西藏拉萨城区黄嘴朱顶雀的巢及巢址选择
2017-04-08周生灵普布次仁
周生灵, 普布, 次仁*
(西藏自治区高原生物研究所,西藏大学,拉萨850000)
西藏拉萨城区黄嘴朱顶雀的巢及巢址选择
周生灵1, 普布2, 次仁2*
(西藏自治区高原生物研究所,西藏大学,拉萨850000)
2015年3—6月,在拉萨城区采用样方法和直接观察法对黄嘴朱顶雀Carduelisflavirostris的巢址选择进行了调查研究。共收集了32个巢,其中93.75%的巢筑在乔木上,雪松Cedrusdeodara占65.63%,侧柏Platycladusorientalis占12.50%,青海云杉Piceacrassifolia占12.50%,垂柳Salixbabylonica占3.13%,6.25%的巢筑在灌木紫叶李Prunuscerasifera上。黄嘴朱顶雀的巢多筑于巢树主干与侧枝的交界处,且在巢树上的位置以向南为主,占40.6%,其次是向东,占28.1%。通过主成分分析,发现影响黄嘴朱顶雀巢址选择的主要环境因子包括巢树树种、巢树高度、巢树胸径、木本植物的覆盖度、巢离水源与公路的距离、巢离巢树主干的距离以及巢向。
黄嘴朱顶雀;拉萨城区;巢址选择
鸟类巢址研究是对鸟巢以及巢周围的生态因子在鸟类选择巢址过程中的作用和地位的研究,从而揭示鸟类选择某个区域作为巢址的主导因素和影响因子,对保护和利用鸟类资源具有重要的理论和实践意义,并且巢址选择对鸟类的生存和繁殖起着决定性的作用(Badyacv,1995;丁长青,郑光美,1997)。鸟类巢址选择的一个非常重要意义是避免被捕食(包括亲鸟、卵、雏鸟)、寄生虫感染、巢破坏等(Eggersetal.,2006;Watsonetal.,2006;Lima,2009);巢址环境还会影响到鸟类的胚胎发育、雏鸟发育及亲鸟在繁殖期间的安全和体能消耗等(Kim & Monaghan,2005;D'Albaetal.,2011)。
黄嘴朱顶雀Carduelisflavirostris隶属于雀形目Passeriformes燕雀科Fringillidae朱顶雀属Carduelis,是典型的高山鸟类,分布海拔2 500~5 000 m,部分甚至在海拔5 000 m以上的高原地区。该物种的繁殖期基本上集中在3月中旬至7月中旬,受栖息地环境条件影响较大,巢址选择有明显的地区差异。据记载,欧洲黄嘴朱顶雀的巢址一般选择在石南属Photinia植物、欧洲蕨Pteridiumaquilinum、羊胡子草属Eriophorum和针叶幼林等植物上,但对其具体的选择依据并不清楚(Josepetal.,1999);此外,也有人认为它筑巢于石缝中、荆豆属Ulex植物上、草丛及墙缝中,偶尔也筑巢于啮齿目Rodentia动物的洞穴中(Noel,1973;Nicholasetal.,2010);有研究发现,黄嘴朱顶雀在英国的分布范围与数量均有所下降(Dereketal.,2010)。国内学者分别对西藏北部当雄地区和青海海北地区黄嘴朱顶雀的繁殖生态做了初步的研究,主要集中在窝卵数、卵大小以及雏鸟生长情况等方面(张晓爱等,2006;Luetal.,2011),对巢址选择的研究不是很详细。因此,本研究选择在拉萨城区对黄嘴朱顶雀的巢及巢址选择进行了详细的研究,为鸟类的保护和利用提供一定的基础数据;同时与其他地区的研究结果进行对比,揭示鸟类对不同环境的适应性策略。
1 研究地自然状况
拉萨城区平均海拔3 650 m,地处91°10′27.67″~91°10′57.90″E,29°38′39.03″~29°38′59.01″N,为雅鲁藏布江支流拉萨河中游河谷平原,日照长、辐射强,年温差较小,日温差较大,受下沉气流的影响,全年多晴朗天气,冬无严寒,夏无酷暑,属高原季风半干旱气候,年平均气温7.5 ℃,最冷月均温-2.2 ℃,最热月均温15.5 ℃。拉萨地区年平均降水量444.8 mm,其中6—9月占92.4%,多夜雨,夜雨量最高达到84%,平均相对湿度45%,年平均蒸发量2 205.6 mm,太阳年总辐射值202.4 kCal/(cm2·年),全年无霜期133 d(王晓军,程绍敏,2009)。研究区域内的植被主要是人工次生林,优势树种主要有雪松Cedrusdeodara、紫叶李Prunuscerasifera、青海云杉Piceacrassifolia、银白杨Populusalba、槐Sophorajaponica、乌柳Salixcheilophila、垂柳Salixbabylonica等;常见的草本植物有白草Pennisetumflaccidum、波斯菊Cosmosbipinnata、茎直黄芪Astragalusstrictus、蒲公英Taraxacummongolicum等,可为黄嘴朱顶雀提供食物来源;常见的雀形目鸟类包括树麻雀Passermontanus、灰背伯劳Laniustephronotus、白鹡鸰Motacillaalba、褐岩鹨Prunellafulvescens、鸲岩鹨P.rubeculoides等,其中灰背伯劳为黄嘴朱顶雀的天敌之一;在冬季,褐岩鹨和鸲岩鹨均有与黄嘴朱顶雀混群的现象。
2 研究方法
本研究主要采用了直接观察法、样方法和定点观察法(借助SWAROVSKI 10×30双筒望远镜)。寻巢采用直接观察法,即在研究区域内匀速前进,发现黄嘴朱顶雀成鸟(包括鸣叫和活动)便开始跟踪观察,直到发现巢;每发现一个巢,用自制的布质标签和记号笔在巢树上做双重标记,并用GPS进行定位,记录其精确的地理位置,用GPS定位数据在Google Earth上绘制巢点分布图,并根据巢点分布图上的比例尺计算巢之间的距离。巢址环境变量的测定采用样方法,测量的环境变量包括11个:巢树树种、巢树郁闭度、巢离地面高度、巢树高度、巢树胸径、巢向、巢离主干的距离、巢树离公路的距离、巢树离水源的距离、木本植物的覆盖度(5 m×5 m)、草本植物的覆盖度(1 m×1 m);并在黄嘴朱顶雀筑巢结束后进行巢的测量,主要变量有巢高、内径、外径以及巢质量(繁殖期结束后收集所有的繁殖巢,晒干称重),并分析巢材的主要组成;最后在研究区域内,随机选取12个10 m×10 m的样方,进一步确定研究区域内的木本植物资源状况,包括物种组成及其数量。筑巢情况采用定点观察法,即从08∶00—20∶00,对正在筑巢的黄嘴朱顶雀进行定点观察,记录其叼巢材的行为、频率及取材地点。
数据分析采用R i386 3.1.1,主要进行基础分析和主成分分析(PCA)。巢树的选择系数采用被修改的Ivlev’s的Selectivity index:Ei=(ri-ni)/(ri+ni)。为了便于统计分析,给巢树树种和巢在巢树主干上的方向赋予一定的数值:雪松=1,青海云杉=2,侧柏=3,紫叶李=4;同样,东、西、南、北巢向分别赋予1、2、3和4。
3 结果
3.1 黄嘴朱顶雀巢的分布及基本特征
通过对32个巢的分析发现,黄嘴朱顶雀在拉萨城区的巢并非随机分布,而是具有一定的选择性。巢间距最小为3.2 m,最大达到2 010 m(图1)。
黄嘴朱顶雀巢大多为筑在乔木上的编织巢(浅杯状巢),巢材主要为巢附近的干树枝和枯草,巢内部铺垫有人体和动物的毛发、布条、羽毛以及棉花等比较柔软的材料,巢的编织比较精致。观察发现,编织顺序是自上而下、由外及内(n=8),巢的基本特征见表1。
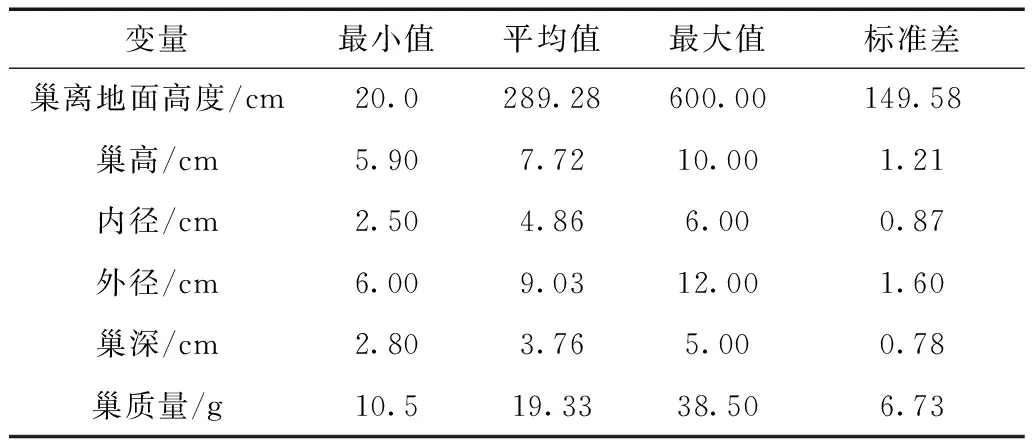
表1 西藏拉萨城区黄嘴朱顶雀巢的基本特征Table 1 Basic nest characteristics of Carduelis flavirostrisin urban area of Lhasa
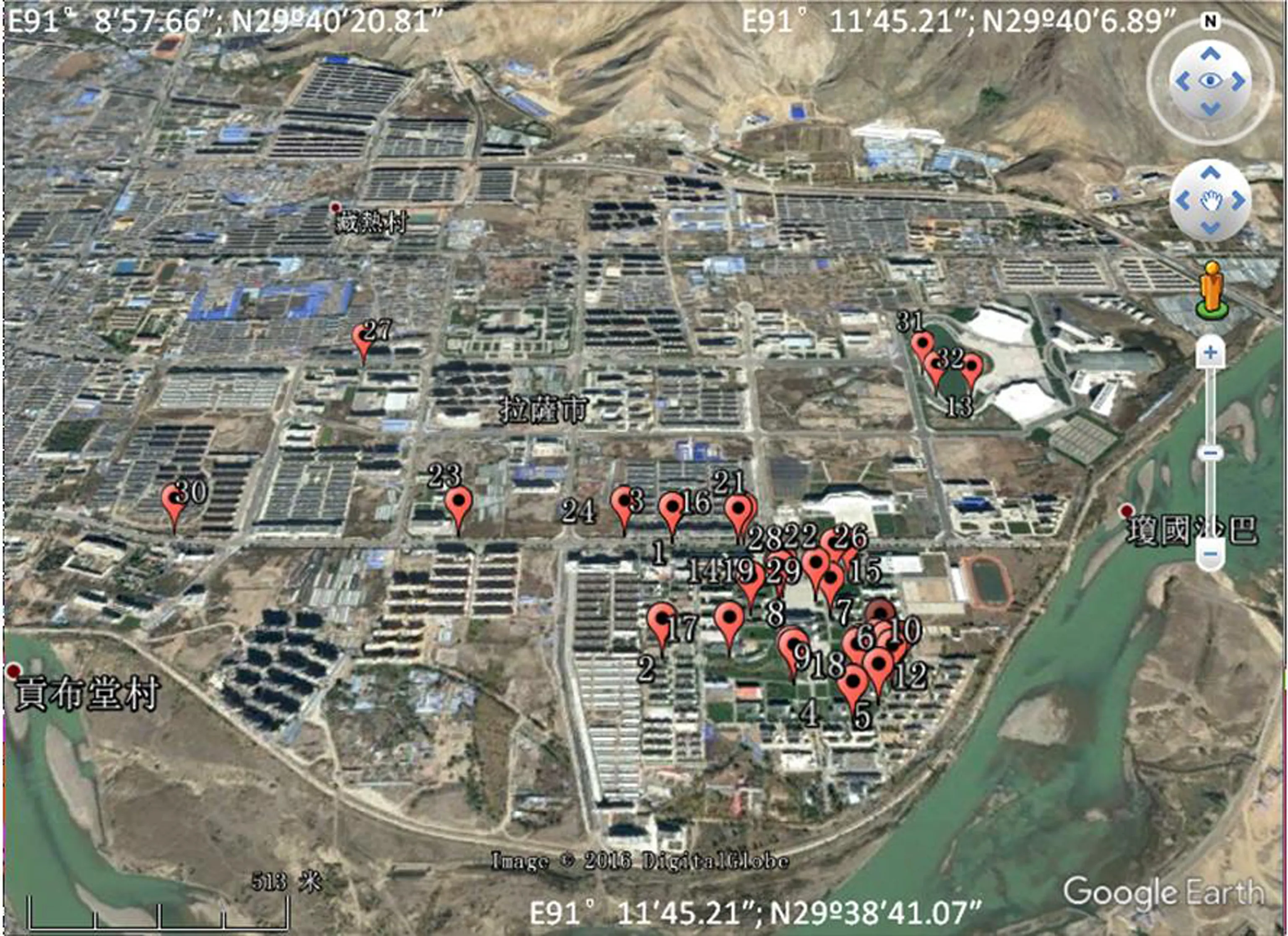
图1 研究区域内黄嘴朱顶雀巢址分布图
Fig. 1 Distribution of nests in the study area ofCarduelisflavirostris
拉萨城区黄嘴朱顶雀的繁殖活动为3月中旬至6月中旬。黄嘴朱顶雀巢址选择由双亲共同承担,雌鸟更具有主动性,筑巢主要由雌鸟承担,在整个筑巢过程中,雌鸟负责叼巢材、编织巢;雄鸟负责陪同与警觉,并具有明显的领域行为(驱赶侵入巢区的鸟,包括同种和异种的鸟类)。
3.2 巢树及巢向的选择
研究区域内共有11种人工种植的木本植物,雪松和垂柳是优势种,其次为银白杨、侧柏、乌柳、槐及圆柏等。黄嘴朱顶雀65.63%(n=32)的巢筑在雪松上,侧柏和青海云杉上的均占12.50%。在32对研究对象中,有2对亲鸟选择在紫叶李上筑巢;有1对亲鸟在垂柳上筑巢,但仅筑了2天就放弃了,推测是筑假巢或躲避天敌。在黄嘴朱顶雀巢址选择过程中巢树选择系数比较高的有雪松(0.436)、青海云杉(0.319)、紫叶李(0.319)、侧柏(0.319),但也有部分植物是黄嘴朱顶雀选择巢址时避开的物种,例如:垂柳(-0.784),乌柳、槐、银白杨、梨、榆及圆柏(均为-1)(表2)。根据32个巢在巢树上的位置来看,黄嘴朱顶雀78.13%的巢筑在巢树主干和侧枝的交界处,位于巢树主干南边的巢占40.6%,位于东边的占28.1%,北边和西边分别占21.9%和9.4%。
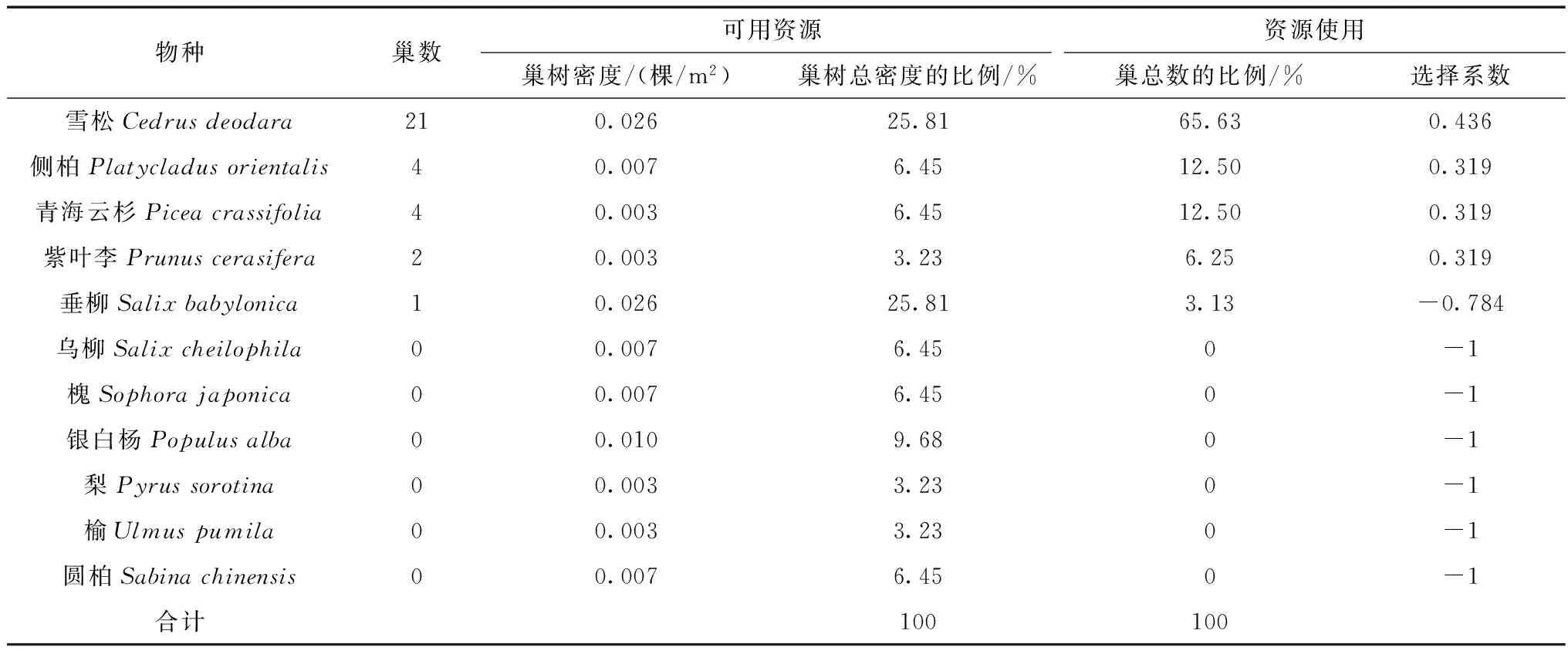
表2 黄嘴朱顶雀在拉萨城区巢树选择Table 2 Species of trees selected for nest site by Carduelis flavirostris in urban area of Lhasa
3.3 主成分分析
对巢树树种、巢树郁闭度、巢树胸径、巢树离水源的距离、草本和木本植物的覆盖度等11个环境变量进行了主成分分析,结果显示,前3个主成分能够解释黄嘴朱顶雀巢址选择变量的76.70%。前4个主成分的转置矩阵结果显示巢树树种、巢树高度、巢树胸径、木本植物的覆盖度对主成分1贡献较大,对主成分2贡献较大的变量包括巢离巢树主干的距离、巢树离水源的距离和巢树离公路的距离,只有巢向对主成分3有较大贡献(表3)。
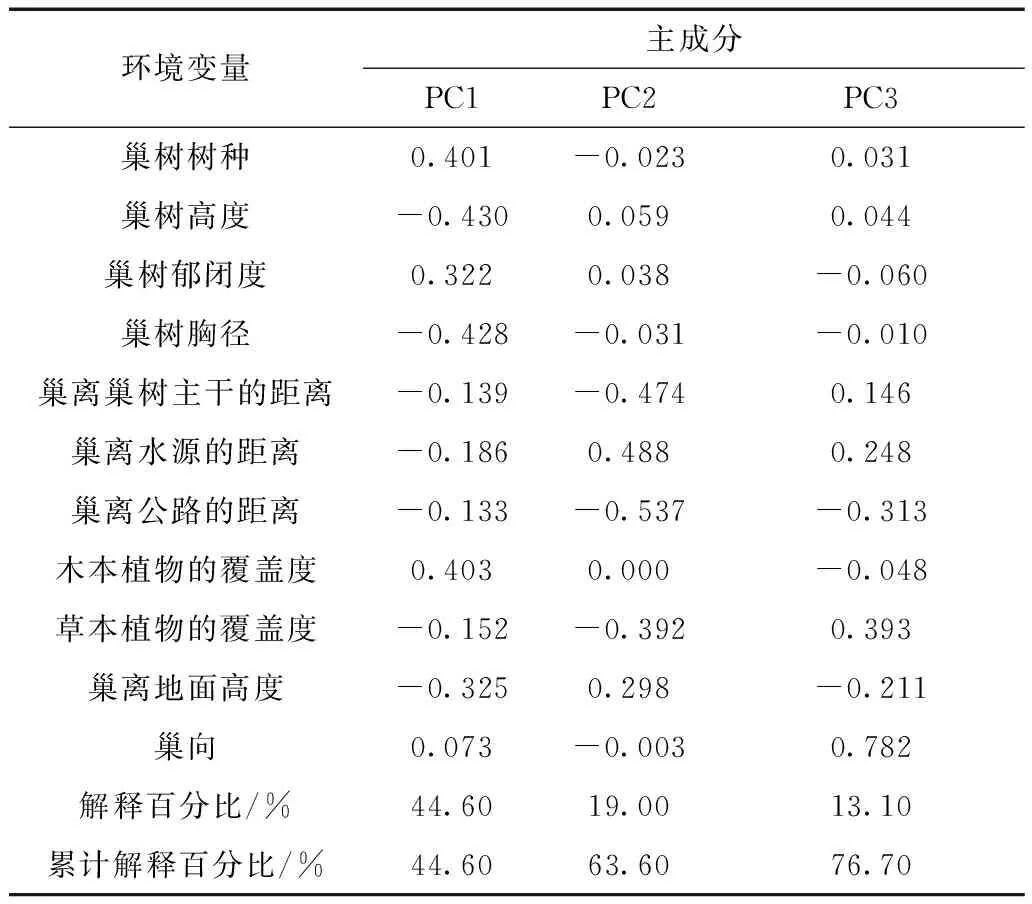
表3 黄嘴朱顶雀巢址选择的原始变量与各主成分之间的相关系数Table 3 Correlation coefficients between original variables and principal components in nest site selection of Carduelis flavirostris
4 讨论
4.1 一般生境选择
巢址选择是鸟类整个繁殖活动的最基本、最重要的步骤。相对适宜的繁殖环境和巢址,对鸟类的繁殖至关重要,直接影响鸟类的繁殖成功率,许多鸟类一般都会选择在繁殖成活率最高且繁殖代价最小的区域筑巢,确保繁殖活动的顺利进行(张正旺等,1994)。黄嘴朱顶雀雌鸟在巢址选择上更具主动性,这一现象符合鸟类巢址选择的一般规律(丁长青,郑光美,1997)。
本研究结果显示,拉萨城区黄嘴朱顶雀的巢树主要选择雪松、侧柏、青海云杉、紫叶李、垂柳等。在研究区域内,雪松和垂柳的密度基本相同,两者均属于优势种,黄嘴朱顶雀选择雪松作为巢树却避开垂柳,分析其主要原因可能是雪松的树叶是硬质针叶形,长枝散生,短枝簇生,此枝叶结构提高了巢的隐蔽性,同时也可以阻碍天敌的攻击;而垂柳枝叶细长,柔软下垂,对成鸟和巢能够提供的保护远不如雪松;因此,即使垂柳与雪松有相同的密度,黄嘴朱顶雀在选择巢址时依然趋向于雪松而避开垂柳。
4.2 影响巢址选择的主要因子分析
主成分分析可以从多指标的复杂事物中找出生命活动的主要倾向,帮助了解生命活动的规律和本质。通过主成分分析,发现黄嘴朱顶雀前3个主成分的累计贡献率达到了76.70%,结果显示影响黄嘴朱顶雀巢址选择的主要环境因子有巢树高度、巢树胸径、巢离水源和公路的距离、巢离巢树主干的距离以及巢向。据相关报道,鸟类对特定的栖息地(包括巢址)的偏爱是由鸟类的形态结构和行为功能及其栖息地中成功获取食物等必要生存条件决定的,而景观的结构特征、取食和营巢机会以及其他物种的存在是鸟类出现栖息地选择的直接诱因(Cody,1985;张正旺,郑光美,1999)。黄嘴朱顶雀属于小型雀形目鸟类,选择大小适中的雪松、青海云杉等作为巢树,具有一定的选择机理,过于粗壮的树木一般都被山斑鸠Streptopeliaorientalis、灰背伯劳等占领(包括筑巢和其他活动);过于细小的树木又不足以提供隐蔽度和安全性。黄嘴朱顶雀选择离水源和公路的距离较为适中的区域作为巢址,既可以在繁殖期间满足水分和食物来源(藏族群众喜欢在公路上投放糌粑等食物来喂养流浪狗,鸟类也可从中取益)的需求,又可以避免因距离太近而造成人为的破坏。拉萨城区黄嘴朱顶雀巢多建筑于巢树主干与侧枝的交界处,这样可以防止巢遭到恶劣天气的破坏,并以向南为主,充足的光照和较高的温度是确保卵顺利孵化的重要因素,同时也影响雏鸟的生长发育(高玮等,2003),而且拉萨城区多西北风,向南是背风向,可以防止大风大雨天气对巢的破坏。
4.3 不同地域巢大小差异的原因
比较不同地域黄嘴朱顶雀巢大小发现,当雄最大,其次为海北,再次是拉萨,最后是英国(Josepetal.,1999;Luetal.,2011;张晓爱等,2006)(表4)。显然,黄嘴朱顶雀的巢大小与地区的海拔基本一致。高海拔地区,年平均气温较低、风大,大而重的巢有助于保温和抵挡风力破坏;相反,低海拔地区年平均气温适宜,小而轻巧的巢既可以节省亲鸟的能量,又有助于隐蔽。虽然海北的海拔(3 200~3 600 m)比拉萨的低(3 650 m),但海北为高寒草甸生态系统,气温低、风力大,因此,海北的巢较拉萨的巢大。
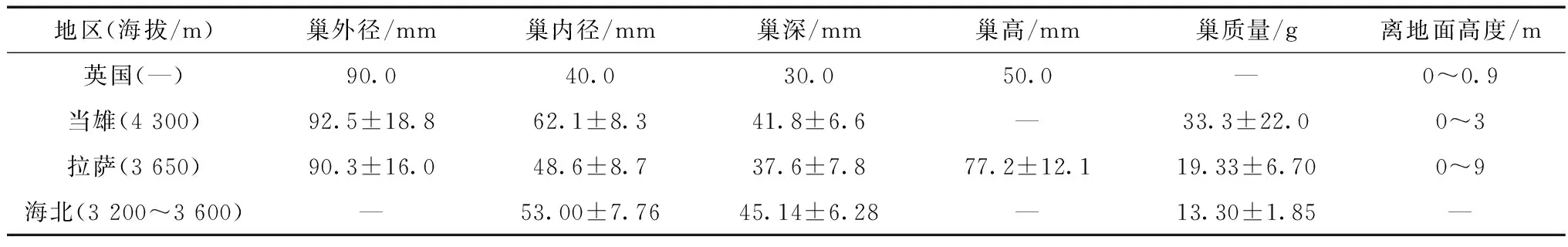
表4 不同地域黄嘴朱顶雀巢的参数Table 4 The nest characteristics of Carduelis flavirostris in different areas
致谢:西藏高原生物研究所副研究员杨乐在文章修改中提出建设性意见,在此表示感谢!
丁长青, 郑光美. 1997. 黄腹角雉的巢址选择[J]. 动物学报, 43(1): 27-33.
高玮, 王海涛, 孙丹婷. 2003. 栗斑腹鹀的栖息地和巢址选择[J]. 生态学报, 23(4): 665-672.
王晓军, 程绍敏. 2009. 西藏主要气候特征分析[J]. 高原山地气候研究, 29(4): 81-84.
张晓爱, 刘泽华, 赵亮, 等. 2006. 青藏高原常见雀形目鸟类的筑巢特征[J]. 动物学研究, 27(2): 113-120.
张正旺, 梁伟, 盛刚. 1994. 斑翅山鹑巢址选择的研究[J]. 动物学研究, 45(4): 37-43.
张正旺, 郑光美. 1999. 鸟类栖息地选择研究进展[C]// 中国动物学会. 中国动物科学研究——中国动物学会第十四届会员代表大会及中国动物学会65周年年会论文集.
Badyacv AV. 1995. Nesting habitat and nesting success of eastern wild turkeys in the Arkansas Ozark Highlands[J]. The Condor, 97(1): 221-232.
Cody ML. 1985. Habitat selection in birds[M]. London: Academic Press.
D’Alba L, Spencer KA, Nager RG,etal. 2011. State dependent effects of elevated hormone, nest site quality, corticosterone levels and reproductive performance in the common eider[J]. General & Comparative Endocrinology, 172(2): 218-224.
Derek M, Chris B, Bryony W,etal. 2010. The movement patterns of two populations of twitesCarduelisflavirostrisin Ireland[J]. Ringing & Migration, 25(1): 15-21.
Eggers S,Griesser M, Ekman J. 2006. Predation risk induces changes in nest-site selection and clutch size in the Siberian jay[J]. Philosophical Transactions of the Royal Society of London series B-Biological Sciences, 273(1587): 701-706.
Josep del H, Andrew E, David C. 1999. Handbook of the birds of the world vol 5[M]. Barcelona: Lynx Edicions: 625-639.
Kim SY, Monaghan P. 2005. Effects ofvegetation on nest microclimate and breeding performance of lesser black-blackd gulls (Larusfuscus)[J]. Journal of Ornithology, 146(2): 176-183.
Lima SL. 2009. Predators and the breeding bird: behavioral and reproductive flexibility under the risk of predation[J]. Biological Reviews, 84(3): 485-513.
Lu X, Guo YY, Liang JJ,etal. 2011. Breeding ecology of the twiteCarduelisflavirostrisin northern Tibet[J]. Ornis Fennica, 88(3): 172-178.
Noel O. 1973. Breeding distribution of the twite in central Britain[J]. Bird Study, 20(2): 51-62.
Nicholas IW, Jeremy DW. 2010. Breeding ecology of twiteCarduelisflavirostrisin a crofting landscape[J]. Bird Study, 57(2): 142-155.
Watson M, Wilson JM, Koshkin M,etal. 2006. Nest survival and productivity of the critically endangered sociable lapwingVanellusgregarious[J]. Ibis, 148(3): 489-502.
Preliminary Report on Nest-site Selection byCarduelisflavirostrisin Urban Area of Lhasa
ZHOU Shengling1, PU Bu2, Tsering Dorge2*
(Tibet Plateau Institute of Biology, Tibet University, Lhasa 850000, China)
The nest-site selection of twite (Carduelisflavirostris) was investigated and studied by quadrate sampling and direct observation method in Lhasa urban area from March to June in 2015. A total of 32 nests were collected, and 93.75% of which were set on macrophanerophytes, with 65.63% were on theCedrusdeodara, 12.50% onPlatycladusorientalis, 12.50% onPiceacrassifolia, 3.13% onSalixbabylonica, and 6.25% on bush ofPrunuscerasifera. The nests of twite were mainly set at the junction of the trunk and lateral branches, and most of the nests were on the south part of nesting trees, which accounted for 40.6%, while 28.1% were on the east part. By principal component analysis, it was found that the main environmental factors which affectedC.flavirostris’s nest-site selection included nest tree species, nest tree height, nest tree diameter at breast height, the coverage of woody plants, the distance between nest tree to water and road, the distance from nest to the trunk of nest tree and nest direction.
Carduelisflavirostris; urban area of Lhasa; nest-site selection
2016-04-15 接受日期:2017-01-20
周生灵(1990—), 硕士, 实习研究员, 主要从事动物生态研究, E-mail:919374208@qq.com
*通信作者Corresponding author, E-mail:tseringdorge@163.com
10.11984/j.issn.1000-7083.20160084
Q959.7; Q958.1
A
1000-7083(2017)02-0188-05
致谢:感谢安徽大学青年骨干教师培养对象(第三批)和环境保护部生物多样性保护专项资助。
