密云水库流域土壤动物群落组成及多样性
2017-04-05林青战王汉男韩娜娜王立秀郭强
林青战++王汉男++韩娜娜 王立秀 郭强++周亮++赵奎军


摘要:为探讨密云水库流域土壤动物的群落组成及物种多样性特征,为密云水库生态系统的保护和可持续发展提供科学依据,于2015年6—7月,采用Tullgren干漏斗法和手捡法对北京密云水库流域的5个生境[杏林(Ⅰ)、紫穗槐林(Ⅱ)、柳树林(Ⅲ)、桑树林(Ⅳ)和沙棘林(Ⅴ)]进行土壤动物调查研究。结果:共捕获土壤动物3 087头,隶属2门4纲15目,其中优势类群有4类,即中气门亚目、等节跳科、球角跳科和双翅目;常见类群有4类,即隐气门亚目、长角跳科、同翅目、鞘翅目幼虫,两者共占总捕获量的94.5%。在空间分布上,除生境Ⅴ外,其他生境土壤动物的垂直分布均呈现明显的表聚性。在5个不同生境中,土壤动物群落的Shannon-Wiener多样性指数的大小顺序为Ⅰ=Ⅲ﹥Ⅴ﹥Ⅳ﹥Ⅱ;Margalef丰富度指数的大小顺序为Ⅰ﹥Ⅲ﹥Ⅴ﹥Ⅱ﹥Ⅳ;Simpson优势度指数的大小顺序为Ⅴ﹥Ⅳ﹥Ⅰ﹥Ⅲ﹥Ⅱ;Jaccard相似性系数表明,Ⅲ和Ⅳ之间为极相似,其他生境两两之间为中等相似。
关键词:密云水库;土壤动物;生态系统;群落组成;多样性
中图分类号:S154.5文献标志码: A
文章编号:1002-1302(2016)12-0445-04
收稿日期:2015-10-15
基金项目:国家基础科学人才培养基金(编号:J1210069-32)。
作者简介:林青战(1991—),男,山东菏泽人,硕士研究生,主要从事土壤动物生态学和弹尾目系统分类学研究。E-mail:linzhan20092513417@163.com。
通信作者:谢桂林,博士,副教授,主要从事弹尾目昆虫系统分类学和土壤动物生态学研究。E-mail:desoria@qq.com。
土壤动物是森林、草地、湿地、农田等陆地生态系统中的重要组成部分[1-2],联系着地上和地下生态系统[3],在物质循环和能量流动等方面起着重要的作用[4],对植被、土壤等环境因子的变化有一定的响应[5-6]。大部分土壤动物是土壤有机物质或动植物残体的分解者,尤其在凋落物分解过程的调节中起到重要作用[7],对保持土壤肥力和生态系统的物质循环有重要意义[8-9],土壤动物在生态系统中的作用不言而喻[10];因此,土壤动物是陆地生态系统物质循环、能量流动和信息传递的中心环节,是生态系统演化的重要驱动因子[11]。同时,土壤动物在自然界中占据了多样性更高、空间尺度更广的生境,具有广谱的生物地理学和生态学探针功能[12],对生境变化高度敏感,可以用来描述生境的特征及指示生境的变化,在各类生态系统生物多样性监测和保护方面具有重要意义[13]。近年来,土壤动物的生态学研究越来越受到学者们的关注[14]。
密云水库是潮白河上最重要的水库,是北京市重要水源地,该流域人地关系紧张,生态环境脆弱[15]。近年来,随着人类活动日益频繁,流域内的地表植被发生了一系列变化,进而对流域生态产生了一系列影响。人为因素的干扰程度决定了局部环境条件,因此影响了物种多样性[16]。本研究以位于密云水库流域5個不同的生境作为试验样地,旨在深入探讨密云水库流域土壤动物群落的组成、结构及其多样性特征,为密云水库生态系统的保护和可持续发展提供科学依据,因此具有重要的生态学意义。
1研究区概况与研究方法
1.1研究区概况
密云水库位于北京市密云区(40°14′~41°05′ N、116°07′~117°30′ E),距北京市中心约100 km,水库最大水面积188 km2,由潮河、白河2个水系组成(图1)。
[TPLQZ1.tif]
密云水库流域属于暖温带半湿润季风型大陆性气候,四季分明,干旱冷暖变化明显,无霜期为150 d左右,日照充足,水热同期,年平均气温为10.8 ℃,降水分布一般是从东南向西北递减,年均降水量为300~700 mm。降水绝大多数集中在汛期,降水所形成的地表径流是河流的主要补给形式。流域内土壤分为褐土、棕壤、草甸土和粟钙土等4类,流域内植被以人工林和天然次生林为主[15,17]。
在密云水库流域选取5个不同生境作为采样地进行调查研究,分别为生境Ⅰ:杏林,属于落叶乔木,高5~8 m;生境Ⅱ:紫穗槐林,属于落叶灌木,高1~4 m;生境Ⅲ:柳树林,属于落叶乔木,高3~10 m;生境Ⅳ:桑树林,属于落叶乔木,高 6~11 m;生境Ⅴ:沙棘林,属于落叶灌木,高1~5 m。各生境的土壤类型均为棕壤,植被都属于人工林。
1.2研究方法
1.2.1样本采集
于2015年6—7月分别在5个采样地取样,每个样地随机选取3个100 cm×100 cm的样方,中小型土壤动物的采集使用环切刀(体积为100 cm3),采用五点取样法取样。因地理条件限制,即乱石块上面的覆土层较浅,故只采2层土样(0~5 cm和5~10 cm)。在样方内随机选取10 cm×10 cm的样方来采集大型土壤动物,同样采集2层,以供手捡。每个样方内采集10个供烘烤的小样样品和2个手捡大样样品,共采集180个样品。
1.2.2分离与鉴定
采用改良的Tullgren漏斗分离装置进行土壤动物分离[18],烘烤时间为48 h。收集到的土壤动物置于75%乙醇中保存。依据《中国土壤动物检索图鉴》[19]、《昆虫分类(上)》[20]等参考书目,在体视显微镜(MOTIC SMZ-168)下鉴定并统计所收集的土壤动物。
1.2.3数据处理
土壤动物多样性特征主要采用Shannon-Wiener多样性指数(H′)、Margalef丰富度指数(M)、Pieluo均匀度指数(J)和Simpson优势度指数(C)来计算,计算公式如下:
Shannon-Wiener多样性指数(H′):H′=-∑PilnPi;
Margalef丰富度指数(M):M=(S-1)/lnN;
Pielou均匀度指数(J):J= H′/lnS;
Simpson优势度指数(C):C=∑(ni/ N)2。
式中,S代表土壤动物的总类群数,Pi代表第i类土壤动物的个体数与土壤动物总个体数的比,ni为第i类群个体数,N为群落的总个体数。
采用Jaccard相似性系数(q)比较群落之间的相似性。计算公式如下:
[JZ]q=c/(a﹢b-c)。
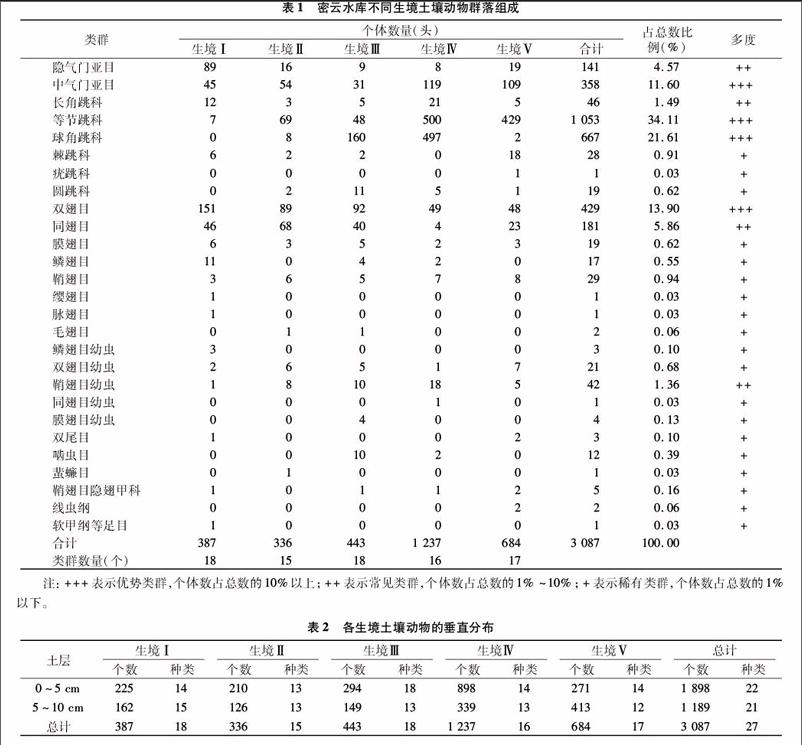
式中,a为A样方内群落的类群数,b为B样方内群落的类群数,c为A、B 2个样方内共有群落的类群数。当0 土壤动物群落多度等级的划分:个体数占总捕获量10%以上的为优势类群,个体数占总捕获量1%~10%之间的为常见类群,个体数占总捕获量1%以下的为稀有类群。 采用BioDiversity Pro、Microsoft Excel进行计算、制图,采用SPSS 18.0进行单因素方差分析和LSD多重比较分析。 2结果与分析 2.1土壤动物群落的组成 本试验获得各种土壤动物27类,共3 087头,分别隶属于2门4纲15目(表1)。其中优势种群为中气门亚目、等节跳科、球角跳科和双翅目4类,占总捕获量的81.22%;常见种群为隐气门亚目(又称甲螨)、长角跳科、同翅目和鞘翅目幼虫4类,占总捕获量的13.28%;其余19类为稀有种群,占总捕获量的5.50%。在所得中小型土壤动物中,数量最多、分布最广的类群是螨类和弹尾类,主要活动在有植物生长的地方。相比之下弹尾类的数量要多于螨类,弹尾类包括长角跳科、等节跳科、球角跳虫科、棘跳科、疣跳科和圆跳科,共 1 814头;螨类包括隐气门亚目和中气门亚目,共499头。 通过分析各生境土壤动物的类群数、类群组成及个体数量发现,各生境的土壤动物类群数大小顺序为Ⅰ=Ⅲ﹥Ⅴ﹥Ⅳ﹥Ⅱ。土壤动物个体数大小顺序为Ⅳ﹥Ⅴ﹥Ⅲ﹥Ⅰ﹥Ⅱ,5个生境的土壤动物个体数差异不显著(F=2.050,P>0.05)。通过进一步的LSD多重比较分析发现,生境Ⅰ与生境Ⅳ、生境Ⅱ与生境Ⅳ之间土壤动物个体数差异显著(P<0.05)。 5个生境的土壤动物群落组成存在差异,许多因素可以影响到土壤动物群落的组成和数量,包括土壤的物理和化学性质(地表凋落物、有机质含量、疏松程度、颗粒大小、污染程度和湿度等)、植被情况、人为干扰等[21]。因此,了解在生境中影响物种偏好的因素,对预测物种的分布和群落组成起到重要作用[22]。 2.2土壤动物的垂直分布特征 各生境土壤动物在0~5 cm和5~10 cm土层的分布如表2所示,生境Ⅰ至生境Ⅳ土壤动物的数量和种类数随着土层深度的加深而降低,呈现出明显的表聚性;而生境Ⅴ的土壤动物数量则随着土层深度的加深而增加,原因为沙棘林地紧靠湖泊,土壤层松软,水分易下渗,同时表层凋落物较少,土壤表层湿度远小于深层,从而土壤动物往深层下移,故表现出与总体分布规律的差异。 土壤动物个体数在不同生境同一土层上的分布存在一定差异。在0~5 cm土层,各生境土壤动物个体数大小顺序为Ⅳ﹥Ⅲ﹥Ⅴ﹥Ⅰ﹥Ⅱ(F=1.947,P>0.05),而LSD多重比较分析发现生境Ⅰ与生境Ⅳ、生境Ⅱ与生境Ⅳ之间土壤动物个体数差异显著(P<0.05);5~10 cm土层,各生境土壤动物个体数大小顺序为Ⅴ﹥Ⅳ﹥Ⅰ﹥Ⅲ﹥Ⅱ(F=1.045,P>0.05)。 2.3土壤动物的多样性指数 在群落研究中,物种多样性指数是代表群落组织水平及其功能特性的综合指标,不同群落具有不同的物种多样性水平[23],反映群落内物种的多少和生态系统食物网的复杂程度,从而能够反映出各生境间的相似性或差异性。土壤动物群落的Shannon-Wiener多样性指数(H′)、Pielou均匀度指数(J)、Simpson优势度指数(C)、Margalef丰富度指数(M)等是土壤动物群落结构、功能和差异性的重要指标,可以反映土壤动物群落结构的稳定性和復杂性。其中群落丰富度和群落均匀度与群落多样性密切相关,一方面,群落内组成类群越丰富,则群落多样性越大;另一方面,群落内个体在类群间分配越均匀,即群落均匀度越大,则群落多样性越大[24]。对于陆地动物群落,通常认为环境条件越优越,动物群落多样性指数越高,种的构成越复杂,种类数目越多,同种的个体数量越少;环境条件越偏离正常,动物群落多样性指数越低,种的构成越简单,群落个性越强,个别种类个体数量越多[25]。 由表3可知,密云水库流域5个不同生境下土壤动物多样性存在一定的差异性。各生境的H′指数大小依次为Ⅰ=Ⅲ﹥Ⅴ﹥Ⅳ﹥Ⅱ,M指数大小依次为Ⅰ﹥Ⅲ﹥Ⅴ﹥Ⅱ﹥Ⅳ,C指数大小依次为Ⅴ﹥Ⅳ﹥Ⅰ﹥Ⅲ﹥Ⅱ,J指数在5种生境中表现出同一性。 2.4土壤动物群落的相似性 计算5个生境土壤动物群落的Jaccard相似性系数(q),比较群落之间的相似性,并对各生境土壤动物群落的相似性系数进行聚类分析。 3结论与讨论 (1)在密云水库流域的5个不同生境中,共捕获土壤动物3 087头,隶属于2门4纲15目。其中优势类群和常见类群构成该地区土壤动物群落的主体,决定整个群落的特性和功能;稀有类群个体总数占总捕获量的5.5%,对环境因素有较高的敏感性,可以作为土壤环境的指示生物。 (2)土壤动物个体数在垂直分布上呈现明显的表聚性(生境Ⅴ除外),受到不同土壤层理化性质的影响。不同生境同一土层的土壤动物个体数分布也存在一定差异。由于植被组成和土壤养分含量差异较大,以及因距离湖泊的距离远近而引起的温湿度不同,使各生境在同一土层的土壤动物种类组成和数量存在差异。在生境Ⅰ、Ⅱ、Ⅲ和Ⅳ中,土壤环境较稳定,土壤动物类群数和个体数是随着土层的加深而减少,符合从地表向下,随土壤深度增加动物类群及数量逐渐降低的规律。在生境Ⅴ中,由于地表土壤较干燥,致使土壤动物往下一定范围内集中。
(3)土壤动物的个体数随生境的变化有很大波动,而类群数变化较小。各群落土壤动物的种类分布与数量分布差异很大,生境Ⅰ、Ⅲ的类群数最多;生境Ⅳ的个体数最多。生境Ⅰ中土壤动物的Shannon-Wiener指数和Margalef数最高,而生境Ⅱ、Ⅳ中的较低,表现出一致性;生境Ⅴ的Simpson指数最高,而生境Ⅱ的最低,这与Shannon-Wiener指数存在差异,是由于生境Ⅴ的土壤特征即表层土壤湿度低于深层导致的。
(4)对密云水库流域土壤动物群落结构组成及物种多样性的进一步研究,可以考虑生态周年的4个不同季节土壤动物的群落动态分析。由于土壤动物群落的结构组成与土壤的理化性质密切相关,可以对不同生境每一土层的土壤理化性质分别进行测定,更深入地了解土壤动物群落结构特征的影响因素,从而为密云水库流域的科学管理、生物多样性的保护、生态环境的改善等提供更多的科学理论依据。
本研究区位于北京密云水库流域,自然环境相对稳定。但是由于近年来的人为活动,影响了该区域土壤动物的多样性。但是关于该区域土壤动物与环境变化的相关性研究较少,有待于进一步的研究来揭示其规律。
致谢:本研究得到国家基础科学人才培养基金子项目“高效农田土壤动物群落结构及生物防治关键技术研究”(编号:J1210069-32)、黑龙江省教育厅科学技术研究项目 “黑龙江省等跳科系统分类及其活性物质抗癌性研究”(编号:12521001)、黑龙江省博士后资助项目“跳虫系统分类及其活性物质应用研究”(编号:LRB09-535)、东北农业大学博士启动基金“中国弹尾纲等跳科系统分类学研究”(编号:2010RCB57)的资助。研究过程中还得到哈尔滨师范大学臧淑英教授的大力支持,在此表示感谢。
参考文献:
[1]章家恩,秦钟,李庆芳. 不同土地利用方式下土壤动物群落的聚类与排序[J]. 生态学杂志,2011,30(12):2849-2856.
[2]李曉东,史沉鱼,覃国乐,等. 濒危植物单性木兰林区土壤动物群落结构与季节动态[J]. 华中农业大学学报,2015,34(4):20-26.
[3]Wardle D A,Bardgett R D,Klironomos J N,et al. Ecological linkages between aboveground and belowground biota[J]. Science,2004,304(5677):1629-1633.
[4]Xia L,Wu F Z,Yang W Q,et al. Contribution of soil fauna to the mass loss of Betula albosinensis leaf litter at early decomposition stage of subalpine forest litter in western Sichuan[J]. Chinese Journal of Applied Ecology,2012,23(2):301-306.
[5]Song B,Yin X Q,Zhang Y,et al. Dynamics and relationship of Ca,Mg,Fe in litter,soil fauna and soil in Pinus koraiensis-broadleaf mixed forest[J]. Chinese Geographical Science,2008,18(3):284-290.
[6]Gao M X,Zhang X P. Fluctuation of soil fauna community during defoliation decomposition under lime and EM treatment[J]. Acta Ecologica Sinica,2011,31(1):164-174.
[7]Zhang W D,Yuan S F,Hu N,et al. Predicting soil fauna effect on plant litter decomposition by using boosted regression trees[J]. Soil Biology & Biochemistry,2015,82:81-86.
[8]Caruso T,Taormina M,Migliorini M. Relative role of deterministic and stochastic determinants of soil animal community:a spatially explicit analysis of oribatid mites[J]. Journal of Animal Ecology,2012,81(1):214-221.
[9]Nielsen U N,Osler G H R,Campbell C D,et al. The enigma of soil animal species diversity revisited:the role of small-scale heterogeneity[J]. PLoS One,2010,5(7):e11567.
[10]尹文英. 土壤动物学研究的回顾与展望[J]. 生物学通报,2001,36(8):1-3.
[11]吾玛尔·阿布力孜,古丽布斯坦·努尔买买提,阿布都肉苏力·吐孙,等. 新疆阜康绿洲不同生境土壤动物群落多样性及其季节动态[J]. 生态学杂志,2013,32(6):1412-1420.
[12]Tan B,Wu F Z,Yang W Q,et al. Characteristics of soil animal community in the subalpine/alpine forests of western Sichuan during onset of freezing[J]. Acta Ecologica Sinica,2010,30(2):93-99.
[13]Bokhorst S,Wardle D A,Nilsson M C,et al. Impact of understory mosses and dwarf shrubs on soil micro-arthropods in a boreal forest chronosequence[J]. Plant and Soil,2014,379(1):121-133.
[14]王振海,殷秀琴,蒋云峰. 长白山苔原带土壤动物群落结构及多样性[J]. 生态学报,2014,34(3):755-765.
[15]Li Y F,Luo Y C,Liu G,et al. Effects of land use change on ecosystem services:a case study in Miyun reservoir watershed[J]. Acta Ecologica Sinica,2013,33(3):726-736.
[16]Thakur M P,Berg M P,Fisenhauer N,et al. Disturbance-diversity relationship for soil fauna are explained faunal community biomass in a salt marsh[J]. Soil Biology & Biochemistry,2014,78(78):30-37.[HJ1.65mm]
[17]陳涛,牛瑞卿,李平湘,等. 密云水库流域植被覆盖度变化对输沙量的影响[J]. 生态环境学报,2010,19(1):152-159.
[18]尹文英,杨逢春,王振中,等. 中国亚热带土壤动物[M]. 北京:科学出版社,1992:4-16.
[19]尹文英,胡圣豪,沈韫芬,等. 中国土壤动物检索图鉴[M]. 北京:科学出版社,1998.
[20]郑乐怡,归鸿. 昆虫分类(上)[M]. 南京:南京师范大学出版社,1999.
[21]Gao Y,Xiong Y,Potapov M. Checklist of Chinese Isotomidae(Collembola) and a description of a new Subisotoma[J]. Zootaxa,2009(2242):55-63.
[22]Heiniger C,Barot S,Ponge J F,et al. Collembolan preferences for soil and microclimate in forest andpasture communities[J]. Soil Biology & Biochemistry,2015,86:181-192.
[23]朱永恒,赵春雨,王宗英,等. 我国土壤动物群落生态学研究综述[J]. 生态学杂志,2005,24(12):1477-1481.
[24]白义,施时迪,齐鑫,等. 台州市路桥区重金属污染对土壤动物群落结构的影响[J]. 生态学报,2011,31(2):421-430.
[25]黄丽荣,李春英,张雪萍,等. 大兴安岭北部森林生态系统土壤动物群落特征[J]. 东北林业大学学报,2009,37(7):75-79.
