杂色山雀体型、子代性别比与鸣唱特征的相关性研究
2017-04-08赵婷婷刘燕张志委霍亚鹏林景峰万冬梅殷江霞
赵婷婷, 刘燕, 张志委, 霍亚鹏, 林景峰, 万冬梅, 殷江霞*
(1.辽宁大学生命科学院辽宁省动物资源与疫病防治重点实验室,沈阳110036;2. 仙人洞自然保护区管理局,辽宁大连116400)
杂色山雀体型、子代性别比与鸣唱特征的相关性研究
赵婷婷1, 刘燕1, 张志委1, 霍亚鹏1, 林景峰2, 万冬梅1, 殷江霞1*
(1.辽宁大学生命科学院辽宁省动物资源与疫病防治重点实验室,沈阳110036;2. 仙人洞自然保护区管理局,辽宁大连116400)
鸣声在鸣鸟的生活史中具有重要作用,是其与外界进行信息交流最有效的方式之一。鸟类的鸣唱与其个体因素(如体型、子代性别比等)有着十分重要的联系。为了揭示个体体型、子代性别比与鸣唱特征之间的相关性,于2014—2015年3—6月在辽宁省仙人洞国家级自然保护区对杂色山雀指名亚种Parusvariusvarius的鸣声进行录制,测量了各体型参数,并对每巢雏鸟进行性别鉴定。结果显示:(1)杂色山雀体型与其鸣唱的最高频率具有显著相关性,但单一体型参数(体质量、跗蹠长度)与其鸣声参数相关性均不显著;(2)鸣唱语句长度与子代性别比具有显著相关性,子代性别比与其他鸣声参数均不存在显著相关性。本研究结果有助于进一步了解小型鸟类体型与鸣唱的关系,以及鸣唱与子代性别比之间的关系。
杂色山雀;鸣唱;体型;子代性别比
声信号在鸣鸟信息传递中具有重要作用,是鸣鸟生命活动与外界环境联系的枢纽(Weeden & Falls,1959)。鸟类鸣唱易受到多种因素的影响,如:种群数量、个体因素和环境因素等(Staiceretal.,1996)。个体体型与后代性比是鸟类重要的个体因素(Lislevandetal.,2007)。鸣唱是鸣鸟重要的第二性征,具有吸引配偶、保卫领域等作用(Beecher & Brenowitz,2005;Searcyetal.,2006;de Kortetal.,2009),研究表明雌鸟会通过雄鸟鸣唱来调节后代性比,雄鸟鸣唱频率越高,雄性子代越多(Dreissetal.,2006)。对鸟类而言,个体体型是其最重要的特征之一(Budka & Osiejuk,2013),有研究发现鸣鸟可以通过改变体型(尤其是体质量)影响发声器官进而改变鸣唱频率(Taylor & Reby,2010)。因此,研究鸟类体型、后代性比与鸣唱之间的相关性十分必要。
长期以来学者认为声音频率可以表征物种体型,主要是因为体型大小决定发声器官的大小,研究发现发声器官粗大的鸟类具有较低的声音频率(Fletcher,2004;Osiejuketal.,2005)。但是,最新研究发现由于物种之间的差异,鸟类体型与声音频率的相关性并非一致,如红腹灰雀Pyrrhulapyrrhula体型越大,声音频率越低(Potvin,2013);而灰眉岩鹀Emberizagodlewskii体型与声音频率不存在相关性(鲁思凡等,2014)。目前,行为学研究领域对于出现这种差异的解释还不够完善(Petersetal.,2009)。Patel等(2010)认为鸟类体型与鸣唱之间的相关性取决于个体体型变异系数,变异系数越大,相关性越显著。也有研究认为在小型雀形目Passeriformes鸟类中,鸣唱与体型之间不存在显著相关性(Cardoso,2011),为了更清楚地了解体型对鸟类鸣唱的影响,本研究选择小型雀形目鸟类杂色山雀Parusvariusvarius作为研究对象,探究其鸣唱与体型之间的相关性。
体型综合参数通常可以反映鸟类的身体状态(鲁思凡等,2014)。当个体处于较好的身体状态时,声音频率可能会发生变化(Cynxetal.,2005)。例如,一些物种喜欢利用鸣唱频率显示强大的竞争力(Araya-Ajoyetal.,2009),当物种处于良好的身体状态时,声音频率就会显著增高(Morton,2000)。因此,体型综合参数常被用作分析鸣唱与体型的相关性。此外,跗蹠、体质量等也是量化鸟类体型的重要指标(Zollinger & Brumm,2015),有研究发现一些物种为了改变鸣声频率而有意识地调节体质量,导致鸣声频率与体质量存在显著相关性(Appleby & Redpath,1997;Evansetal.,2006;Mageretal.,2007)。本研究通过对杂色山雀跗蹠、体质量和体型综合参数与鸣唱相关性的研究,为鸟类鸣声研究提供基础资料。
亲本的优良性状(如鸣唱)是决定子代质量的重要因素,也是决定后代性比的关键因素(Trivers & Willard,1973)。尽管这个机制还未被实验验证,但是过去的研究发现雄性的生存技能(如鸣唱)与后代性比存在显著相关性(Nageretal.,1999;Sheldon,1999)。当雄性鸣唱频率较高时,亲本会有意识地调节后代性比,增加雄性子代数量,这可能是由于亲本希望将优秀的鸣唱性状传递给雄性子代;相反,雄性鸣唱频率较低时,其子代中雌性数量较多(Dreissetal.,2006)。在鸟类中,鸣唱和羽色等第二性征是雌性选择配偶的重要指标,可能也是调节后代性比的关键因素(Dreissetal.,2006)。例如,雄性白领姬鹟Ficedulaalbicollis前额白斑越大,其子代中雄性数量越多(Ellegrenetal.,1996),这主要是因为雄性前额白斑的面积可以显示其身体状况(Gustafssonetal.,1995),并且雌性对这种性状的偏爱也具有遗传性(Sheldonetal.,1999)。在对蓝山雀Paruscaeruleus的研究中发现,雄性的鸣唱是吸引雌性的重要信号,其后代性比与雄性鸣唱语句长度存在显著相关性(Dreissetal.,2006)。然而,在歌带鹀Melospizamelodia的研究中却未发现后代性比与鸣唱特征存在显著相关性。鉴于后代性比与鸣唱相关性的研究结论不一致,不同物种(即使同一物种的不同种群)都会存在差异,因此对杂色山雀后代性比和鸣唱关系的研究是很有必要的。
杂色山雀体型较小,全长约13 cm,是一种小型森林洞巢鸟类(Yamaguchi & Higuchi,2005)。本研究选择杂色山雀作为研究物种,主要考虑以下几个原因:首先,杂色山雀鸣唱的研究相对匮乏;其次,杂色山雀在繁殖期鸣声丰富多变,最近有研究指出其多变的鸣声使其有可能成为研究鸟类鸣唱发展的模式系统(李剑等,2013)。同时Andersson(1994)认为鸟类鸣声是最能反映雄性身体状况的生理指标,也是最有效的性吸引指标(Cockburnetal.,2002)。本研究以杂色山雀为研究对象,探索鸣唱与体型、子代性别比(雄性子代数/总子代数)的相关性,为小型雀形目鸟类鸣声行为学研究提供更多数据参考。
1 研究地概况和实验方法
1.1 研究地概况
本研究在辽宁省仙人洞国家级自然保护区内开展,保护区位于辽宁省庄河市境内,地理坐标为122°53′24″~123°03′30″E,39°54′00″~40°03′00″N,总面积达35.747 km2,海拔200~700 m,属温带季风性气候,植被类型在辽东地区具有极好的代表性。杂色山雀是当地留鸟,繁殖期为3—7月。在保护区设置人工巢箱用于繁殖招引研究对象。
1.2 数据收集
声音采集时间为2014—2015年4—6月。录音前期对研究区内的杂色山雀进行彩环标记,血样采集采取非伤害性取样法,使用昆虫针扎破鸟翼尖血管,待有少量血流出时,用已编号的无菌滤纸按住伤口,待伤口停止流血取下滤纸。将带有少量鸟血的滤纸放入自封袋以备提取基因组DNA,使用分子生物学方法(霍亚鹏等,2009)进行性别鉴定,游标卡尺测量体型参数,精确到小数点后2位,体型参数主要包括:体质量、翅长、尾长、跗蹠长度,采样和测量工作在5~10 min内完成。
在不干扰鸟类的前提下对20只雄性杂色山雀进行鸣声录制,录制距离5~10 m,有效录音时长200 h,每只杂色山雀有效录音时长以录制其所有曲目为宜(鲁思凡等,2014),录音时长至少500 s。录音器材为TASCAM HD-P2高保真录音机(Tascam,日本)和Sennheiser MKH416 P48强指向话筒(Sennheiser,德国)。录音时采样频率为44.1 kHz,采样大小为16 bit。录音前通过Panda 8×42双筒望远镜确认鸟类个体并记录彩环颜色。利用Avisoft-SAS Lab Pro(德国)生成语图和测定各项鸣唱参数(图1)。测定参数包括主峰频率、最高频率、最低频率、语句长度。
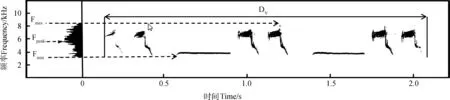
图1 杂色山雀鸣唱语图
Fmax. 最高频率, Fmin. 最低频率, Fpeak. 主峰频率, DV. 语句长度。
Fmax. maximum frequency, Fmin. minimum frequency, Fpeak. peak frequency, DV. duration of verse.
1.3 数据分析
对杂色山雀体质量、翅长、尾长和跗蹠长度使用主成分分析获得体型综合参数,其中第一主成分解释原始变量的34.9%(表1),可以作为体型综合指标(鲁思凡等,2014)。对体质量、跗蹠长度和体型综合参数与杂色山雀鸣唱声音参数进行相关性分析;对杂色山雀声音参数与子代性别比进行相关性分析。统计性显著水平设为0.05。所有数据使用Excel和SPSS 17.0分析。

表1 杂色山雀体型参数的主成分分析Table 1 Principal components analysis of Parus varius variusbody measurements
2 结果
杂色山雀体型参数:体质量、翅长、尾长和跗蹠长度的变异系数相近(CV:4.00%~6.00%;表2);鸣唱语句长度变异系数最大(CV=7.00%;表2)。相关性分析的结果显示,杂色山雀鸣唱的最高频率与体型综合参数具有显著相关性(r=0.645,P=0.019;表3),而其他鸣唱参数与体型综合参数的差异均无统计学意义(P>0.05;表3)。同时,跗蹠长度、体质量与鸣唱各参数不存在显著相关性(P>0.05;表3)。
根据杂色山雀子代性别比和鸣唱参数的相关性分析,发现杂色山雀子代性别比与语句长度具有显著相关性(r=0.696,P=0.025;表3),而子代性别比与其他鸣唱参数均不存在显著相关性(P>0.05;表3)。

表2 杂色山雀的鸣唱特征和体型参数(n=20)Table 2 Descriptive statistics of acoustic parameters and body size of Parus varius varius (n=20)

表3 杂色山雀鸣唱特征与体型参数的相关性Table 3 Correlations of acoustic parameters with body size of Parus varius varius
3 讨论
本研究结果显示杂色山雀鸣唱的最高频率与体型综合参数具有显著相关性,体型越大,鸣唱的最高频率越高。鸣唱是反映鸣鸟身体状态的有效信号,鸣唱的最高频率对食物争夺和领地占据具有较大影响(Zollinger & Brumm,2015),最高频率越高,鸣鸟就可能会占据更好的生存环境,优越的生存环境是保证鸣鸟良好身体状态的重要因素(Krebsetal.,1978)。体型综合参数通常可以反映鸟类个体身体状态,较大的体型意味着较好的身体状态(Zollinger & Brumm,2015)。因此,鸣唱的最高频率与体型具有显著相关性,这与已有研究吻合。本研究发现杂色山雀鸣唱的最高频率与体质量、跗蹠长度不存在显著相关性,说明影响杂色山雀鸣唱的最高频率的因素是体型综合参数而不是单一体型参数(体质量、跗蹠长度),可能是由于体型综合参数代表个体综合身体状态。对于繁殖期鸟类而言,身体状态是繁殖成功的重要条件,因此导致体型综合参数与鸣唱相关性显著(Linhart & Fuchs,2015)。
杂色山雀鸣唱与单一体型参数(体质量、跗蹠长度)不存在显著相关性,可能与体型变异系数和自身调节有关。Patel等(2010)发现鸟类鸣唱参数与体型参数变异系数具有相关性,体型参数变异系数越大,体型与鸣唱相关性越大,例如,黑天鹅Cygnusatratus体质量变异系数约为13%,其体质量与鸣唱具有显著相关性。本研究发现鸣唱各参数分别与体型单一参数(体质量、跗蹠长度)不存在显著相关性,可能是由于杂色山雀的体型变异系数较小。鸣唱与体型相关性不仅与变异系数相关,鸟类也可以通过调节发声器官来调节声音频率以适应环境。在繁殖季,雌鸟偏爱鸣唱频率较高的雄鸟(Cardoso,2011),雄鸟通过调节发声器官以提高声音频率,获得竞争优势,这可能也会影响鸣唱与体型之间的相关性。
在某些物种中,部分体型参数(体质量、跗蹠长度)与鸣唱相关性的强弱可能还受生存环境的影响(Fischeretal.,2004)。生存在同一集群中,体型与声音频率相关性的减弱是很有必要的,尤其是在繁殖期,主要是由于体型与鸣唱之间是否相关对于能否成功繁殖具有十分重要的作用(Eyetal.,2007)。例如,在雌鸟和雄鸟齐聚的区域,体质量与声音频率具有显著相关性,但在单一性别的集群中,这种相关性却不显著(Logueetal.,2007)。因此,本研究中发现杂色山雀鸣唱与部分体型参数相关性不显著可能是由多方面原因导致的。
鸣唱语句长度是衡量雄性身体状态的一个重要指标(Carew,2000)。本研究未发现鸣唱语句长度与杂色山雀体型之间存在显著相关性,然而,鸣唱语句长度与子代性别比存在显著相关性。当雄性具有较长的鸣唱语句时,亲本会有意识地调节子代性别比,导致雄性子代数量增加,可能是由于大部分雀形目鸟类只有雄性子代可以遗传亲本鸣唱语句长度的性状(Sheldonetal.,1999),这些鸟类希望通过调节子代性别比获得更大生殖回报。对于鸣唱语句长度影响子代性别比的生理机制目前尚不清楚。Emlen(1997)认为雌性亲本可以选择适合的卵细胞,或者消除不合适的胚胎,但是具体的机制有待深入研究。本研究结果显示,杂色山雀雄鸟鸣唱语句长度可能与子代性别比有密切的联系,但其他声音参数与子代性别比却未发现显著相关性。
本研究结果发现:杂色山雀体型综合参数与鸣唱的最高频率具有显著相关性,说明体型综合参数是影响杂色山雀鸣唱的最高频率的重要因素,并非单一因素影响杂色山雀鸣唱最高频率;雄性鸣唱语句长度与子代性别比具有显著相关性,说明鸣唱语句长度是影响子代性别比的一个重要因素。本研究丰富了小型鸟类体型与鸣唱的关系,以及鸣唱与子代性别比之间的关系。
霍亚鹏, 曹军, 张宇婷, 等. 2009. 杂色山雀性别分子鉴定方法的研究[J]. 辽宁林业科技, (1): 3-7.
李剑, 万冬梅, 李东来, 等. 2013. 杂色山雀指名亚种繁殖期的鸣声行为[J]. 动物学杂志, 48(4): 513-520.
鲁思凡, 刘金, 夏灿玮. 2014. 体征对灰眉岩鹀鸣唱频率特征的影响[J]. 动物学杂志, 49(3): 334-340.
Andersson MB. 1994. Sexual selection[M]. New Jersey: PrincetonUniversity Press: 176.
Appleby BM, Redpath SM. 1997. Indicators of male quality in the hoots of tawny owls (Strixaluco)[J]. Journal of Raptor Research, 31(1): 65-70.
Araya-Ajoy Y, Chaves-Campos J, Kalko EKV,etal. 2009. High-pitched notes during vocal contests signal genetic diversity in ocellated antbirds[J]. PLoS ONE, 4(12): e8137. DOI:10.1371/journal.pone.0008137.
Beecher MD, Brenowitz EA. 2005. Functional aspects of song learning in songbirds[J]. Trends in Ecology & Evolution, 20(3): 143-149.
Budka M, Osiejuk TS. 2013. Formant frequencies are acoustic cues to caller discrimination and are a weak indicator of the body size of corncrake males[J]. Ethology, 119(11): 960-969.
Cardoso GC. 2011. Paradoxical calls: the opposite signaling role of sound frequency across bird species[J]. Behavioral Ecology, 23(2): 237-241.Carew TJ. 2000. Behavioral neurobiology: the cellular organization of natural behavior[M]. Sunderland: Sinauer Associates Publishers: 435.
Cockburn A, Legge S, Double MC. 2002. Sex ratios: concepts and research methods[J]. American Journal of Human Biology, 15(2): 236-237.
Cynx J, Bean NJ, Rossman I. 2005. Testosterone implants alter the frequency range of zebra finch songs[J]. Hormones and Behavior, 47(4): 446-451.
de Kort SR, Eldermire ER, Cramer ER,etal. 2009. The deterrent effect of bird song in territory defense[J]. Behavioral Ecology, 20(1): 200-206.
Dreiss A, Richard M, Moyen F,etal. 2006. Sex ratio and male sexual characters in a population of blue tits,Paruscaeruleus[J]. Behavioral Ecology, 17(1): 13-19.
Ellegren H, Gustafsson L, Sheldon BC. 1996. Sex ratio adjustment in relation to paternal attractiveness in a wild bird population[J]. Proceedings of the National Academy of Sciences of the United States of America, 93(21): 11723-11728.
Emlen ST. 1997. When mothers prefer daughters over sons[J]. Trends in Ecology & Evolution, 12(8): 291-292.
Evans S, Neave N, Wakelin D. 2006. Relationships between vocal characteristics and body size and shape in human males: an evolutionary explanation for a deep male voice[J]. Biological Psychology, 72(2): 160-163.
Ey E, Pfefferle D, Fischer J. 2007. Do age and sex-related variations reliably reflect body size in non-human primate vocalizations? A review[J]. Primates, 48(4): 253-267.
Fischer J, Kitchen DM, Seyfarth RM,etal. 2004. Baboon loud calls advertise male quality: acoustic features and their relation to rank, age, and exhaustion[J]. Behavioral Ecology and Sociobiology, 56(2): 140-148.
Fletcher NH. 2004. A simple frequency-scaling rule for animal communication[J]. The Journal of the Acoustical Society of America, 115(5): 2334-2338.
Gustafsson L, Qvarnström A, Sheldon BC. 1995. Trade-offs between life-history traits and a secondary sexual character in male collared flycatchers[J]. Nature, 375(6529): 311-313.
Krebs J, Ashcroft R, Webber M. 1978. Song repertoires and territory defence in the great tit[J]. Nature, 271(5645): 539-542 .
Linhart P, Fuchs R. 2015. Song pitch indicates body size and correlates with males’ response to playback in a songbird[J]. Animal Behaviour, 103: 91-98.
Lislevand T, Figuerola J, Székely T. 2007. Avian body sizes in relation to fecundity, mating system, display behavior, and resource[J]. Ecology, 88(6): 1605.
Logue DM, Droessler EE, Roscoe DW,etal. 2007. Sexually antithetical song structure in a duet singing wren[J]. Behaviour, 144(3): 331-350.
Mager JN, Walcott C, Piper WH. 2007. Male common loons,Gaviaimmer, communicate body mass and condition through dominant frequencies of territorial yodels[J]. Animal Behaviour, 73(4): 683-690.
Morton ES. 2000. An evolutionary view of the origins and functions of avian vocal communication[J]. Japanese Journal of Ornithology, 49(2): 69-78.
Nager RG, Monaghan P, Griffiths R,etal. 1999. Experimental demonstration that offspring sex ratio varies with maternal condition[J]. Proceedings of the National Academy of Sciences of the United States of America, 96(2): 570-573.
Osiejuk TS, Ratyńska K, Cygan JP,etal. 2005. Frequency shift in homologue syllables of the ortolan buntingEmberizahortulana[J]. Behavioural Processes, 68(1): 69-83.
Patel R, Mulder RA, Cardoso GC. 2010. What makes vocalisation frequency an unreliable signal of body size in birds? A study on black swans[J]. Ethology, 116(6): 554-563.
Peters G, Baum L, Peters MK,etal. 2009. Spectral characteristics of intense mew calls in cat species of the genusFelis(Mammalia: Carnivora: Felidae)[J]. Journal of Ethology, 27(2): 221-237.
Potvin DA. 2013. Larger body size on islands affects silvereyeZosteropslateralissong and call frequency[J]. Journal of Avian Biology, 44(3) : 221-225.
Searcy WA, Anderson RC, Nowicki S. 2006. Bird song as a signal of aggressive intent[J]. Behavioral Ecology and Sociobiology, 60(2): 234-241.Sheldon BC, Andersson S, Griffith SC,etal. 1999. Ultraviolet colour variation influences blue tit sex ratios[J]. Nature, 402(6764): 874-877.
Sheldon BC. 1999. Sex allocation: at the females’ whim[J]. Current Biology, 9(13): R487-R489.
Staicer CA, Spector DA, Horn AG. 1996. Ecology and evolution of acoustic communication in birds[M]. New York: Cornell University Press: 426-453.
Taylor AM, Reby D. 2010. The contribution of sourceefilter theory to mammal vocal communication research[J]. Journal of Zoology, 280(3): 221-236.
Trivers RL, Willard DE. 1973. Natural selection of parental ability to vary the sex ratio of offspring[J]. Science, 179(4068): 90-92.
Weeden JS, Falls JB. 1959. Differential responses of male ovenbirds to recorded songs of neighboring and more distant individuals[J]. The Auk, 76(3): 343-351.
Yamaguchi N, Higuchi H. 2005. Extremely low nesting success and characteristics of life history traits in an insular population ofParusvariusnamiyei[J]. The Wilson Bulletin, 117(2): 189-193.
Zollinger SA, Brumm H. 2015. Why birds sing loud songs and why they sometimes don’t[J]. Animal Behaviour, 105: 289-295.
The Correlation Research of the Relationship between Body Size,Sex Ratio of Offspring and Bird Song inParusvariusvarius
ZHAOTingting1,LIUYan1,ZHANGZhiwei1,HUOYapeng1,LINJingfeng2,WANDongmei1,YINJiangxia1*
(1. Key Laboratory of Animal Resource and Epidemic Disease Prevention, Department of Life Sciences, Liaoning University,Shenyang 110036, China; 2. Xianrendong Nature Reserve Administration, Dalian, Liaoning Province 116400, China)
Bird song is an important behavior in birds, with great evolutionary and ecological implications. It is one of the best effective ways for communication among individuals within the bird communities. It has been suggested that there is a significant relationship between bird song and individual intrinsic factors (body size, sex ratio of offspring). To investigate the relationship between the singing behavior of varied tits (Parusvariusvarius) and body size as well as sex ratio of offspring, we recorded the songs ofP.variusvariusbetween March and June in 2014-2015 at Xianrendong Nature Reserve, Liaoning province. Additionally, the body size and sex ratio of offspring ofP.variusvariuswere also measured. The results showed that: (1) there was a positive relationship between integrated morphological measurement and maximum frequency of bird song; (2) there was a positive relationship between sex ratio of offspring and duration of verse of bird song. This study enriched the small birds’ body size relationship with the song, and the relationship between the song and sex ratio of offspring.
Parusvariusvarius; bird song; body size; sex ratio of offspring
2016-07-17 接受日期:2016-10-31
国家自然科学基金项目(31100271)
赵婷婷(1991—), 女, 硕士, 研究方向为鸟类学, E-mail:1074878569@qq.com
*通信作者Corresponding author, 副教授, E-mail:xia0615@126.com
10.11984/j.issn.1000-7083.20160190
Q959.7
A
1000-7083(2017)02-0168-06
