鸭抗菌肽Cathelicidin的衍生物设计及抑菌机制研究
2017-04-01冯兴军李丹朱健刘超群高伟陈惠娴
冯兴军,李丹,朱健,刘超群,高伟,陈惠娴
(东北农业大学动物科学技术学院,哈尔滨 150030)
鸭抗菌肽Cathelicidin的衍生物设计及抑菌机制研究
冯兴军,李丹,朱健,刘超群,高伟,陈惠娴
(东北农业大学动物科学技术学院,哈尔滨 150030)
鸭抗菌肽Cathelicidin分子[Cath(1-20)]富含疏水性氨基酸及正电荷,通过人工方法设计合成Cath(1-20)的截短肽衍生物,筛选新型安全Cath(1-20)衍生肽,探讨其作用机制。结果表明,与母肽Cath(1-20)相似,三种衍生肽Cath(5-20)、Cath(4-20)、Cath(3-20)均具有明显广谱抑菌活性,MIC值在1~8μmol·L-1范围内,但仅Cath (5-20)溶血及细胞毒性明显降低,治疗指数达19.5,显著高于Cath(1-20)、Cath(4-20)及Cath(3-20)。Cath(1-20)及其衍生肽可破坏E.coli UB1005的内膜完整性,产生去极化作用,并存在时间和剂量依赖效应;透射电镜结果显示Cath(1-20)及Cath(5-20)可破坏细菌细胞膜。研究证实氨基端的色氨酸对鸭抗菌肽Cath(1-20)溶血及细胞毒性起关键作用,其作用靶点主要为细菌细胞膜。
抗菌肽;鸭;色氨酸;抑菌活性;作用机制
抗菌肽是由生物细胞特定基因编码产生的一类小分子多肽,是有机体非特异性免疫重要组成部分[1],参与特异性免疫功能,在宿主先天和适应性免疫功能中发挥重要作用。
抗菌肽,对细菌、真菌、病毒、寄生虫,甚至肿瘤细胞具有抑制作用,不易产生耐药性[2-3],成为新型抗菌制剂研究热点[4]。然而价格昂贵、抗菌活性不理想、甚至存在溶血及细胞毒性等问题限制其研发应用。因此在研究抗菌肽构效关系及作用机制基础上,以自然存在抗菌肽为模板作改造和设计,筛选理想的新型抗菌肽分子成为重要研发方向[5-7]。
Cathelicidins是广泛存在于人和动物体内的一类抗菌肽,Cathelicidins前体由三部分构成,N端信号肽、C端功能肽以及中间保守cathelin域三部分组成。受特定因子刺激时,C-端成熟肽被蛋白酶水解,发挥生物学功能[8]。Cathelicidins抗菌肽可直接抑制和杀死各种病原体,具有多种免疫调节作用,抑制组织损伤、加快伤口修复、促进血管生成等重要生物学功能[9]。
鸭抗菌肽Cath(1-20)是鸭体内发现的第一种Cathelicidins抗菌肽,抑菌活性较强但对真核细胞毒副作用较大[10]。本研究以设计抗菌肽Cath(1-20)为切入点,筛选细胞选择性高的Cath(1-20)抗菌肽衍生物,阐明Cath(1-20)构效关系,探讨其抑菌机制,为开发新型抗菌肽分子提供理论基础。
1 材料与方法
1.1 材料
1.1.1 菌种及细胞
革兰氏阴性菌:大肠杆菌(Escherichia coliATCC 25922)、大肠杆菌(Escherichia coliUB 1005)、鸡白痢沙门氏菌(Salmonella pullorumC79-13)、鼠伤寒沙门氏菌(Salmonella typhimuriumC77-31),革兰氏阳性菌:金黄色葡萄球菌(StaphylococcusaureusATCC 29213)、表皮葡萄球菌(Staphylococcus epidermidisATCC 12228)、粪链球菌(EnterococcusfaecalisATCC 29212)及人胚胎成纤维细胞MRC-5均为东北农业大学动物营养研究所保存。
1.1.2 试剂
Mueller Hilton肉汤(MHB)培养基购自北京奥博星生物技术有限公司;DMEM培养基、胎牛血清、牛血清白蛋白(BSA)购自浙江天杭生物科技股份有限公司;二甲基亚砜(DMSO)、邻硝基苯β-D-半乳吡喃糖苷(ONPG)购自Amresco(美国);Triton X-100、3-(4,5-二甲基噻唑-2)-2,5-二苯基四氮唑溴盐(MTT)、细胞荧光探针diSC3-5购自Sigma-Aldrich(中国);抗菌肽Cath(1-20)及其衍生肽由上海吉尔生化有限公司采用固相合成法合成,多肽纯度及分子量利用ACQUITYTM高效液相色谱和Waters XevoTMTQMS三重四极杆串联质谱仪(UPLC-MS/MS,美国Waters公司)测定。其他试剂均为国产分析纯。
1.2 方法
1.2.1 肽的设计与合成
利用截取法从氨基端依次删除氨基酸残基,设计合成Cath(1-20)三种衍生肽Cath(5-20)、Cath(4-20)、Cath(3-20)(序列见表1),纯度大于95%。抗菌肽理化参数利用在线工具ProtParam(ExPASyroteomics Server:http://www.expasy.org/tools/ protparam.htm l)及抗菌肽数据库(http://aps.unmc. edu/AP/main.ph)计算获得。
1.2.2 最小抑菌浓度测定
最小抑菌浓度(Minimal Inhibitory Concentrations,MIC)测定参照文献[11]方法。待测菌接种于MHB培养基中,220 r·min-1、37℃过夜培养,转移至新鲜MHB中,继续培养至生长对数期。MHB校正细菌浓度至OD600=0.5,将菌体浓度调整为105cfu·mL-1。将肽溶解于50μL 0.2%BSA(包含0.01%乙酸)溶液,分别取活化培养的菌液50 μL,接种于96孔培养板中,加入溶解抗菌肽(物质的量浓度分别为256、128、64、32、16、8、4、2、1、0.5、0.25μmol·L-1),50μL,37℃培养16~18 h。未加菌孔作为阴性对照,加菌液和MHB孔作为阳性对照,每孔3个平行,3次独立试验。无肉眼可见细菌生长的最低抗菌肽浓度即为该测定抗菌肽对检测菌MIC。
1.2.3 溶血活性测定
参照文献[12]方法通过溶血活性检测肽对哺乳动物血红细胞溶解能力。肝素钠抗凝管收集健康人血液,1 000 r·min-1离心5min,弃上清,收集血红细胞,PBS冲洗3遍,重悬后,得到血红细胞悬液。将肽溶解于PBS中,与红细胞悬液混合,肽终浓度为256、128、64、32、16、8、4、2、1、0.5、0.25μmol·L-1)。以仅加PBS为阴性对照,以含0.2%Triton X-100为阳性对照。37℃孵育1 h,然后1 000 r·min-1,4℃离心5min,吸取上清,酶标仪测定OD570条件下吸光值。每个多肽溶液做3个平行,每组取平均值,3次独立试验。将引起5%溶血活性对应的最低肽浓度定义为最小溶血浓度(Minimal hemolytic concentration,MHC)。抗菌肽溶血率计算公式:
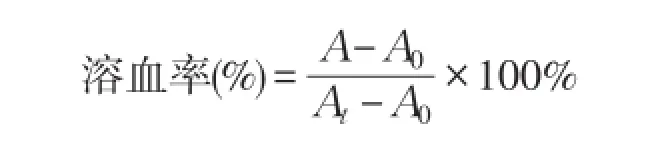
A:抗菌肽样品处理组吸光度值;
At:0.2%Triton X-100阳性对照吸光度值;A0:PBS阴性对照吸光度值。
1.2.4 细胞毒性测定
利用含10%胎牛血清DMEM培养液将对数生长期的MRC-5细胞配成细胞悬液,以每孔2×104个细胞接种到96孔板中,分别加入不同浓度抗菌肽,5%CO2、37℃培养18~24 h,每孔加MTT溶液(0.5mg·mL-1用PBS配制,pH7.4)50μL,37℃继续孵育4 h,弃孔内培养上清液,每孔加150μL DMSO,振荡10min,使结晶物充分融解,测定570 nm吸光值。细胞存活率计算公式:
细胞存活率(%)=(A570处理组)/(A570对照组)×100%。
1.2.5 对细菌细胞膜通透性影响
取对数期大肠杆菌UB1005菌液(108cfu·mL-1)200μL,PBS(pH 7.4)洗涤两次,加入作用浓度为1.5mmol·mL-1的ONPG溶液与1×MIC浓度的抗菌肽溶液,37℃处理1 h,每5min测定420 nm吸光值,不加任何药物作用的菌体组作为阴性对照,以0.2%Triton X-100作用的菌体作为阳性对照。
1.2.6 对细菌细胞膜的去极化作用
参照文献[13]方法,利用细胞膜电势能敏感型荧光染料diSC3-5测定抗菌肽对细胞膜的去极化作用。大肠杆菌UB1005培养至对数生长期,4 000 r·min-1离心5 min,收集菌体,5mmol·L-1HEPES缓冲液(pH 7.4,含20 mmol·L-1葡萄糖和5 mmol·L-1EDTA)洗涤3次后,重悬稀释至OD600= 0.05。向配制的细菌悬浮液中加入终浓度为0.4 μmol·L-1的diSC3-5,室温避光振荡孵育1 h。再加入终浓度为0.1mol·L-1的KCl,室温振荡避光孵育15~30 min,取2mL细菌悬浮液于石英比色皿中,加入不同浓度抗菌肽,在激发波长622 nm,发射波长670 nm条件下测定荧光强度。
1.2.7 透射电镜检测
取对数期大肠杆菌ATCC25922,4000 r·mL-1离心5min,收集菌体,10 mmol·L-1PBS洗涤后,PBS重悬稀释至OD600=0.2,1×MIC浓度的抗菌肽溶液37℃作用1 h,4 000 r·mL-1离心5 min,收集菌体,PBS洗涤3次。5%戊二醛固定2~4 h,PBS洗涤3次,1%锇酸固定80min。离心洗涤,乙醇梯度(50、70、90、100%)脱水,继续用1:1的100%乙醇丙酮混合液、丙酮处理,环氧树脂包埋。切片用醋酸铀和柠檬酸铅染色,HITACHIH-7650透射电镜观察。
2 结果与分析
2.1 肽的设计及其分子特性
由表1可知,通过电喷雾电离质谱(ESI-MS)检测化学合成的肽分子质量与其理论分子质量一致,表明Cath(1-20)及设计的三种衍生肽合成成功。肽平均疏水性由Kyte&Doolittl检测,母肽及其三种衍生肽疏水性不强,三种衍生肽正电荷均为+5。
2.2 溶血活性
由图1可知,母肽Cath(1-20)具有极强溶血活性,在128μmol·L-1浓度下溶血率超过95%,Cath(3-20)也表现极强溶血活性,与母肽溶血活性相当,Cath(4-20)在低浓度下溶血活性较母肽明显降低,但高浓度下溶血活性较强,Cath(5-20)未表现明显溶血活性,即使在最高浓度下(128μmol·L-1),溶血率仅为18%。母肽Cath(1-20)及三种衍生肽Cath(3-20)、Cath(4-20)、Cath(5-20)引起人血红细胞发生5%溶血的最小浓度分别为2、2、8、64μmol·L-1。
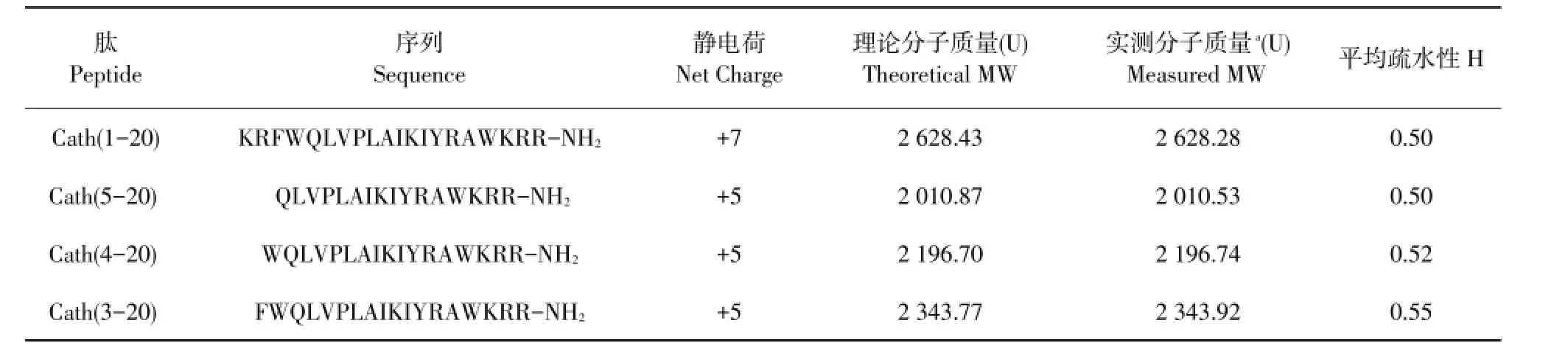
表1 肽的物理化学参数Table 1 Physicochem ical param etersof the peptides
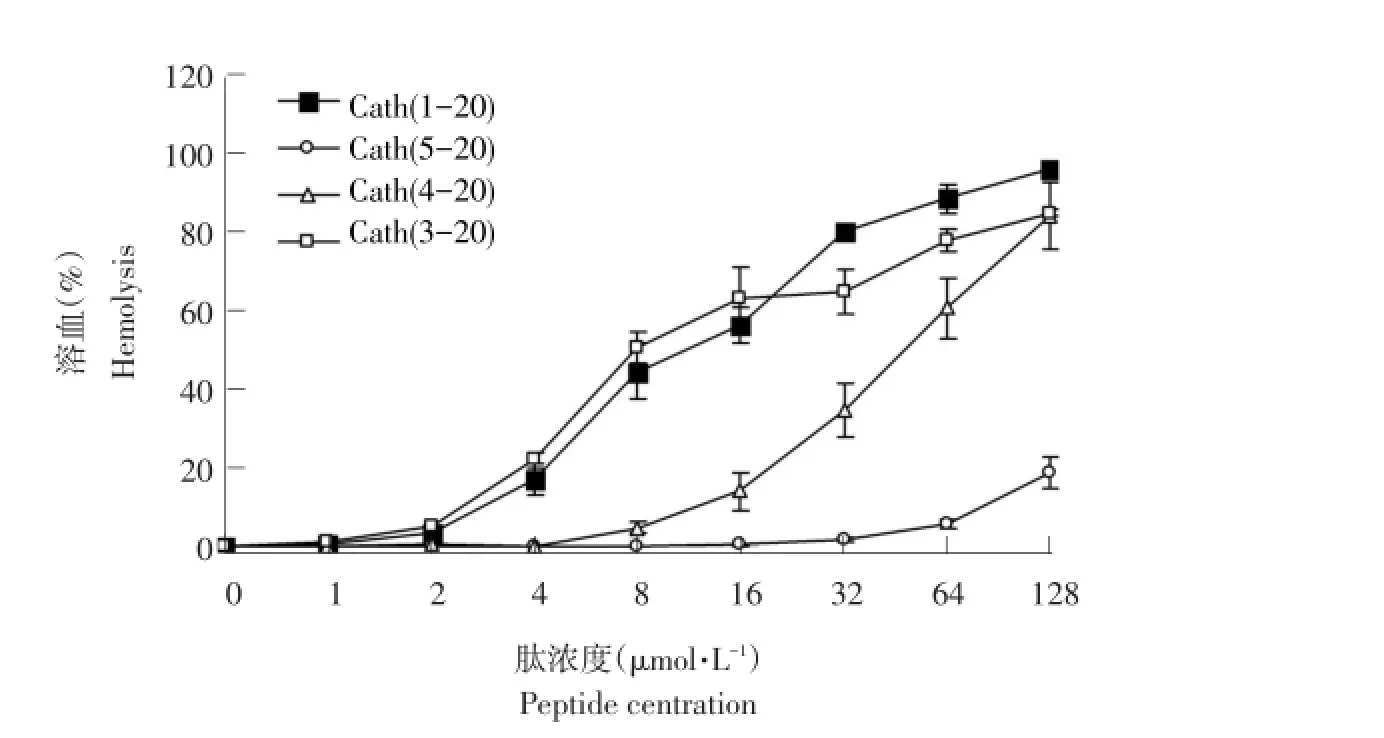
图1 肽的溶血活性Fig.1 Hemolytic activity of the peptides
2.3 细胞毒性
由图2可知,母肽Cath(1-20)及其三种衍生肽Cath(3-20)、Cath(4-20)、Cath(5-20)细胞毒性呈剂量依赖性,随抗菌肽浓度增加,细胞毒性逐渐增强。Cath(1-20)及Cath(3-20)具有极强细胞毒性,Cath(5-20)细胞毒性较母肽明显降低,在低浓度时细胞毒性很低,在128μmol·L-1,细胞死亡为40%,而其他三种肽在该浓度下细胞死亡率均超过90%。
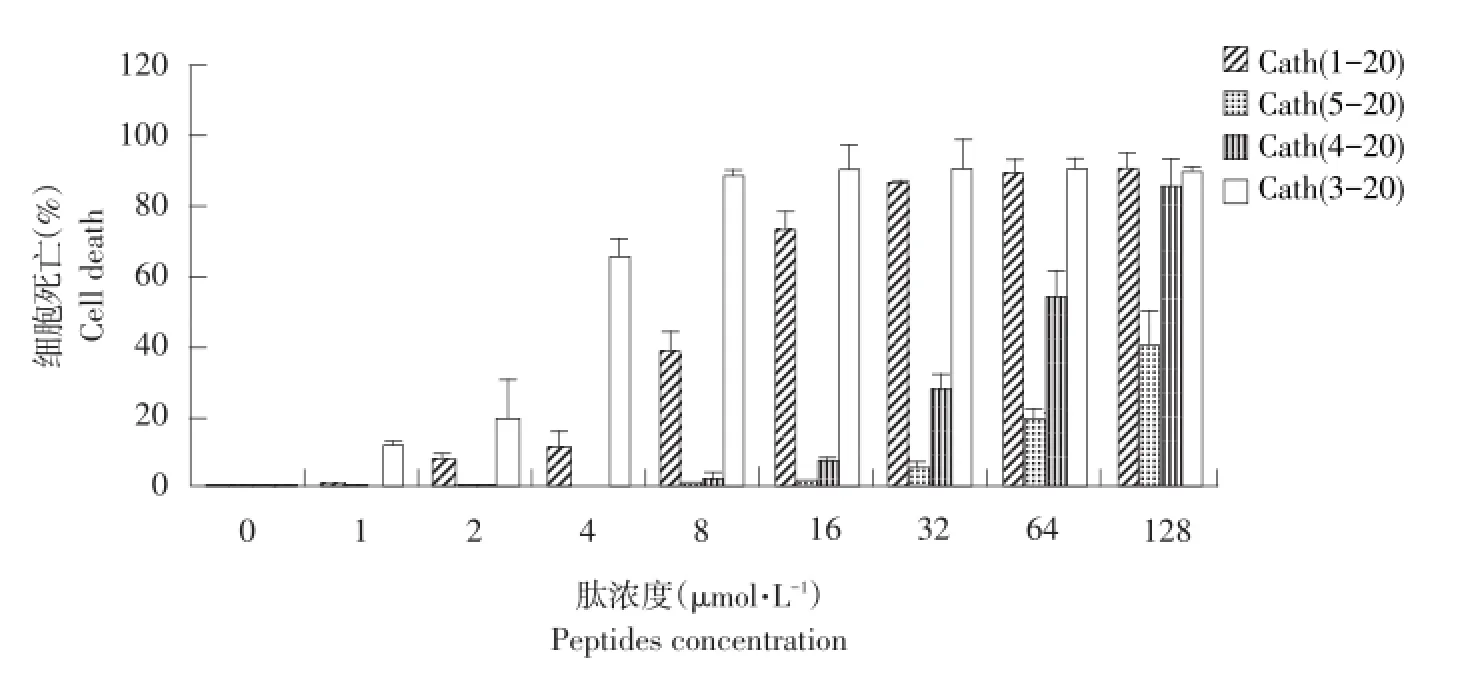
图2 肽的细胞毒性Fig.2 Toxicity of the pep tides
2.4 抑菌活性
采用微量肉汤稀释法测定母肽Cath(1-20)及其三种衍生肽Cath(3-20)、Cath(4-20)、Cath(5-20)体外抑菌活性,见表2。
由表2可知,所有抗菌肽对革兰氏阳性菌和阴性菌均具有较强抑菌活性,MIC为1~8μmol·L-1。与母肽Cath(1-20)相比,三种衍生肽抑菌活性变化不大,但均有一定程度提高。母肽Cath(1-20)治疗指数很低,仅为0.55,三种衍生肽治疗指数均提高,尤其是Cath(5-20),治疗指数达到19.51,较母肽提高35倍,表明Cath(5-20)具有极好细胞选择性。
2.5 对细菌细胞膜通透性影响
试验利用β-半乳糖苷酶对ONPG水解检测抗菌肽对细菌细胞膜通透性影响。β-半乳糖苷酶是细菌细胞质膜上水解酶,当细胞膜被破坏后,细胞膜通透性改变,β-半乳糖苷酶释放到细胞外,同时ONPG进入细胞内部,细胞内外β-半乳糖苷酶与ONPG作用,将其水解成半乳糖和黄色邻硝基苯酚,通过培养液OD值变化得知β-半乳糖苷酶活性,评价细菌细胞膜通透性变化。由图3可知,母肽Cath(1-20)及三种衍生肽均可迅速破坏细菌内膜,呈时间依赖性,说明抗菌肽作用于细胞膜靶向性,通过破坏细胞膜通透性,使胞内物质流至胞外而达到杀菌目的。
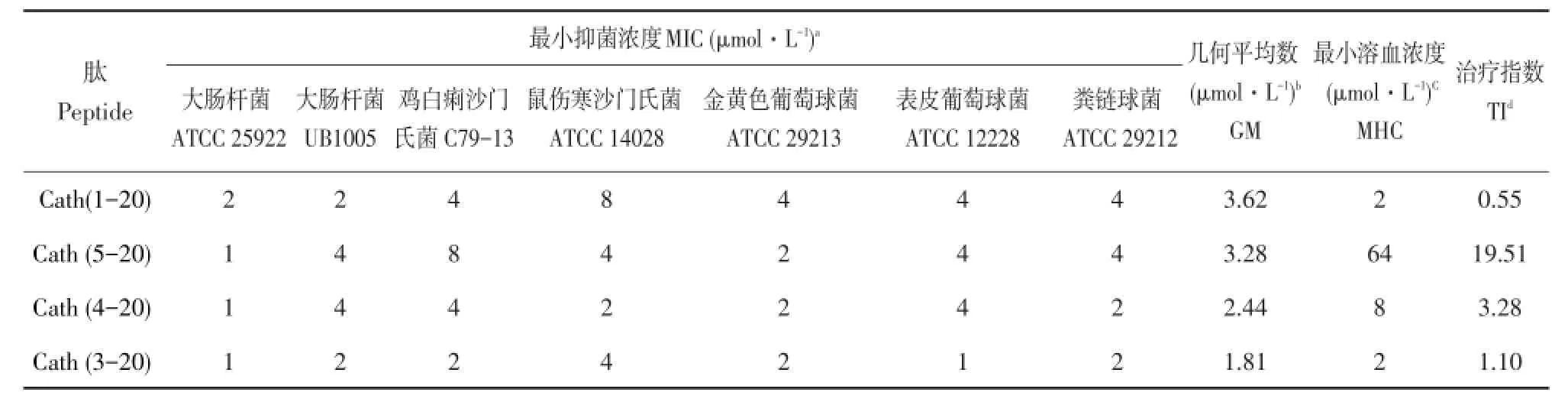
表2 肽的最小抑菌浓度、最小溶血浓度及治疗指数Table2 M IC,MHC and TIof the pep tideand itsderivatives
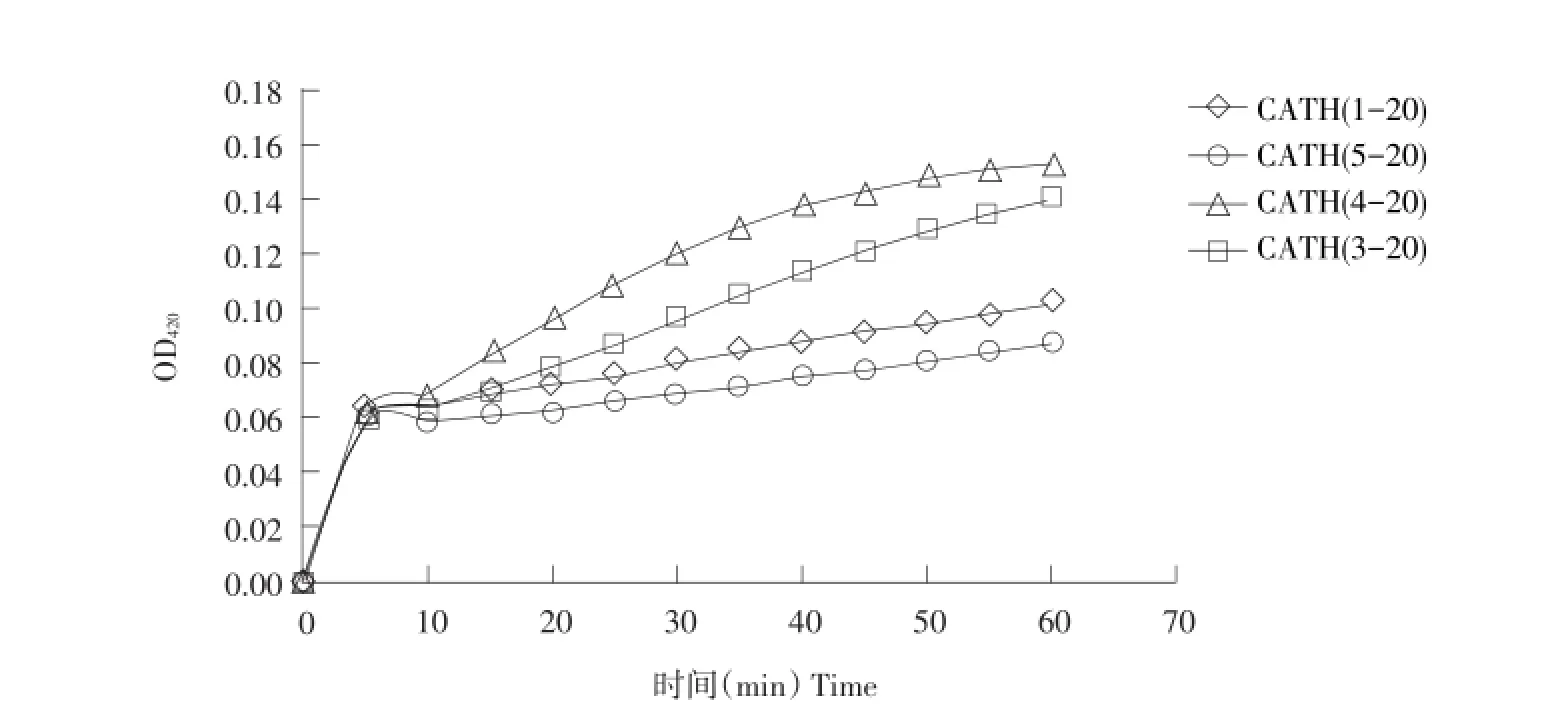
图3 肽对大肠杆菌UB1005细胞膜通透性影响Fig.3 Effectof the peptideson themembrane permeability of E.coli UB1005
2.6 细菌细胞质膜去极化
diSC3-5作为一种膜电位敏感染料,进入细胞后发生聚集并自行荧光淬灭。如果膜被破坏或形成通道,diSC3-5将释放到介质中引起荧光增加,表明细胞膜去极化过程。因此可通过检测荧光释放量衡量抗菌肽对细胞质膜破坏程度。由图4可见,随着肽加入,可检测细菌细胞膜发生去极化,其强度具有时间效应。母肽Cath(1-20)去极化过程的速度与强度最弱。三种衍生肽均强于母肽,其中Cath(4-20)最强,Cath(3-20)次之,Cath(5-20)最低,说明Cath(1-20)及其衍生肽可更快更有效破坏细胞内膜。
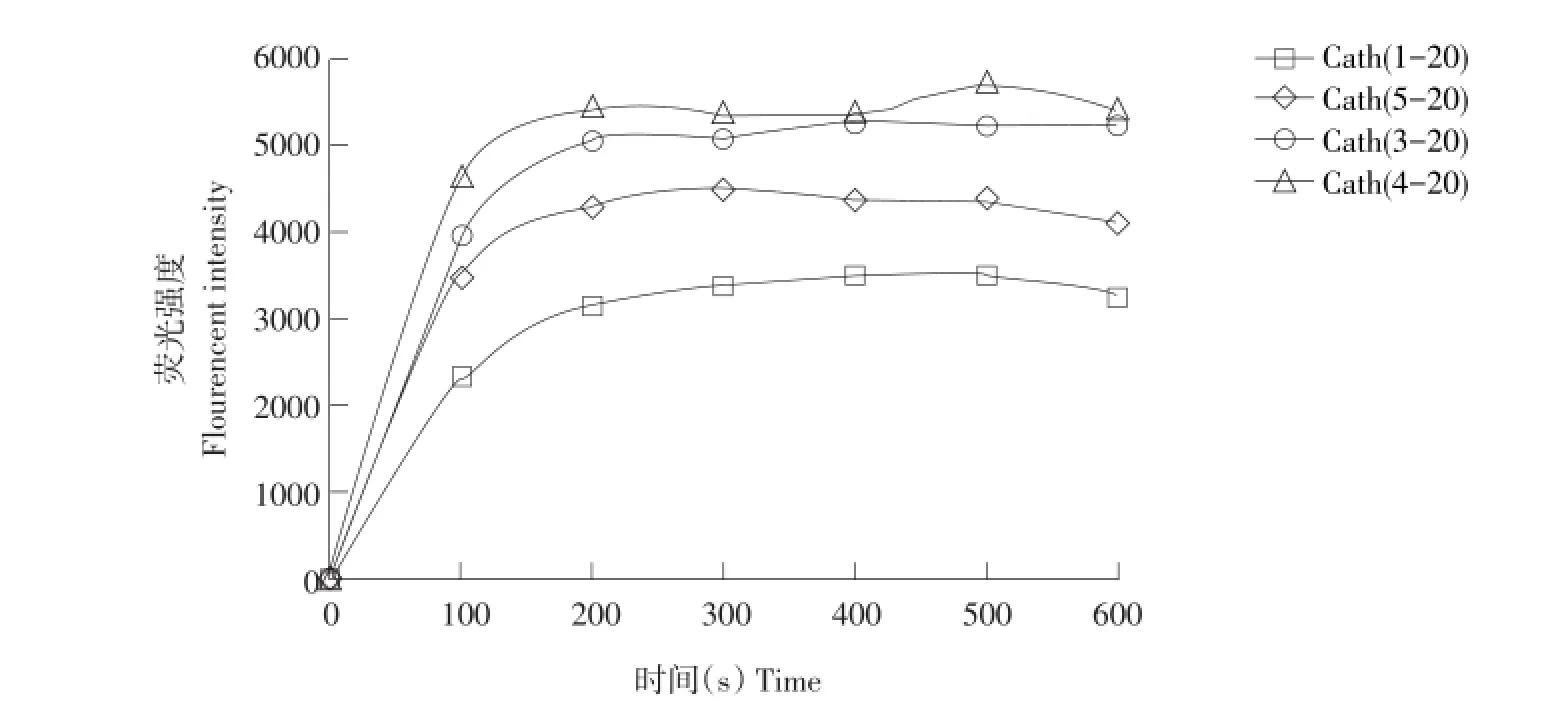
图4 肽对大肠杆菌UB1005细胞膜去极化作用Fig.4 Cytop lasm icm em brane depolarization of thepeptide against E.coli UB1005
2.7 对细菌结构影响
未加抗菌肽和抗菌肽作用1 h后大肠杆菌透射电镜照片(见图5)。未加抗菌肽作用的对照菌体(图5 A)细胞膜边缘清晰完整,内部结构致密,胞质内容物未见明显散失。而经抗菌肽处理大肠杆菌菌体(图5B、C)细胞膜不完整,胞膜形态混乱。内部结构和对照组相比改变明显,内容物释放,形成“鬼影”细胞(失去细胞质但具有细胞壁等外架结构)。电镜图反映抗菌肽对细胞膜完整性的破坏,导致细胞内容物释放。
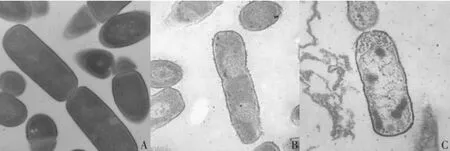
图5 大肠杆菌ATCC25922在抗菌肽作用后结构形态变化Fig.5 Structu ral change of E.coli ATCC 25922 treated w ith pep tides
3 讨论
抗菌肽作为抗生素替代物,不易产生耐药性、无残留、无污染,作为新型抗菌药物已在广泛应用。与传统抗生素相比,抗菌肽抗菌活性仍不理想,甚至存在细胞毒副作用。结构改造天然抗菌肽是克服天然肽链毒性最常用方法,效果理想。以天然抗菌肽为模板,通过氨基酸替换、自身氨基酸删减与截短、截取两种或两种以上抗菌肽片段拼接是改良抗菌肽抑菌活性、消除细胞毒副作用的有效方法[13-15]。
抗菌肽依赖自带正电荷与细菌细胞膜上负电荷基团发生静电吸引相互结合[16]。抗菌肽自带足够正电荷,是抗菌肽与细菌结合,发挥抑菌作用前提。带有正电荷氨基酸,如精氨酸(R)、赖氨酸(K)是抗菌肽抑菌活性关键。抗菌肽需这些氨基酸提供正电荷与氢键才可与带有负电荷细菌质膜结合。研究表明,利用精氨酸或赖氨酸替代抗菌肽原有氨基酸,增加抗菌肽阳离子特性及其正电荷,可显著增强抗菌肽与细菌质膜结合力,提高抗菌肽抑菌活性[17-19]。鸭抗菌肽Cath(1-20)在氨基酸与羧基端均富含精氨酸和赖氨酸。本研究中,删除精氨酸和赖氨酸衍生肽Cath(3-20)与Cath(4-20)抑菌活性与母肽Cath(1-20)相比未降低。表明氨基端精氨酸和赖氨酸的存在对Cath(1-20)抑菌活性无改善作用,原因可能是衍生肽所带正电荷(+5)对于抗菌肽Cath(1-20)发挥抑菌功能已提供足够阳离子特性。
色氨酸(W)在天然抗菌肽中比例较高,对抗菌肽抑菌活性具有重要影响。作为一种芳香族氨基酸,色氨酸含芳香族侧链结构可作用于脂质双层界面区域,将抗菌肽锚定于细菌质膜上,使抗菌肽破坏细菌细胞膜结构[17,20]。通常色氨酸含量越高,抗菌肽抑菌活性越强。Pasupuleti等通过优化天然抗菌肽的色氨酸含量与比例改善抗菌肽活性,或利用色氨酸设计全新抗菌肽分子[21-23]。本研究中,删除氨基端色氨酸的衍生肽Cath(5-20)与未删除该氨基酸残基的母肽及另两种衍生肽相比,抑菌活性未降低,但溶血活性及细胞毒性却明显降低。该结果表明,在鸭抗菌肽Cath(1-20)分子中色氨酸数量对其抗菌活性影响不大,该氨基酸是高溶血及细胞毒性关键因素。
破坏细胞膜是抗菌肽发挥抑菌作用重要机制之一[24]。本试验研究抗菌肽Cath(1-20)及其衍生肽对细菌细胞膜作用机制。Cath(1-20)及其衍生肽可破坏细胞膜,快速提高ONPG在细菌胞内浓度,使细胞膜快速去极化。透射电镜结果也表明Cath(1-20)及Cath(5-20)使细菌细胞膜完整性受损,细胞内容物释放,杀死细菌。
本研究通过截短抗菌肽,删除鸭抗菌肽Cath(1-20)氨基酸端氨基酸,获得三种衍生肽Cath(5-20)、Cath(4-20)、Cath(3-20)。体外试验表明,Cath(5-20)抑菌活性与母肽相当,但溶血及细胞毒性很低,可有效消除Cath(5-20)细胞毒副作用,保留抗菌活性,是一种富有开发潜力的新型抗菌肽分子。
[1]FjellCD,Hiss JA,Hancock R E,etal.Designing antimicrobial peptides:form follows function[J].Nat Rev Drug Discov,2012,11 (1):37-51.
[2]Choi K Y,Chow L N Y,Mookherjee N.Cationic hostdefence peptides:multifaceted role in immunemodulation and inflammation [J].JInnate Immun,2012,4(4):361-370.
[3]Mansour SC,Pena O M,Hancock R E.Host defense peptides: Front-line immunomodulators[J].Trends Immun,2014,35(9): 443-450.
[4]Dennison SR,Harris F,MuraM,etal.A novel form of bacterial resistance to the action of eukaryotic host defense peptides,the use of a lipid receptor[J].Biochemistry,2013,52(35):6021-6029.
[5]Brogden N K,Brogden K A.Will new generationsofmodified antimicrobial peptides improve their potential as pharmaceuticals [J]?Int JAntimicrob Agents,2011,38(3):217-225.
[6]SawaiM V,Waring A J,KearneyW R,etal.Impactofsingle-residuemutations on the structure and function of ovispirin/novispirin antimicrobial peptides[J].Protein Eng,2002,15(3):225-232.
[7]Yang S T,Shin SY,Lee CW,et al.Selective cytotoxicity following Arg-to-Lys substitution in tritrpticin adopting a unique amphipathic turn structure[J].FEBSLett,2003,540(1-3):229-233.
[8]Chromek M.The role of the antimicrobial peptide cathelicidin in renaldiseases[J].Pediatr Nephrol,2015,30(8):1225-1232.
[9]Agier J,EfenbergerM,Brzezińska-Błaszczyk E.Cathelicidin impacton inflammatory cells[J].Centr Eur J Immunol,2015,40(2): 225-235.
[10]Gao W,Xing L,Qu P,et al.Identification of a novel cathelicidin antimicrobial peptide from ducksand determination of its functional activity and antibacterialmechanism[J].Sci Rep,2015(5): 17260.
[11]冯兴军,李静,宋雪莹,等.抗菌肽FAPs在毕赤酵母中的重组表达研究[J].东北农业大学学报,2013,44(9):68-72.
[12]马得莹,张婷婷,徐杨,等.重组鸡β-防御素2蛋白的表达与生物学活性鉴定[J].东北农业大学学报,2015,46(8):56-60.
[13]Qu P,GaoW,Chen H,etal.The central hinge link truncation of the antimicrobial peptide fowlicidin-3 enhances its cellselectivity without antibacterial activity loss[J].Antimicrob Agents Chemother,2016,60(5):2798-2806.
[14]Jacob B,Park IS,Bang J K,et al.Short KR-12 analogs designed from human cathelicidin LL-37 possessing both antimicrobial and antiendotoxic activities withoutmammalian cell toxicity [J].JPept Sci,2013,19(11):700-707.
[15]Jittikoon J,Ngamsaithong N,Pimthon J,etal.EffectofN-terminal truncation on antibacterial activity,cytotoxicity and membrane perturbation activity of Cc-CATH3[J].Arch Pharm Res, 2015,38(10):1839-1849.
[16]Jiang Z,Vasil A I,Hale JD,et al.Effects of net charge and the number of positively charged residues on the biologicalactivity of amphipathic alpha-helical cationic antimicrobial peptides[J].Biopolymers,2008,90(3):369-383
[17]Chan D I,Prenner E J,VogelH J.Tryptophan-and arginine-rich antimicrobial peptides:Structures and mechanisms of action[J]. Biochim BiophysActa,2006,1758(9):1184-1202.
[18]Junkes C,Harvey RD,Bruce KD,etal.Cyclic antimicrobial R-, W-rich peptides:The role of peptide structure and E.coli outer and innermembranes in activity and themode of action[J].Eur Biophys J,2011,40(4):515-528.
[19]Han H M,Gopal R,Park Y.Design and membrane-disruption mechanism of charge-enriched AMPs exhibiting cell selectivity, high-salt resistance,and anti-biofilm properties[J].Amino Acids, 2016,48(2):505-522.
[20]Bi X,Wang C,Ma L,etal.Investigation of the role of tryptophan residues in cationic antimicrobialpeptides to determine themechanism of antimicrobial action[J].JAppl Microbiol,2013,115(3): 663-672.
[21]Pasupuleti M,Chalupka A,Morgelin M,et al.Tryptophan endtagging of antimicrobial peptides for increased potency against Pseudomonas aeruginosa[J].Biochim Biophys Acta,2009,1790 (8):800-808.
[22]GopalR,Seo CH,Song P I,etal.Effectof repetitive lysine-tryptophan motifs on the bactericidal activity of antimicrobial peptides[J].Amino Acids,2013,44(2):645-660.
[23]Torcato IM,Huang Y H,Franquelim H G,etal.Design and characterization of novel antimicrobial peptides,R-BP100 and RWBP100,with activity against Gram-negative and Gram-positive bacteria[J].Biochim Biophys Acta,2013,1828(3):944-955.
[24]Mahlapuu M,Håkansson J,Ringstad L,etal.Antimicrobial peptides:an emerging category of therapeutic agents[J].FrontCell InfectMicrobiol,2016(6):194.
Design of derivatives of duck antimicrobial peptide Cathelicidin andantibacterial mechanism
FENG Xingjun,LIDan,ZHU Jian,LIU Chaoqun,GAO Wei,CHENHuixian
(School of Anim al Science and Technology,Nor theast Agricul tural University,Harbin 150030, China)
The duck antimicrobial peptide Cathelicidin [Cath(1-20)] contains a series of amino acidresidues with the characteristics of net positive charge and hydrophobicity. To screen novel derived peptidesof Cath(1-20) with ideal activity and investigated its antibacterial mechanism, truncated peptide derivatives ofCath(1- 20) were designed and synthesized by chemical method. The results showed that similar to theparent peptide Cath(1- 20), its three derivatives, Cath(5-20), Cath(4-20), Cath(3- 20), showed strongantimicrobial activity against a broad range of bacteria in vitro, with minimum inhibitory concentrations in therange of 1-8 μmol· L-1. However, only Cath(5-20) did not show obvious hemolytic activity and cytotoxicity. Thetherapeutic index of Cath(5-20) was 19.5, much higher than those of Cath(1-20), Cath(4-20) and Cath(3-20).Cath(1-20) and its derivatives were able to increase inner membrane permeability and depolarize the cellmembrane of E. coli UB1005 in a dose- and time-dependent manner. The result of transmission electronmicroscopy also showed that Cath(1-20) and Cath(5-20) could destroy bacterial cell membrane. This studydemonstrated that the tryptophan at the amino terminal played crucial role in the hemolysis and cytotoxicityof duck antimicrobial peptide Cath (1-20) and its main target was bacterial cell membrane.
antimicrobial peptides; duck; tryptophan; antibacterial activity; action mechanism
S852.2
A
1005-9369(2017)03-0040-08
时间2017-3-21 14:04:00[URL]http://kns.cnki.net/kcms/detail/23.1391.S.20170321.1404.024.htm l
冯兴军,李丹,朱健,等.鸭抗菌肽cathe licidin的衍生物设计及抑菌机制研究[J].东北农业大学学报,2017,48(3):40-47.
Feng Xing jun,Li Dan,Zhu Jian,et al.Design of derivatives of duck antim icrobial peptide Cathelicidin and antibacteria l mechanism[J].Journalof Northeast Agricultural University,2017,48(3):40-47.(in Chinese w ith English abstract)
2017-02-13
黑龙江省自然科学基金(C2016022);东北农业大学“学术骨干”项目(15XG15)
冯兴军(1978-),男,研究员,博士,硕士生导师,研究方向为动物营养与饲料科学。E-mail:fengxingjun@neau.edu.cn
