材料各性状的主成分分析
不同新麦草种质资源的10个表型性状主成分分析表明:主成分构成的主要信息集中在前3个主成分,其累计贡献率达83.340%,特征值总和为8.334,反映了原始总体数据的大部分信息。由表3可知,第一主成分特征值为4.630,贡献率46.304%,其特征向量中符号为正,载荷值较高的性状有小穗长、小花数、颖长度和植株高度,其特征向量值分别为0.922、0.837、0.905和0.693,此类性状中均为生殖器官构成因素,因第一主成分的贡献率最高,在选择品种选育中重点考虑这4个性状;第二主成分特征值2.752,贡献率27.519%,特征向量中符号为正且载荷较高的为叶舌长度、叶片宽度此类型主要反映营养器官构成因素。第三主成分特征值0.952,贡献率9.517%,特征向量中符号为正且载荷较高的主要有茎干节数、叶片长度、小穗数和颖果长度,第三主成分主要反映营养和生殖器官因素构成,在一定范围内株高、小花数目等性状的增加对于高产具有重要意义,因此在育种中应适当把握;在前3个主成分包括的10个表型性状中的小穗长、小花数、颖长度和植株高度等性状是造成新麦草种质资源表型差异的主要因素,也是新麦草种质育种选择过程中主要考虑的形态指标。
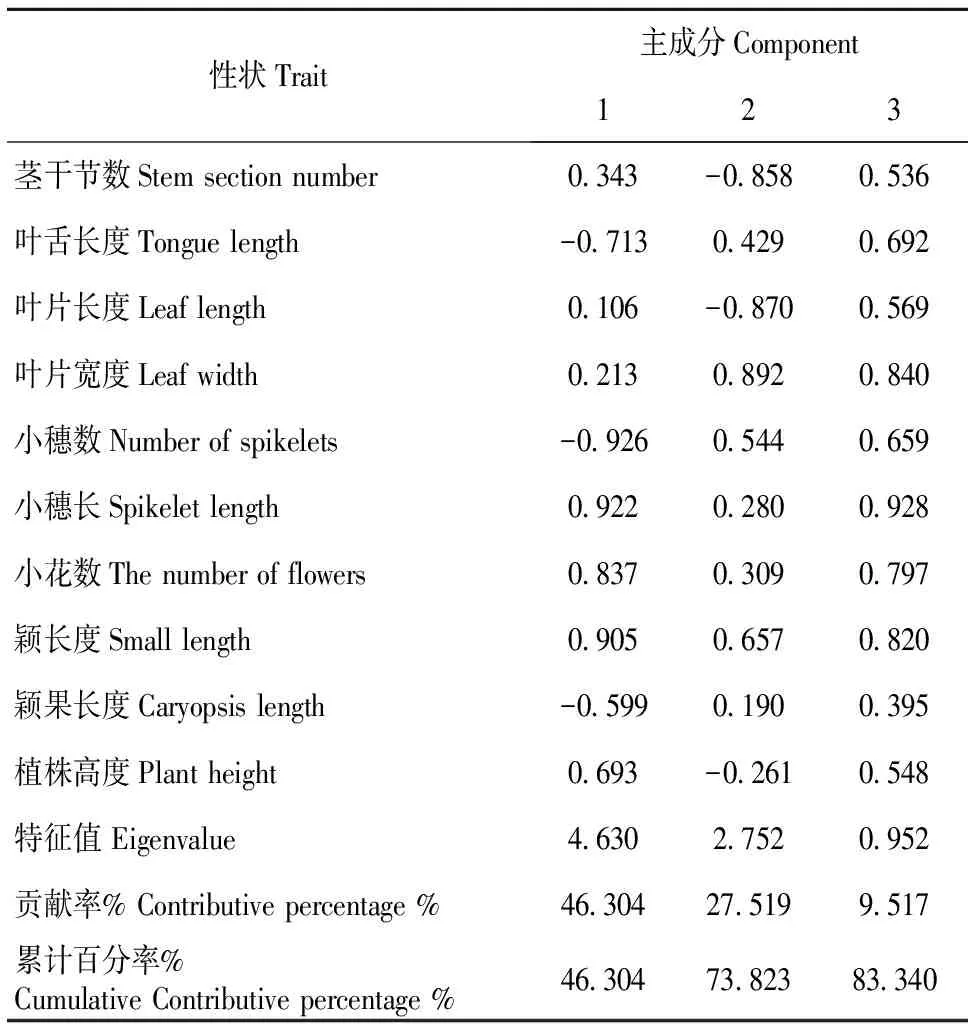
表3 新麦草主要表型性状的主成分分析
2.3 不同居群各性状的聚类分析
采用SPSS 11.5软件对供试的11份新麦草种质依据10个表型性状系统聚类来进行分析(见图1),结果表明在欧氏距离为15时,可将其大致分为3类。第Ⅰ类包括4份材料,分别来自于哈萨克的R8、玻利维亚的R9、吉尔吉斯的R10和陕西杨凌的R11;第Ⅱ类包括5份材料,分别来自于新疆昌吉呼图壁县的R1、美国的R4、R5、R6、R7;第Ⅲ类包括2份材料,均来自于新疆;11份新麦草种质资源基本上按地理类别区别开来,个别的有交叉。11份新麦草种间和材料间表型差异很明显,也反应了新麦草种质资源较丰富多样。生长在同一地区的新麦草种质资源生境大致相同,其性状表现也较相近,说明本试验的系统聚类,较好地反映了种质材料和种间的遗传差异。
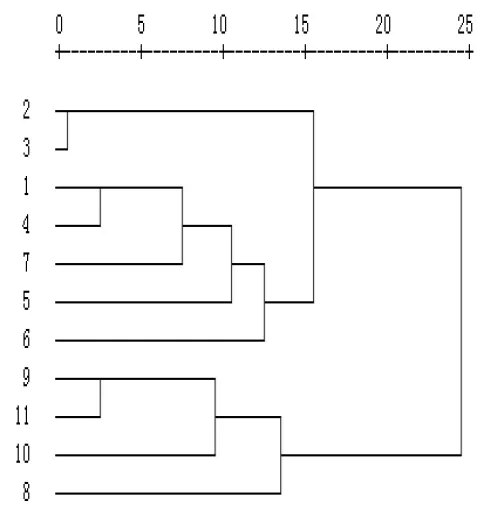
图1 基于表型性状的葱属资源聚类图
2.4 不同居群各性状的相关分析
对11份新麦草种质材料的10个表型性状间的相关关系进行分析(见表4),结果表明10个性状不同程度地与其他性状呈显著或极显著相关性。其中茎干节数与叶片长度呈显著正相关,与叶片宽度呈显著负相关,相关系数分别为0.585和-0.519;叶片宽度与小花数呈显著正相关,相关系数为0.481;小穗数分别与小穗长、植株高度呈显著负相关,相关系数分别为-0.598和-0.527,而与颖长度呈极显著负相关,相关系数为-0.782;小穗长与颖长度呈极显著正相关,相关系数为0.748;颖长度与植株高度呈显著正相关,相关系数为0.527。表型性状间的相关关系反映了新麦草植物发育过程中各部位差异化协调发育的特点〔19〕。
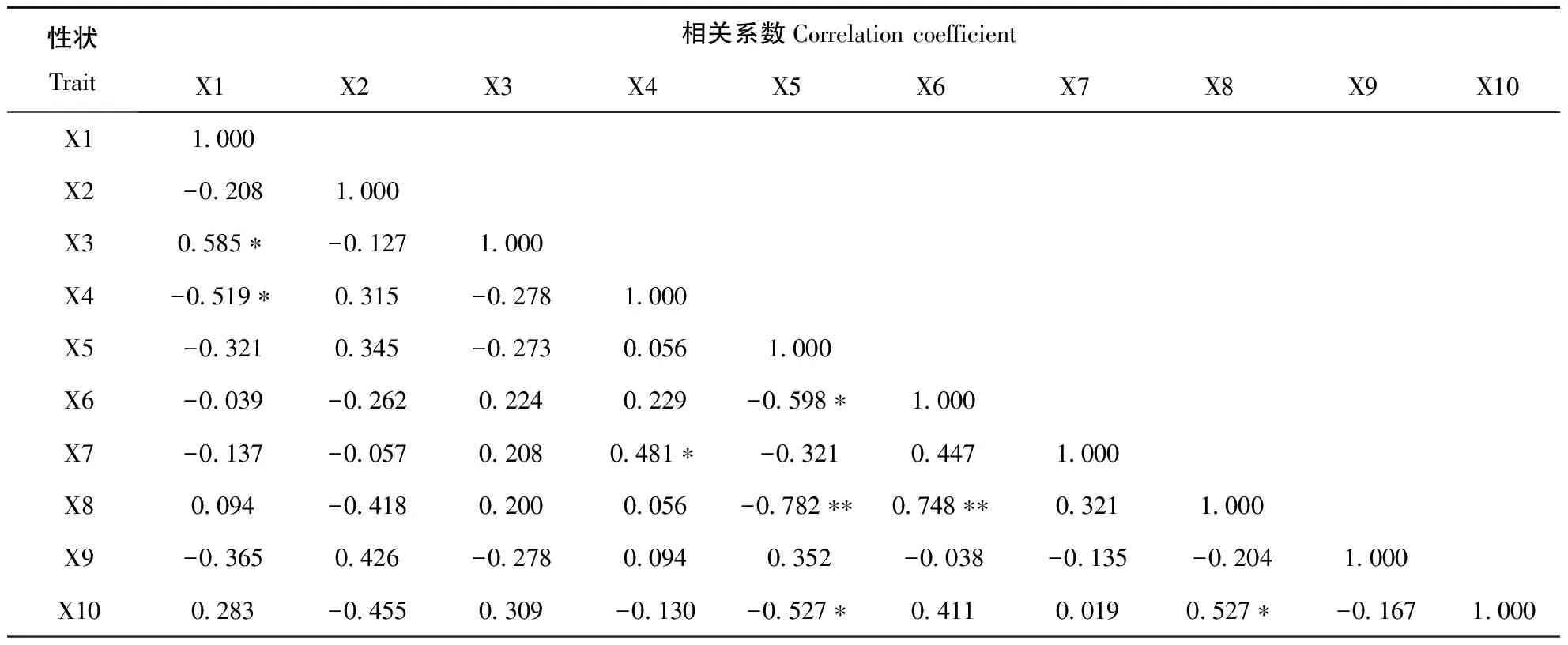
表4 新麦草表型性状间的相关关系
X1:茎干节数;X2:叶舌长度;X3:叶片长度;X4:叶片宽度;X5:小穗数;X6:小穗长;X7:小花数;X8:颖长度;X9:颖果长度;X10:植株高度。
X1:Stem section number;X2:Tongue length;X3:Leaf length;X4:Leaf width;X5:Number of spikelets;X6:Spikelet length;X7:The number of flowers;X8:Small length;X9:Caryopsis length;X10:Plant height.
3 结果与讨论
新麦草是禾本科小麦族的重要物种,作为放牧刈割兼用型优良的牧草,在我国北部干旱地区草地畜牧业退耕和草地生态重建中发挥了重要作用。新麦草作为一种很有推广前景的饲用牧草,其生产和利用价值越来越得到人们的关注。表型变异研究是遗传多样性研究的基础和先导〔20〕。表型性状受生境条件和空间距离的双重影响〔21〕。新麦草属植物的演化十分复杂,在长期演化及生境变化的过程中,各物种基因库形成于不同的自然选择压力,导致物种的演化式样存在巨大差别,并且可以体现在种内不同居群间的形态分化上〔22〕。本研究表型多样性丰富,各性状在材料间和材料内存在着广泛变异,这可能是自身遗传因素和环境因子共同作用的结果。陈仕勇等采用酸性聚丙烯酰胺凝胶电泳技术 (A-PAGE) 对来自中国新疆、俄罗斯西伯利亚及蒙古等地的35份野生新麦草种质进行醇溶蛋白的遗传变异分析,认为新麦草的遗传变异83.42%存在于类群内,16.58%存在于类群间,这与新麦草是异花授粉的繁育方式相关,种质间的大量基因流增加了种质间的变异,认为中国新疆、俄罗斯西伯利亚及蒙古地区是新麦草种质重要的分化中心,特别是中国新疆类群应是新麦草资源保护的重点地区〔23〕。本研究中来自于新疆、陕西、美国、哈萨克、玻利维亚和吉尔吉斯的11份新麦草,分布范围较为广泛,说明新麦草种质具有较强的环境适应能力,从11份新麦草的不同表型性状的遗传多样性入手,发现其存在着丰富的多样性,具有很大的育种潜力。本研究中10个性状指标的差异度主要有茎干节数、叶片宽度、小穗长、小花数和颖果长度等,主成分分析前3个主成分的累计贡献率为83.340%,反映个所有表型性状的大部分相关信息,小穗长、小花数、颖长度和植株高度等性状是造成新麦草种质资源表型差异的主要因素,也是新麦草种质育种选择过程中主要考虑的形态指标。从聚类结果来看,10个表型性状变异相同或地理类别相同与相近的材料大部分聚在一起,但也有交叉,这可能与材料来源地的地理生境和种质材料内在的遗传因素有关,然而这些表型性状易受外界因素的影响,反映的信息具有局限性,将新麦草种植在同一地区,减少了生态环境对其田间形态变异的影响,使田间形态变异更加客观的反映其遗传变异的情况〔24〕。关于新麦草种质的遗传变异需要将多种分析方法相结合进行综合分析,了解其遗传分化等为该种质资源的保护和利用提供更多的信息。新麦草的育种工作在我国处于起步阶段,通过对野生新麦草资源的长年驯化培育出新的品种是我国新麦草育种的重要手段,引种驯化也是育种的一种手段,一般是从加拿大、美国引进新麦草品种,在内蒙古地区进行试验,筛选出适合本地种植的优良品种。
〔1〕Jensen K B,Zhang Y F, Dewey D R. Mode of pollination of perennial species of the Triticeae in relation to genomically defined genera〔J〕. Canadian Journal of Plant Science, 1990, 70: 215-225.
〔2〕云锦凤,王勇,徐春波,等.新麦草新品系生物学特性及生产性能研究〔J〕.中国草地学报,2006,28(5)1-7. Yun J F,Wang Y,Xu C B.et al. The Biological Characters and Production Performance of a New Russian Wildrye Strain〔J〕. Chinese Journal of Grassland,2006,28(5):1-7.
〔3〕于晓丹,张蕴薇.新麦草研究进展〔J〕.草业与畜牧,2010,1:5-8. Yu X D,Zhang Y W. Research Progress ofPsathyrostachysjuncea〔J〕. Cao Ye Yu Xu Mu,2010,1: 5-8.
〔4〕Wei J Z,Campbell W F,Wang R RC.Genetic variability in Russian wildrye(Psathyrostachys juncea)assessed by RAPD〔J〕.Genetic Resources and Crop Evolution, 1997, 44: 117-125.
〔5〕齐晓芳,张新全,凌瑶,等.野生狗牙根种质资源的AFLP遗传多样性分析〔J〕.草业学报,2010,19(3):155-161. Qi X F,Zhang X Q,Lin Y,et al. Genetic diversity of wild Cynodon dactylon germplasm detected by AFLP markers〔J〕.ActaPratacultulturaeSinica,2010, 19(3): 155-161.
〔6〕Wei J Z, Campbell W F, Wang R R C. Allozyme variation in accession of Russian wildrye〔J〕. Crop Science, 1996, 36: 785-790.
〔7〕刘永财,孟林,张国芳,等.新麦草种质遗传多样性的ISSR分析〔J〕.华北农学报,2009,24(5):107-112. Liu Y C,Meng L,Zhang G F.et al.Genetic Diversity Analysis of 15Psathyrostachysjuncea Germplasm Resources by ISSR Molecular Marker〔J〕.Acta Agriculturae Boreali Sinica,2009, 24(5): 107-112.
也可以从经济基础部分质变引起的上层建筑部分质变来界定“中尺度时代”。如古罗马从王政时代转变为共和时代、帝国时代。中国封建社会从诸侯分封制时代转变为中央集权的郡县制时代。资本主义社会从特权选举制时代转变为普选制时代。社会主义社会从“阶级斗争为纲”的政治体制时代转变为“依法治国,建设社会主义法治国家”的现代政治体制时代。
〔8〕杨艳,韩建国,孙彦,等.新麦草遗传多样性等位酶分析〔J〕.草业科学,2007,24(8):59-63. Yang Y,Han J G,Sun Y, et al.Allozyme genetic diversity analysis ofPsathyrostachysjuncea〔J〕.Pratacultural Science,2007, 24(8): 59-63.
〔9〕Yang Y, Han J, Berdahl J D. Allozyme diversity of Russian wildrye accessions〔J〕. Plant Breeding, 2008, 127:480-484.
〔10〕汪生泉,王赟文,韩建国,等.氮肥对新麦草穗颈解剖性状的影响〔J〕.江西农业大学学报,2007,29(6):899-902. Wang S Q.Wang Z W.Han J G,et al. The Efect of Nitrogenon Morphological Structure of Panicle Neck of Psathyrostachys juncea 〔J〕.Acta Agriculturae Universitatis Jiangxiensis,2007,29(6):899-902.
〔11〕孙彦,史德宽,杨青川.施肥和刈割对新麦草产草量影响的研究〔J〕.草地学报,1998,6(1):11-19. Sun Y.Si D K.Yang Q C.Studies tne Effects of Nitrogen,Phosphate,Potassium Fertilizer and Clipping Frequency on tne Yield of Russian Wildrye Grass〔J〕.Acta Agrestia Sinica,1998,6(1):11-19.
〔12〕韩建国,宋锦峰,张蕴薇.放牧强度对新麦草生产特性和品质的影响〔J〕.草地学报,2000,8(4):312-318. Han J G,Sun J F,Zhang Y W.Effects on prodution characters and quality of Russian wildrye Grass under Different Grazing Intensities〔J〕.Acta Agrestia Sinica, 2000,8(4):312-318.
〔13〕Wie JunZhi,Campbell WF et al.Standard Giemsa Cbanded karyotype of Russian wildrye(Psathyrostachys juncea)andits use in identification of a deletion translocationheterozygote〔J〕.Genome,1995,(38):1262-1270.
〔14〕李俊琴,云锦凤,云岚,等.新麦草幼胚愈伤组织诱导〔J〕.中国草地学报,2008,30(2):39-42. Li J Q,Yun J F,Yun L,et al.Callus Induction of Immature Embryo of Psathyrostachys juncea〔J〕.Chinese Journal of Grassland,2008,30(2):39-42.
〔16〕张秀丽,侯建华,云锦凤,等.山丹新麦草多倍体诱导的初步研究〔J〕.中国草地,2005,27(1):34-38. Zhang X L, Hou J H,Yun J F,et al.Chromosome Doubling Induction of Psathyrostachys perennis Keng cv .Shandan〔J〕.Grassland of China,2005,27(1):34-38.
〔17〕Wang ZY,Bell J,Lehmann D. Transgenic Russian wildrye(Psathyrostachys juncea) plants obtained by biolistictransformation of embryogenic suspension cells〔J〕. PlantCell Reports. Springer Verlag,Berlin,Germany,2004,(22):903-909.
〔18〕Guo B L,Yang J X,Li Y C,Yu S S.The application of principal component analysis on mainly economic characters and superior variety selection of apricot for nucleolus using〔J〕.ScientiaSilvaeSinicae,2000,36(6) :53-56.
〔19〕蒋林峰,张新全,付玉凤,等.中国主要鸭茅品种农艺性状变异研究〔J〕.草业学报,2015,24(3):142-154. Jiang Linfeng,Zhang Xinquan,Fu Yufeng.Agronomic trait variation of some main orchardgrass(Dactylis glomerata) cultivars in China〔J〕.ActaPratacultulturaeSinica, 2015,24(3):142-154.
〔20〕黄春琼,刘国道,白昌军,等.475份狗牙根种质资源形态多样性的研究〔J〕.草业学报,2012,21(4):33-42. Huang Chunqiong,Liu Guodao, Bai Changjun.A study on the morphological diversity of 475 accessions of Cynodon dactylon〔J〕.ActaPratacultulturaeSinica, 2012,21(4):33-42.
〔21〕袁庆华,张吉宇,张文淑,等.披碱草和老芒麦野生居群生物多样性研究〔J〕.草业学报,2003,12(5):44-49. Yuan Q H,Zhang J Y,Zhang W S,et al.Biodiversity of native populations of Elymus dahuricus and Elymus sibiricus.Acta Prataculturae Sinica,2003,12(5):44-49.
〔22〕杨瑞武,周永红,郑有良.披碱草属、鹅观草属和猬草属模式种的形态学变异和酯酶同工酶分析〔J〕.四川农业大学学报,2000,18(4):291-295. Yang R W,Zhou Y H,Zheng Y L.Comparison of morphology and esterase isozyme among typespecies of Elymus,Roegneria and Hystrix(Poaceae:Triticeae).Journal of Sichuan Agricultural University,2000,18(4):291-295.
〔23〕陈仕勇,马啸,张新全,等.野生新麦草种质醇溶蛋白遗传变异分析〔J〕.草业学报,2011,20(4):311-315. Chen S Y, Ma X, Zhang X Q, Genetic variation analysis of gliadin from wild germplasm of Psathyrostachys juncea〔J〕.ActaPratacultulturaeSinica, 2011,20(4):311-315.
〔24〕李陈建,付彦博,万江春,等.30份苏丹草种质资源农艺性状的遗传多样性分析〔J〕.草业科学,2015,32(1):85-93. Li C J,Fu Y D,Wan J C,et al. Genetic diversity of agronomic characteristics of 30 Sorghum sudanense germplasm〔J〕. Pratacultural Science,2015,32(1):85-93.
Principal Component and Cluster Analysis of Phenotypic Diversity of Russian wild rye Germplasm Resources
Hao Linfeng1, Li Zhiyong2,Li Jun2, Huang Fan2,Li Hongyan2,*
(1.Baotou City Livestock Improvement Workstation , Baotou City 014010;2.Grassland Research Institute of CAAS, Hohhot 010010)
The stem segments, leaf length, leaf width, spikelet number, spikelet length, number of florets, spikelet number, spikelet number, spikelet number, spikelet number, spikelet number and leaf width of 11 new wild wheat straws from Xinjiang, Shaanxi, America, Kazakhstan, LibyaBolivia and Kyrgyzstan, The length of the caryopsis, the length of the caryopsis and the height of the plant. The genetic diversity was analyzed by means of variance analysis, principal component analysis and cluster analysis, so as to screen out the excellent germplasm and provide the reliable basis for Russian wild rye breeding.The results showed that: 1) The phenotypic traits of Russian wild rye germplasm were highly variable, the coefficient of variation was 14.6% -68.0%, the number of florets was the largest and the leaf width was the smallest; 2) 10 phenotypic traits could be classified as 3 main characters The length of the spikelet, the length of the florets, the length of the plant and the height of the plant were the main characters which affected the phenotypic variation of the new wheat straw; 3) There were significant or extremely significant positive and negative correlations between 10 phenotypic traits. 4) Eleven Russian wild rye materials were divided into 3 groups by Euclidean distance system clustering method. This study revealed morphological genetic diversity of Russian wild rye, which laid a theoretical foundation for the collection, identification, evaluation, conservation and utilization of Russian wild rye resources.
Russian wild rye; Phenotypic traits; Cluster analysis
S543+.9
A
2095—5952(2017)01—0023—07
2016-12-18
农作物种质资源保护项目(2015NWB037)
郝林峰(1964-),男,内蒙古巴彦淖尔市五原县,高级农艺师,主要从事畜牧业生产、家畜改良、育种及农村牧区农济管理研究。E-mail:btglz2008@163.com。
[*通讯作者]李鸿雁,研究员,博士,硕士生导师,主要从事牧草种质资源保存、创新与育种研究。 E-mail:hongyli1964@126.com.
