芝麻种质资源成株期抗旱性关联分析
2017-04-08刘文萍吕伟黎冬华任果香张艳欣文飞韩俊梅张秀荣
刘文萍,吕伟,黎冬华,任果香,张艳欣,文飞,韩俊梅,张秀荣
(1山西省农业科学院经济作物研究所,太原 030031;2中国农业科学院油料作物研究所,武汉 430062)
芝麻种质资源成株期抗旱性关联分析
刘文萍1,吕伟1,黎冬华2,任果香1,张艳欣2,文飞1,韩俊梅1,张秀荣2
(1山西省农业科学院经济作物研究所,太原 030031;2中国农业科学院油料作物研究所,武汉 430062)
【目的】利用33个多态性SSR分子标记分别与18个不同的抗旱性状进行关联分析,发掘与抗旱相关的主要基因位点,为抗旱基因定位和功能标记开发提供基础;通过对100份芝麻种质资源进行抗旱性鉴定,发掘优异的耐旱种质,为芝麻抗旱育种提供指导。【方法】采用盆栽和反复干旱法,对芝麻种质资源群体进行成株期抗旱性鉴定获得表型指标测定值,利用SAS、SPSS和隶属函数等进行统计分析,综合评价其抗旱性,利用GLM模型和MLM模型,将表型数据与分子标记进行关联分析。【结果】研究群体干旱胁迫处理后,材料间响应差异明显,考察的表型性状测定值均小于对照;干旱胁迫条件下18个性状值的变异系数平均为0.31,高于对照(平均为0.19);处理与对照间各性状指标经配对t检验,均达极显著水平;通过连续变数的次数分布统计方法、主成分分析和隶属函数分析,筛选出10个与抗旱性响应关系密切的指标,并筛选出12份高抗旱种质;基于芝麻基因组筛选出的33个多态性SSR标记扫描供试材料,共检测到170个等位变异,平均每个标记5.15个;利用structure数学模型对供试群体进行遗传结构分析,可分为2个亚群;利用GLM模型和MLM模型分别检测到120个和63个标记位点与供试群体抗旱系数显著关联(P<0.05),表型变异解释率分别为3.85%—14.30%和4.00%—12.5%,解释率大于10%的标记位点分别有12个和3个,其中,位点4033-3和4033-2均与第一主成分因子第2次复水前萎蔫叶片数显著关联,且变异解释率均为最高,分别达14.3%和12.5%,2个模型共同检测到的标记位点有5个。通过引物序列在基因组上的位置比对,发现3个可能存在芝麻抗旱相关基因的基因组区段。【结论】利用综合评价方法,筛选出柳林芝麻3号、g80、8602-2等12份高抗旱芝麻种质,同时利用GLM和MLM 2个模型检测到与第2次复水前萎蔫叶片数显著关联的标记位点(位点4033-3和位点4033-2),且变异解释率最高,分别达14.3%和12.5%。
芝麻种质资源;抗旱性;SSR标记;关联分析
0 引言
【研究意义】芝麻隶属胡麻科,一年生草本植物,是中国主要的油料作物之一,在中国有着悠久的种植历史,多数省份皆有种植[1]。山西地处干旱半干旱地区,降水量少,干旱严重,导致芝麻出苗率低、生长缓慢、产量降低,成为限制芝麻增产的主要限制因子,因此,研究芝麻的抗旱性,挖掘抗旱芝麻种质资源,同时,利用SSR分子标记与抗旱性状进行关联分析,发掘与抗旱相关的主要基因位点,开发抗旱分子标记,并将其应用到芝麻育种中,对提高芝麻的抗旱性和产量水平具有很重要意义。【前人研究进展】国内外已对玉米、水稻、大豆、小麦、花生、油菜[2-7]等作物进行了较系统的抗旱性研究。早在20世纪70年代末,CARTER等[8]通过植株萎蔫程度对300余份大豆种质进行了抗旱性鉴定;SONGSRI等[9]、刘吉利等[10]、张智猛等[11]对花生的生长发育和生理生态等方面的干旱适应机制进行了研究;NOROUZI等[12]对甘蓝型油菜研究表明,干旱胁迫下叶片含水量[13]、萎蔫指数[14]、种子活力指数[7]、相对根体积[15]以及脯氨酸[16]等生理指标发生明显变化,千粒重、生物量等下降[17],可作为油菜抗旱性鉴定的指标。在芝麻抗旱性研究方面国内外也有一些报道,MENSAH等[18]利用盆栽法对芝麻苗期进行间隔不同天数浇水处理研究,结果表明,
水分不足会直接影响芝麻植株的生长,株高、叶面积、地下部干重和地上部干重等降低,导致单株产量显著下降;孙建等[19]利用盆栽试验对芝麻苗期进行干旱胁迫研究,结果表明,芝麻苗期进行干旱处理对千粒重和产量的影响较大。【本研究切入点】作物的抗旱性属于较复杂的数量性状,受多基因控制[20],关联分析是研究复杂数量性状的有效方法[21],中国芝麻抗旱性分子基础研究报道较少,仅见黎冬华等[22]采用不同浓度PEG 6000胁迫发芽芝麻种子基于SSR、SRAP和AFLP分子标记的关联分析研究报道,而国外对芝麻抗旱性分子基础研究还尚未见报道。【拟解决的关键问题】本研究通过对芝麻自然研究群体进行抗旱表型鉴定,获得抗旱相关表型数据,并与全基因组分子标记数据进行关联分析,发掘与抗旱相关性状基因位点,筛选得到抗旱种质资源,为芝麻抗旱遗传改良提供理论基础。
1 材料与方法
1.1 试验材料
选用来源于山西和陕西芝麻种质资源共计100份,由国家芝麻种质资源中期库(中国农业科学院油料作物研究所)提供,并进行了4代纯化。
1.2 干旱处理
试验于2014年5月至10月在山西省农业科学院经济作物研究所人工遮雨棚内进行,采用盆栽法[23],每个材料种植6盆,其中3盆用于干旱处理,3盆为对照,共计600盆。盆钵口径35 cm,盆栽土用芬兰进口营养土与蛭石按1∶1等量混合,每盆装3 kg,每盆浇等量的水1 L,待适墒时播种。出苗后间苗2次,每盆保留均匀生长的苗5株,在干旱处理前保持正常供水。在芝麻4对真叶展开时进行干旱处理,采用反复干旱法,处理3次,每次干旱处理后待50%左右的材料出现永久萎蔫时进行复水,之后全部材料按正常管理浇水直至成熟。
1.3 表型性状调查
处理期调查:在干旱处理的每次复水前同一时间段内(3次复水前均在上午9:00以前)调查每株总叶片数(全展叶)、萎焉叶片数(全展叶);每次复水后第2天早上调查每株总叶片数(全展叶)、萎焉叶片数(全展叶)。成熟期调查:调查每个材料处理和对照每株的株高、始蒴高度、单株蒴果数、单株产量、千粒重、单株地上部分生物量、地下部分生物量,计算平均值。
1.4 抗旱能力的综合评价
各指标抗旱系数Xj=处理测定值/对照测定值;负向指标的抗旱系数Xj=1-处理测定值/对照测定值(1)
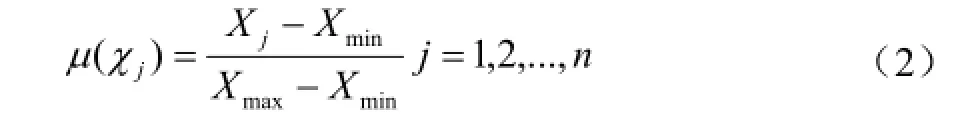
每一个材料各综合指标的隶属函数值由公式(2)求得[24-25],式中,Xj表示第j个综合指标,Xmax和Xmin分别表示第j个综合指标的最大值和最小值。
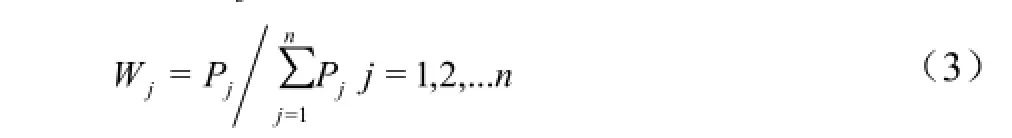
由公式(3)计算各综合指标的权重[24-25],式中,Wj表示第j个综合指标在所有综合指标中的重要程度;Pj为各材料第j个综合指标的贡献率。
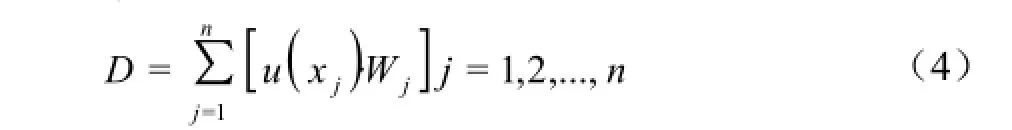
由公式(4)计算各材料的综合抗旱能力(D值)。
用SPSS和SAS 9.1软件进行相关性分析、主成分分析。
1.5 基因组DNA的提取和SSR分子标记分析
每份材料取苗期嫩叶的混合样,采用CTAB法[26]提取基因组DNA,经紫外分光光度计法检测其质量和浓度,稀释至20 ng·μL-1,置-20℃冰箱保存。PCR体系及扩增程序参照文献[27]。PCR扩增产物经6%聚丙烯酰胺凝胶电泳,银染显色后照相。
1.6 关联分析方法数据统计分析
以Structure 2.3.1软件进行群体遗传结构分析,估计最佳群体组群数K,其取值范围为1—10,将MCMC(Markov Chain Monte Carlo)开始时的不作数迭代(length of burn-in period)设为100 000次,再将不作数迭代后的MCMC设为100 000次,迭代次数(number of iterations)设置为5,依据似然值最大原则,选取合适的K值为群体数目,并绘出基于模型的群体遗传结构图。采用Tassel 2.1 软件GLM(general linear model)和MLM(mixed linear model)2种模型程序,进行分子标记数据和表型数据的关联分析。
2 结果
2.1 供试材料对干旱胁迫的响应分析
对供试材料考察了18个性状,包括总叶片数、萎焉叶片数、生物量、单株蒴果数、单株产量、株高等,处理与对照间存在明显差异,各项指标平均值均小于对照,表明各个材料均受到干旱胁迫的影响。干旱胁迫下,18个性状值的变异系数在0.07—0.65,平均为0.31,对照的变异系数在0.06—0.40,平均为0.19,除株高外,其他性状干旱胁迫的变异系数均大于对照。处理与对照间各性状指标经配对t检验,均为极显著,表明干旱处理对各性状产生了明显的抑制作用(表1)。处理组与对照组相比18个性状的平均抗旱系数在0.326—0.951,均小于1.000,也表明测定的各项指标均受到干旱胁迫不同程度的影响(表2)。以上结果表明,研究材料间和调查的18个性状均受到干旱胁迫,对干旱胁迫的响应存在明显差异。
2.2 各项指标抗旱性综合评价
参照连续变数的次数分布统计方法,将供试材料18个性状的抗旱系数以组距为0.2分成7个组区间,制作成次数分布表(表3)。结果表明,在同一组区间各性状的抗旱系数分布次数相差较大,其中,第1次复水前总叶片数、第2次复水前总叶片数、千粒重这三个指标的抗旱系数在0.9<Xj≤1.1区间的分布频率分别为82%、50%和53%,表明这三个性状对干旱胁迫反应迟钝;而第1次复水前萎蔫叶片数、第2次复水前萎蔫叶片数、第2次复水后萎蔫叶片数、第3次复水前总叶片数、第3次复水前萎蔫叶片数、第3次复水后萎蔫叶片数、地上部鲜重、地上部干重、成熟期株高、单株产量这10个指标的抗旱系数在Xj<0.9区间分布频率分别为92%、77%、67%、86%、100%、83%、62%、60%、68%和62%,表明这10个性状对干旱胁迫的反应较敏感。由抗旱系数相关系数矩阵可看出(表4),各单项指标间存在一定的相关性,说明芝麻抗旱性是一个复杂性状。
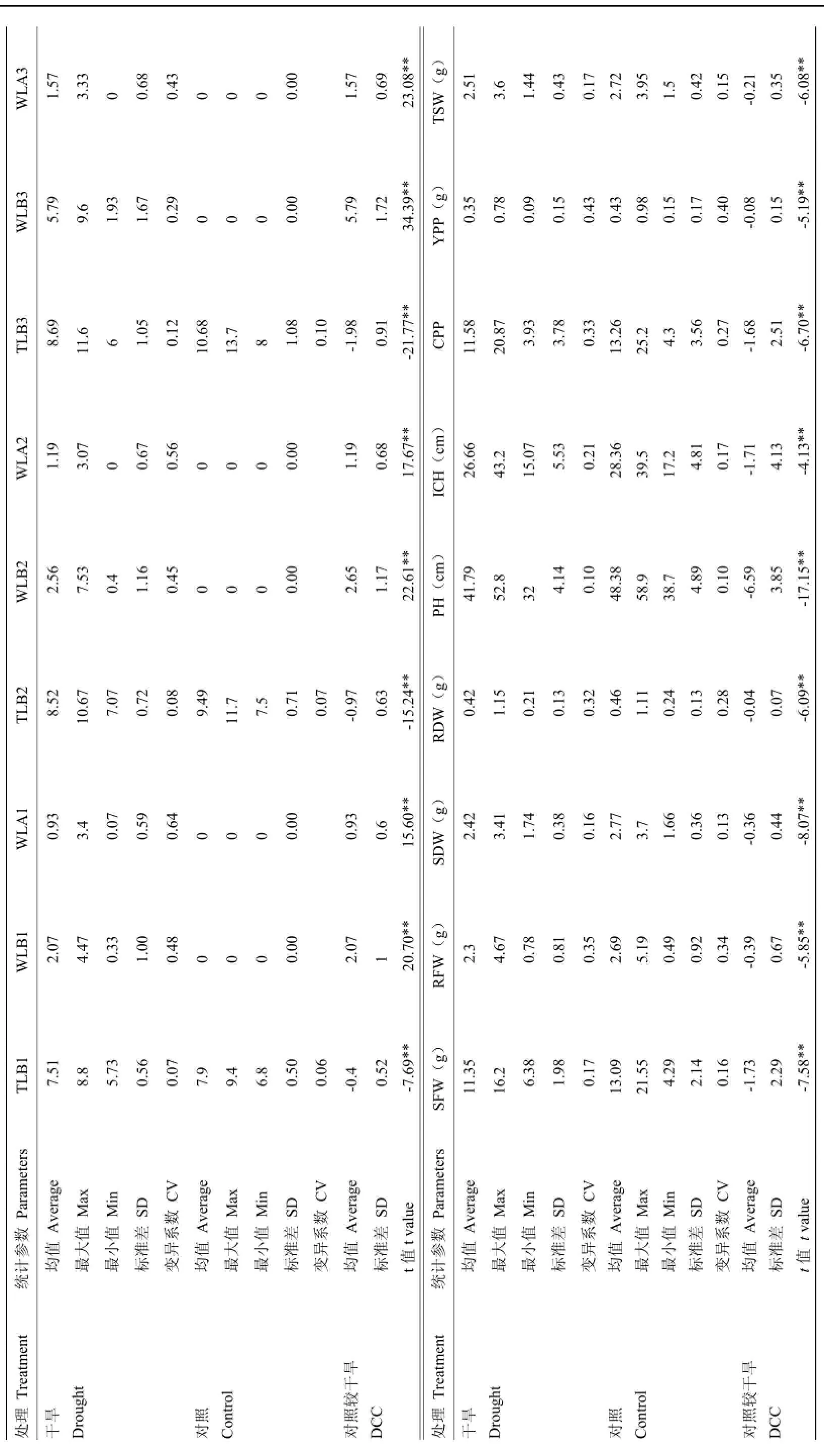
表1 各性测定值的差异分析Table 1 Variance analysis of mred value between characters

表2 各单指标的抗旱系数Table 2 Drought resistance coefficients of single index

续表2 Continued table 2

续表2 Continued table 2

续表2 Continued table 2
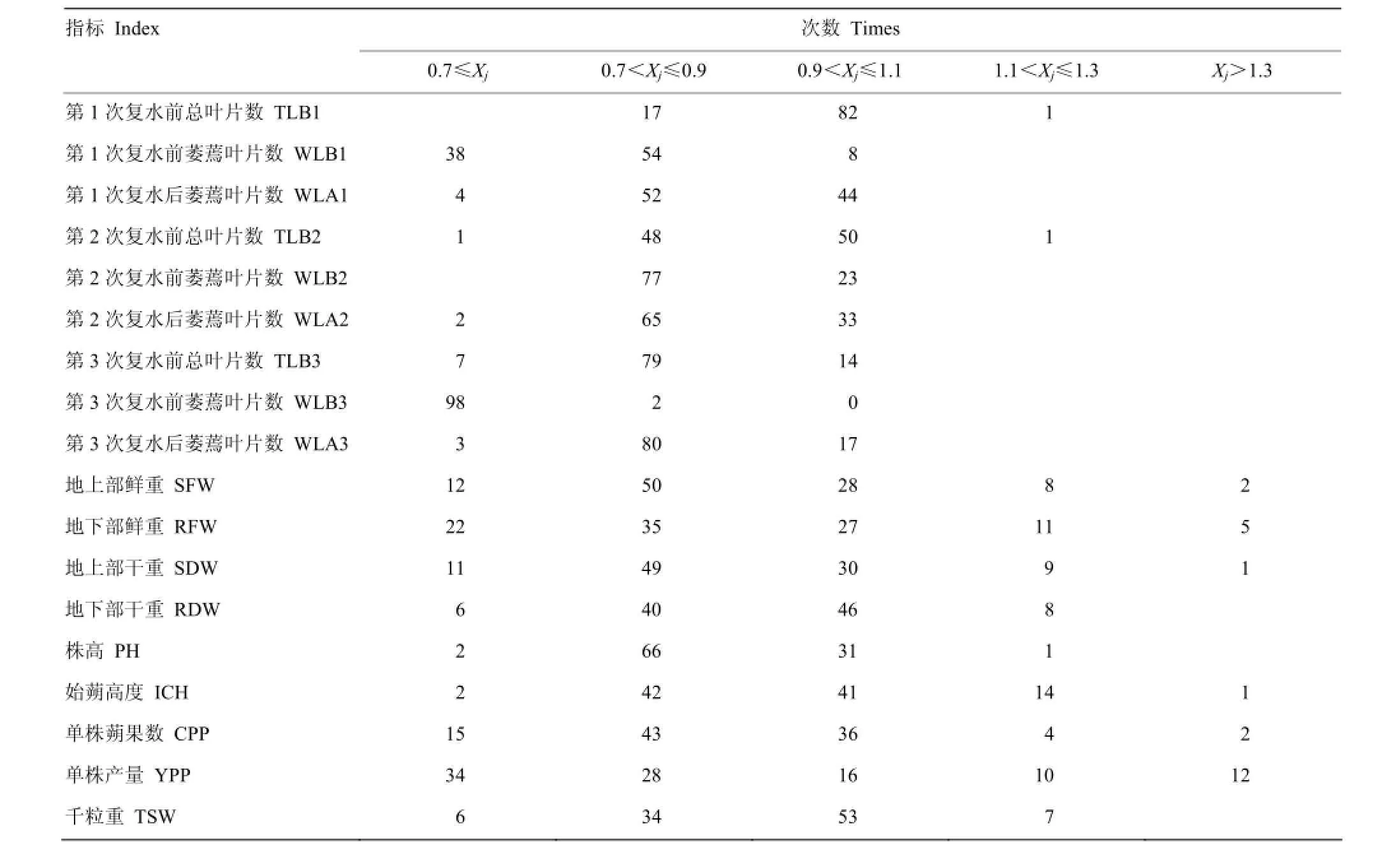
表3 供试材料各性状指标的抗旱系数在不同区间的分布Table 3 Different distributions of drought-resistance coefficients of tested materials traits indexes
对干旱较敏感的10个性状抗旱系数进行主成分分析,结果表明,前5个因子的累积贡献率达到82%,因而取前5个因子,将以上10个性状转换为5个新的独立的综合指标,分别用CI(1)、CI(2)、CI(3)、CI(4)、CI(5)表示。第一主成分中第2次复水前萎蔫叶片数、第3次复水后萎蔫叶片数有较强的载荷量;第二主成分中地上部干重有较强的载荷量;第三主成分中,第1次复水前萎蔫叶片数、第3次复水前总叶片数、单株产量有较强的载荷量;第四主成分中,地上部鲜重、株高有较强的载荷量;第五主成分中,第2次复水后萎蔫叶片数、第3次复水前萎蔫叶片数有较强的载荷量(表5)。
根据公式(2)、(3)、(4)求得供试材料抗旱性综合评价D值(表2)。根据D值大小对供试材料进行抗旱性排序,筛选出12份高抗旱芝麻种质资源(D值>0.65),占供试材料的12%,其中,包括柳林芝麻3号、吴堡县岔上乡步墕村芝麻、g80、8602-2、柳林芝麻1号、临县芝麻2号、临县芝麻5号、g62、g45、吴堡县岔上乡郭家墕村芝麻、2012-48-01、四棱芝麻。
2.3 关联分析
基于芝麻基因组开发多态性较好的33个SSR标记在100份芝麻供试材料中共检测到170个等位变异,平均每个标记5.15个。利用structure数学模型对供试群体进行遗传结构分析,由此判断供试群体可被分为2个亚群,并绘制供试材料群体结构图(图1)。
利用GLM模型检测到120个位点与以上对干旱胁迫响应敏感的10个性状显著关联(P<0.05),每个性状的位点数在4—31个,变异解释率为3.85%—14.30%(表6),有28个位点同时与2个或多个性状关联;位点2177-3关联的频次最高,同时与5个指标显著关联,分别为第1次复水前萎蔫叶片数、第2、3次复水前、后萎蔫叶片数,解释率为6.43%—12.76%,其中第2次复水前萎蔫叶片数、第3次复水后萎蔫叶片数均属于第一主成分中的因子;解释率大于10%的标记位点有12个,其中1816-2、1664-3、1550-5、2177-3、1816-4、4033-3等6个位点与2个第一主成分因子(第2次复水前萎蔫叶片数和第3次复水后萎蔫叶片数)显著关联,4033-3是所有位点中变异解释率最高的,对第2次复水前萎蔫叶片数解释率达14.30%。

表4 各指标抗旱系数的相关系数矩阵Table 4 Correlation coefficient matrix between drought resistance indexes
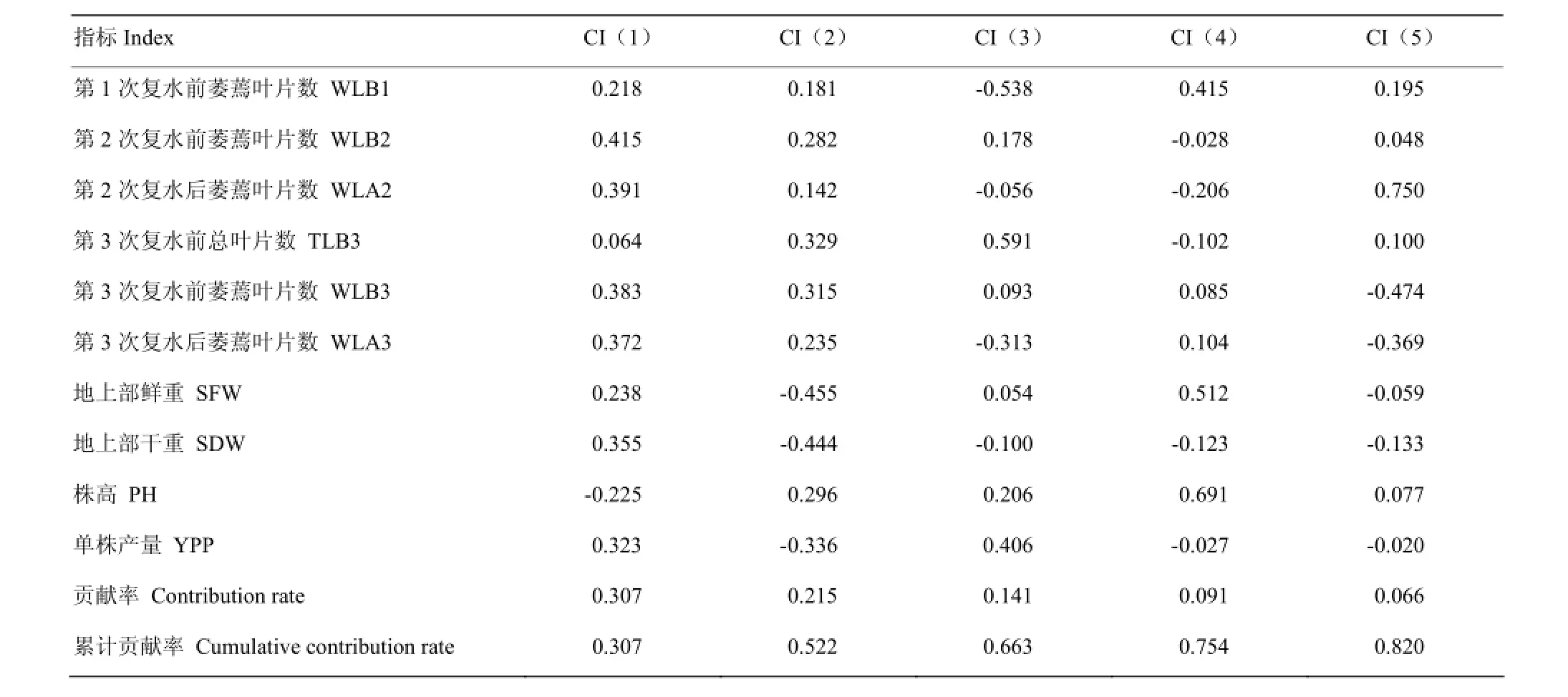
表5 主成分载荷矩阵及贡献率Table 5 Component matrix and contribution rate
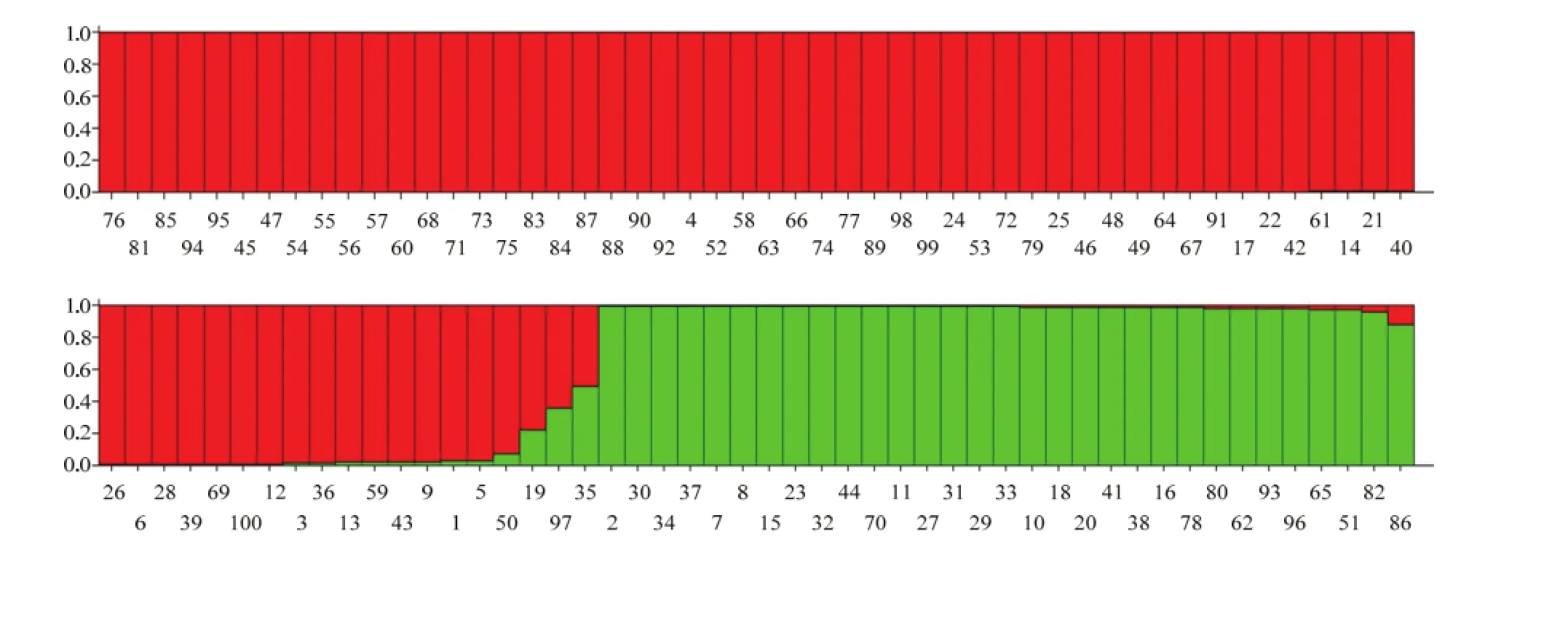
图1 基于SSR标记的100份芝麻种质遗传结构Fig. 1 Population structure of 100 sesame materials based on SSR marker
利用MLM模型检测到63个与10个性状显著关联(P<0.05),每个性状的位点数分别在3—11个,变异解释率为4.00%—12.50%,有13个位点同时与2个或多个性状关联;位点1816-3同时与4个指标显著关联,分别为第1、2次复水前萎蔫叶片数和第2、3次复水后萎蔫叶片数,解释率为4.32%—6.32%,其中第2次复水前萎蔫叶片数、第3次复水后萎蔫叶片数均属于第一主成分中的因子;解释率大于10%的标记位点有3个,分别是2209-7、1935-6和4033-2,其中,变异解释率最高的位点4033-2所关联的性状也为第2次复水前萎蔫叶片数,解释率达12.5%。
利用2种模型均能检测到的标记位点有5个,其中位点4033-6、1664-1同时与第1次复水前萎蔫叶片数显著关联,解释率分别为4.46%—5.88%和4.07%—4.64%,该指标属于第三主成分中的因子;位点2218-1与第2次复水后萎蔫叶片数关联显著,解释率为9.9%—13.66%,该指标属于第五主成分中的因子;位点1550-3同时与地上部鲜重、地上部干重显著关联,解释率分别为5.36%—6.08%和4.09%—7.88%,这两个指标分别属于第四、二主成分中的因子。
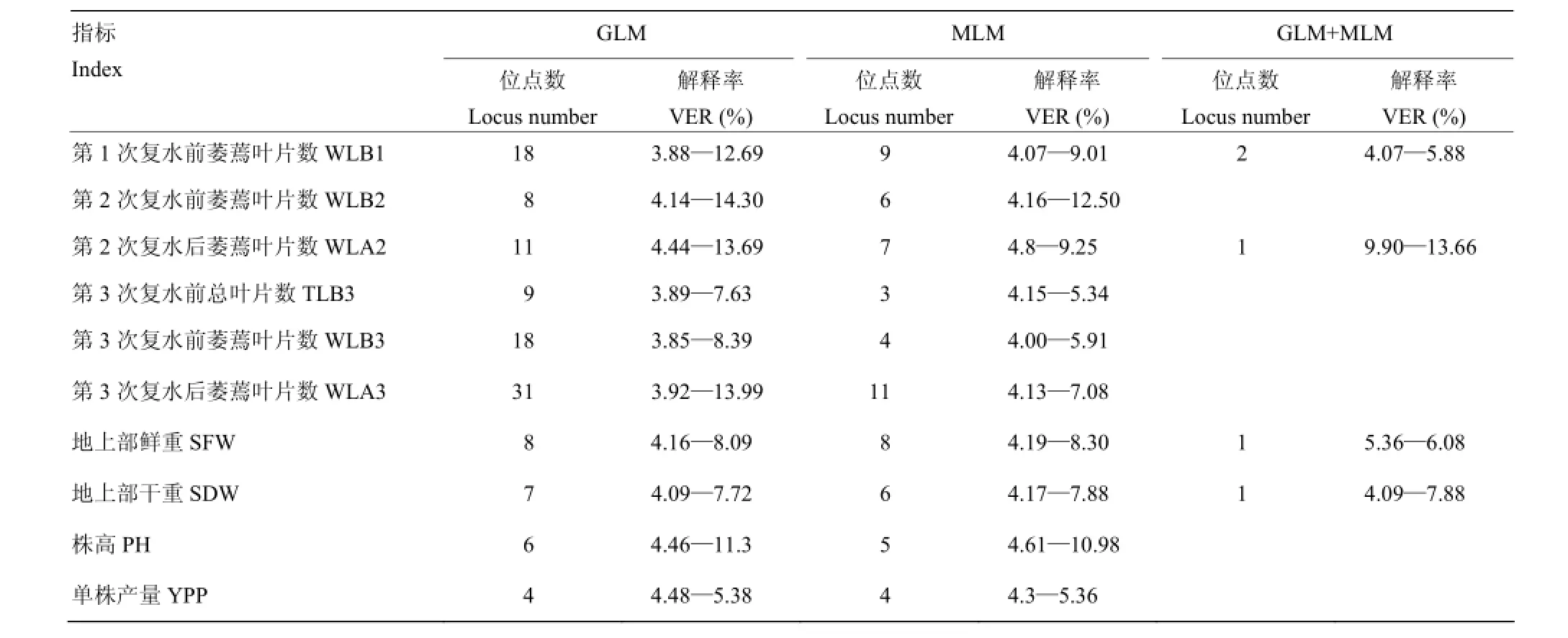
表6 与抗旱性显著相关(P<0.05)的标记位点数和变异解释率Table 6 Number of loci associated with drought resistance(P<0.05)and phenotypic variation explanation rates
对第一主成分因子性状关联到的位点进一步分析,其中贡献率较大且重复率高的标记位点主要有:4033-2和4033-3对第2次复水前萎蔫叶片数变异解释率分别为12.5%和14.3%,2177-3对第2次复水前萎蔫叶片数、第3次复水后萎蔫叶片数变异解释率分别为11.86%和12.21%,1816-2和1826-4对第3次复水后萎蔫叶片数变异解释率分别为10.44%和13.99%,这些标记位点分别来自4033、2177和1816 3个引物,通过比对这三个引物序列在芝麻基因组上的位置,结果表明,4033位于LG4的1 981 965 bp附近,2177位于LG6的11 802 028 bp附近,1816位于LG11的10 438 634 bp附近,推测以上3个基因组区段可能存在芝麻抗旱性相关基因。
3 讨论
近年来,中国国内外对芝麻抗旱性研究多局限于大田干旱胁迫和利用PEG模拟干旱对芝麻种子芽期进行抗旱性鉴定,本研究对芝麻成株期进行反复干旱试验,综合利用与抗旱响应关系密切的多项指标进行芝麻抗旱性综合评价,为芝麻抗旱性精准鉴定提供了方法参考和理论基础。截至目前,国内外对芝麻抗旱性研究的供试材料数量和指标数均少于本研究。KADKHODAIE等[28]评价了10份芝麻品种在不同土壤水分条件下的抗旱性差异;孙建等[19]对5个芝麻品种进行苗期干旱胁迫研究,测定指标均少于10个。本研究对100份芝麻资源测定了18个相关表型性状,其中,总叶片数和萎蔫叶片数属首次采用。采用连续变数的次数分布统计方法对供试材料进行分析发现,千粒重对干旱胁迫反应较迟钝,这与孙建等[19]等的研究结果不一致,而与祁旭升等[25]在胡麻抗旱性研究所得结果相一致;利用综合分析方法得到第2次复水前萎蔫叶片数和第3次复水后萎蔫叶片数2个与抗旱响应最为密切,为芝麻抗旱鉴定提供了可靠的新的指标。
植物的抗旱性是受多因素影响的、且非常复杂的数量性状,不同指标的选择不同,研究得到的结论也不一样。因此,单一指标或少数指标的简单分析很难且可靠地反应不同资源品种的抗旱性。近年来,虽然国内外学者针对不同作物从不同角度研究和提出了更加全面的抗旱评价鉴定方法[29-30],然而现今对抗旱性鉴定评价的指标和方法均没有准确可靠的统一规定,因此,建立科学的抗旱性综合评价方法是芝麻抗旱性鉴定的基础。目前普遍认为多指标多方法相结合的抗旱性综合评价比较可靠[31],朱宗河等[32]对油菜抗旱性的研究表明,利用隶属函数等分析对甘蓝型油菜耐旱性进行综合评价,可筛选甘蓝型油菜耐旱种质。因此,本研究采用综合评价方法,可以较好地揭示各指标与抗旱性的关系,这样既考虑了各指标间的相互关系,又考虑到各指标的重要性。该方法已在大豆[33]、小麦[34]等作物上应用,取得了比较理想的结果。不仅可以缩短芝麻抗旱性鉴定周期,同时为下一步研究耐旱分子机制提供理论依据。
芝麻的抗旱性是复杂的数量性状,关联分析是研究复杂数量性状的有效方法,国内外在芝麻抗旱分子基础研究方面,仅见黎冬华等[22]采用PEG 6000胁迫发芽芝麻种子基于SSR、SRAP和AFLP分子标记的关联分析研究报道,利用GLM模型获得2个解释率(4.965%和4.18%)较高的SRAP标记和1个解释率(4.38%)较高SSR标记。本研究对芝麻成株期进行抗旱鉴定,利用全基因组开发的33个多态性SSR标记与抗旱相关性状进行关联分析,利用GLM和MLM 2种模型分别检测出120个和63个与抗旱性有显著关联(P<0.05)的分子标记位点,变异解释率分别为3.85%—14.30%和4.00%—12.5%,其中,2种模型均检测到与第一主成分因子第2次复水前萎蔫叶片数显著关联的标记位点,且变异解释率均为最高,分别达14.3%(位点4033-3)和12.5%(位点4033-2),通过比对4033引物序列在芝麻基因组上的位置,进一步发现基因SIN_1006195在其附近,该基因属于已报道与抗旱紧密相关的NAC基因家族,可以作为芝麻抗旱后续研究的候选基因之一。研究结果为芝麻抗旱功能分子标记开发和抗旱基因发掘奠定了重要基础。
本研究通过综合评价,还筛选出一批高抗旱种质材料,为抗旱育种研究提供了关键材料。
4 结论
采用抗旱系数进行综合评价,获得柳林芝麻3号、g80、8602-2等高抗旱芝麻种质12份,可用于抗旱性育种;利用GLM和MLM 2个模型均检测到与第一主成分因子第2次复水前萎蔫叶片数显著关联且变异解释率最高的标记位点(位点4033-3和位点4033-2),此标记可应用于分子标记辅助选择。
[1] 黄凤洪. 花生芝麻加工技术. 北京: 金盾出版社, 1995.
HUANG F H.The Processing Technology of Peanut and Sesame. Beijing: Shield Press, 1995. (in Chinese)
[2] NEMETH M, JANDA T, HORVATH E, PALDI E, SZALAI G. Exogenous salicylic acid increases polyamine content but may decrease drought tolerance in maize.Plant Science, 2002, 162: 569-574.
[3] FU B Y, XIONG J H, ZHU L H, ZHAO X Q, XU H X, GAO Y M, LI Y S, XU J L, LI Z K. Identification of functional candidate genes for drought tolerance in rice.Molecular Genetics and Genomics, 2007, 278: 599-609.
[4] 李灿东, 蒋洪蔚, 郭泰, 王志新, 吴秀红, 郑伟, 陈庆山, 胡国华.大豆耐旱选择群体基因型分析与株高QTL定位. 大豆科学, 2011, 30(1): 15-19.
LI C D, JIANG H W, GUO T, WANG Z X, WU X H, ZHENG W, CHEN Q S, HU G H. QTL identification of plant height and analysis of genotype to soybean in selection population.Soybean Science, 2011, 30(1): 15-19. (in Chinese)
[5] PAKNIYAT H, TAVAKOL E. RAPD markers associated with drought tolerance in bread wheat (Triticum aestivumL.).Pakistan Journal of Biological Sciences, 2007, 10: 3237-3239.
[6] 贺鸿雁, 孙存华, 杜伟, 李扬. PEG 6000胁迫对花生幼苗渗透调节物质的影响. 中国油料作物学报, 2006, 28(1): 76-78.
HE H Y, SUN C H, DU W, LI Y. Effects of PEG 6000 osmotic stress on osmolytes of peanut seed-ling.Chinese Journal of Oil Crop Sciences, 2006, 28(1): 76-78. (in Chinese)
[7] 李震, 杨春杰, 张学昆, 邹崇顺, 程勇, 郑普英, 李桂英. PEG胁迫下甘蓝型油菜品种(系)种子发芽耐旱性鉴定. 中国油料作物学报, 2008, 30(4): 438-442.
LI Z, YANG C J, ZHANG X K, ZOU C S, CHENG Y, ZHENG P Y, LI G Y. Evaluation of drought tolerance in rapeseed (Brassica napusL.) during germination under PEG6000 stress.Chinese Journal of Oil Crop Sciences, 2008, 30(4): 438-442. (in Chinese)
[8] CARTER T E JR, RUFTY T W, KUO C G. Soybean plant introduction exhibiting drought and aluminum tolerance. Adaptation of food crops to temperature and water stress. Proceedings of an international symposium, Taiwan, 1993: 335-346.
[9] SONGSRI P, JOGLOY S, KESMALA T, VORASOOT N, AKKASAENG C, PATANOTHAI A. Responses of reproductive characters of drought resistant peanut genotypes to drought.Asian Journal of Plant Sciences, 2008, 7(5): 427-439.
[10] 刘吉利, 赵长星, 吴娜, 王月福, 王铭伦. 苗期干旱及复水对花生光合特性及水分利用效率的影响. 中国农业科学, 2011, 44(3): 469-476.
LIU J L, ZHAO C X, WU N, WANG Y F, WANG M L. Effects of drought and rewatering at seedling stage on photosynthetic characteristics and water use efficiency of peanut.Scientia Agricultura Sinica, 2011, 44(3): 469-476. (in Chinese)
[11] 张智猛, 戴良香, 丁红, 陈殿绪, 杨伟强, 宋文武, 万书波. 中国北方主栽花生品种抗旱性鉴定与评价. 作物学报, 2012, 38(3): 495-504.
ZHANG Z M, DAI L X, DING H, CHEN D X, YANG W Q, SONG W W, WAN S B. Identification and evaluation of drought resistance in different peanut varieties widely grown in Northern China.Acta Agronomica Sinica, 2012, 38(3): 495-504. (in Chinese)
[12] NOROUZI M, TOORCHI M, SALEKDEH G H, MOHAMMADI S A, NISHABOURI M R, AHARIZAD S. Effect of water deficit on growth, grain yield and osmotic adjustment in rapeseed.Journal of Food, Agriculture & Environment, 2008, 6(2): 132-138.
[13] ESLAM B P. Evaluation of physiological indices, yield and its components as screening techniques for water deficit tolerance in oilseed rape cultivars.Journal of Agricultural Science and Technology, 2009, 11: 413-422.
[14] 李震, 吴北京, 陆光远, 程勇, 邹崇顺, 张学昆. 不同基因型油菜对苗期水分胁迫的生理响应. 中国油料作物学报, 2012, 34(1): 33-39.
LI Z, WU B J, LU G Y, CHENG Y, ZOU C S, ZHANG X K. Differences in physiological responses ofBrassica napusgenotypes under water stress during seedling stage.Chinese Journal of Oil Crop Science, 2012, 34(1): 33-39. (in Chinese)
[15] 张霞, 谢小玉. PEG胁迫下甘蓝型油菜种子萌发期抗旱鉴定指标的研究. 西北农业学报, 2012, 21(2): 72-77.
ZHANG X, XIE X Y. Studies on identification indexes of drought resistance by PEG during seed germination of rapeseed (Brassica napusL.).Acta Agriculturae Boreali-occidentalis Sinica, 2012, 21(2): 72-77. (in Chinese)
[16] 谢小玉, 张霞, 张兵. 油菜苗期抗旱性评价及抗旱相关指标变化分析. 中国农业科学, 2013, 46(3): 476-485.
XIE X Y, ZHANG X, ZHANG B. Evaluation of drought resistance and analysis of variation of relevant parameters at seedling stage of rapeseed (Brassica napusL.).Scientia Agricultura Sinica, 2013, 46(3): 476-485. (in Chinese)
[17] ABBASIAN A, RAD A H S. Investigation the response of rapeseed cultivars to moisture regimes in different growth stages.Journal of Central European Agriculture, 2011, 12(2): 353-366.
[18] MENSAH J K, OBADONI B O, ERUOTOR P G, ONOMEIRIEGUNA F. Simulated flooding and drought effects on germination, growth and yield parameters of sesame (Sesamum indicumL.).African Journal of Biotechnology, 2006, 5(13): 1249-1253.
[19] 孙建, 饶月亮, 乐美旺, 颜廷献, 颜小文, 周红英. 干旱胁迫对芝麻生长与产量性状的影响及其抗旱性综合评价. 中国油料作物学报, 2010, 32(4): 525-533.
SUN J, RAO Y L, LE M W, YAN T X, YAN X W, ZHOU H Y. Effects of drought stress on sesame growth and yield characteristics and comprehensive evaluation of drought tolerance.Chinese Journal of Oil Crop Sciences, 2010, 32(4): 525-533. (in Chinese)
[20] 唐富福. 水稻农艺和品质若干性状的全基因组初步关联分析[D].杭州: 浙江大学, 2013.
TANG F F. Genome-wide association analysis in some agronomic and quality traits of rice[D]. Hangzhou: Zhejiang University, 2013. (in Chinese)
[21] 薛瑞, 周广奇, 胡新文, 郭建春, 唐燕琼. 柱花草种质抗旱性综合评价. 中国农学通报, 2009, 25(11): 224-233.
XUE R, ZHOU G Q, HU X W, GUO J C, TANG Y Q. Comprehensive evaluation of drought resistance in stylosanthes germplasms.Chinese Agricultural Science Bulletin, 2009, 25(11): 224-233. (in Chinese)
[22] 黎冬华, 刘文萍, 张艳欣, 王林海, 危文亮, 高媛, 丁霞, 王蕾, 张秀荣. 芝麻耐旱性的鉴定方法及关联分析. 作物学报, 2013, 39(8): 1425-1433.
LI D H, LIU W P, ZHANG Y X, WANG L H, WEI W L, GAO Y, DING X, WANG L, ZHANG X R. Identification method of drought tolerance and association mapping for sesame (Sesamum indicumL.).Acta Agronomica Sinica, 2013, 39(8): 1425-1433. (in Chinese)
[23] 孙建, 张秀荣, 张艳欣, 王林海, 黄波. 湿害处理对不同生育时期芝麻叶片保护酶活性和种子产量的影响. 应用与环境生物学报, 2009, 15(6): 790-795.
SUN J, ZHANG X R, ZHANG Y X, WANG L H, HUANG B. Effects of waterlogging on leaf protective enzyme activities and seed yield of sesame at different growth stages.Chinese Journal of Applied & Environmental Biology, 2009, 15(6): 790-795. (in Chinese)
[24] 刘海卿, 孙万仓, 刘自刚, 武军艳. 北方寒旱区白菜型冬油菜抗寒性与抗旱性评价及其关系. 中国农业科学, 2015, 48(18): 3743-3756.
LIU H Q, SUN W C, LIU Z G, WU J Y. Evaluation of drought resistance and cold resistance and research of their relationship at seedling stage of winter rapeseed (Brassica campestrisL.) in cold and arid regions in North China.Scientia Agricultura Sinica, 2015, 48(18): 3743-3756. (in Chinese)
[25] 祁旭升, 王兴荣, 许军, 张建平, 米君. 胡麻种质资源成株期抗旱性评价. 中国农业科学, 2010, 43(15): 3076-3087.
QI X S, WANG X R, XU J, ZHANG J P, MI J. Drought-resistance evaluation of flax germplasm at adult plant stage.Scientia Agricultura Sinica, 2010, 43(15): 3076-3087. (in Chinese)
[26] POREBSKI S, BAILEY L G, BAUM B R. Modification of a CTABDNA extraction protocol for plants containing high polysaccharide and polyphenol components.Plant Molecular Biology Reporter, 1997, 15: 8-15.
[27] ZHANG Y X, ZHANG X Y, HUA W, WANG L H, CHE Z. Analysis of genetic diversity among indigenous landraces from sesame (Sesamum indicumL.) core collection in China as revealed by SRAP and SSR markers.Genes and Genomics, 2010, 32: 207-215.
[28] KADKHODAIE A, ZAHEDI M, RAZMJOO J, PESSARAKLI M. Changes in some anti-oxidative enzymes and physiological indices among sesame genotypes (Sesamum indicumL.) in response to soil water deficits under field conditions.Acta Physiology Plant, 2014, 36: 641-650.
[29] CONDON A G, RICHARDS R A, REBETZKE G J, FARQUHAR G D. Breeding for high water use efficiency.Journal of Experimental Botany, 2004, 55: 2447-2460.
[30] SUBRAHMANYAM D, SUBASH N, HARIS A, SIKKA A K. Influence of water stress on leaf photosynthetic characteristics in wheat cultivars differing in their susceptibility to drought.Photosynthetica, 2006, 44: 125-129.
[31] SUBRAHMANYAM D, SUBASH N, HARIS A, SIKKA A K. Influence of water stress on leaf photosynthetic characteristics in wheat cultivars differing in their susceptibility to drought.Photosynthetica, 2006, 44: 125-129.
[32] 朱宗河, 郑文寅, 张学昆. 甘蓝型油菜耐旱相关性状的主成分分析及综合评价. 中国农业科学, 2011, 44(9): 1775-1787.
ZHU Z H, ZHENG W Y, ZHANG X K. Principal component analysis and comprehensive evaluation on morphological and agronomic traits of drought tolerance in rapeseed (Brassica napusL.).Scientia Agricultura Sinica, 2011, 44(9): 1775-1787. (in Chinese)
[33] 杨万明, 王敏, 李贵全, 杜维俊. PEG胁迫下大豆BIL群体芽期性状与耐旱性评价. 中国油料作物学报, 2013, 35(5): 564-571.
YANG W M, WANG M, LI G Q, DU W J. Germination traits of soybean BIL population under PEG stress and assessment of their drought toleranceChinese Journal of Oil Crop Sciences, 2013, 35(5): 564-571. (in Chinese)
[34] 贾寿山, 朱俊刚, 王曙光, 史雨刚, 孙黛珍. 山西小麦地方品种萌发期的抗旱性. 华北农学报, 2011, 26(2): 213-217.
JIA S S, ZHU J G, WANG S G, SHI Y G, SUN D Z. Drought resistance of Shanxi wheat landraces at sprouting stage.Acta Agriculturae Boreali-Sinica, 2011, 26(2): 213-217. (in Chinese)
(责任编辑 李莉)
Drought Resistance of Sesame Germplasm Resources and Association Analysis at Adult Stage
LIU WenPing1, LÜ Wei1, LI DongHua2, REN GuoXiang1, ZHANG YanXin2, WEN Fei1, HAN JunMei1, ZHANG XiuRong2
(1Institute of Economic Crops Research, Shanxi Academy of Agricultural Sciences, Taiyuan 030031;2Institute of Oil Crops Research, Chinese Academy of Agricultural Sciences, Wuhan 430062)
【Objective】By association mapping between 33 SSR markers and 18 sesame drought-resistance phenotypes, identifying the associated loci, providing a basis for drought-resistance gene mapping and functional marker development, identifying drought-resistant sesame germplasm resources and providing parental materials and varieties for drought-resistance breeding.【Method】Sesame germplasm populations were cultivated using pots and stressed by drought-water-drought duplication treatment, morphology and growth index during the drought-stress period and mature period were analyzed, then the combinationmethod of SAS, SPSS, and membership function were used for drought-resistance evaluation. At the same time, association analysis was carried out using GLM and MLM models based on molecular markers and drought-resistance phenotype data.【Result】After drought stress, obviously different responses were observed among materials in the population, each index that related to drought resistance averaged smaller than the control; the averaged coefficient of variation of 18 traits was 0.31, which was higher than the control. T test of all traits between treatment and control showed a significant difference. A total of 10 indices related to drought-resistance were screened out, and 12 germplasms with high drought-resistance were identified by functional and clustering analysis. A total of 170 loci were detected by 33 SSR makers in the tested materials, the average loci number was 5.15 per marker. Genetic structure analysis indicated that the population consists of 2 sub-groups. There were 120 loci and 63 loci that were detected by association analysis based on GLM model and MLM model, respectively, the explanation rate ranged from 3.85% to 14.3%, and from 4.00% to 12.5%. There were 12 loci and 3 loci were detected with higher explanation rate than 10% by GLM and MLM, respectively, 5 loci were detected simultaneously by GLM and MLM. By comparison of the primers sequences with sesame genome, 3 genome regions were presumed located genes related with drought-resistance.【Conclusion】This research selected 12 sesame germplasms with high drought-resistance by comprehensive evaluation method, at the same time, 4033-3 and 4033-2 loci that were detected with the highest explanation rate by association analysis based on GLM model and MLM model, respectively, and the explanation rate were 14.3% and 12.5%.
sesame germplasm; drought resistance; SSR markers; association analysis
10.3864/j.issn.0578-1752.2017.04.003
2016-08-25;接受日期:2016-10-24
国家现代农业芝麻产业技术体系(CARS-15)、山西省科技攻关项目(20140311010-3)、山西省农业科学院科技自主创新能力提升工程项目(2016zzcx-07)、芝麻高产、优质、抗逆新品种选育(16 yzgc 009)
联系方式:刘文萍,Tel:13593405471;E-mail:wenggeping@163.com
