热带西太平洋Y3和M2海山微食物网主要类群生态分布与比较*
2017-03-31赵燕楚王超锋张武昌孙晓霞李学刚
赵 丽 赵燕楚 王超锋 张武昌 孙晓霞,李学刚 赵 苑① 肖 天
(1. 中国科学院海洋生态与环境科学重点实验室(中国科学院海洋研究所) 青岛 266071; 2. 青岛海洋科学与技术国家实验室海洋生态与环境科学功能实验室 青岛 266071; 3. 中国科学院大学 北京 100049; 4. 中国科学院海洋研究所 山东胶州湾海洋生态系统国家野外科学观测研究站 青岛 266071)
海洋微食物环(Microbial loop)对海洋生态系统的物质循环和能量传递起着非常重要的作用, 是对经典食物链的有效补充, 海洋微食物网(Microbial food web)是微食物环概念的延伸(Garrisonet al,2000)。海洋微食物网主要包括浮游病毒(Virioplankton)、异养细菌(Heterotrophic bacteria,HB)、聚球藻(Synechococcus, SYN)、原绿球藻(Prochlorococcus, PRO)、微微型真核浮游生物(Picoeukaryotes, PEUK)、自养鞭毛虫(Autotrophic nanoflagellate, ANF)、混合营养鞭毛虫(Mixotrophic nanoflagellate, MNF)、异养鞭毛虫(Heterotrophic nanoflagellate, HNF)和浮游纤毛虫(Planktonic ciliate)等生物类群, 除 SYN、PRO、PEUK和 ANF是自养生物外, 其他类群都是异养生物(张武昌等, 2016)。
不同类型的海洋生态系统存在显著差异。大洋生态系统相对稳定, 通常以寡营养盐、低生产力闻名,深海由于沉积物、热液、冷泉等极端环境的存在, 形成了独特的生态系统。不同于大洋或深海, 海山(seamount)是指自海底升高 1000m、但仍未突出海平面的隆起(Rogers, 1994)。根据海山山顶的深度可将海山分为三类: 浅海山(<200m)、中层海山(200—400m)和深层海山(>400m)(Genin, 2004; 张武昌等, 2014)。海山特殊的地形地貌使其拥有复杂的水文地理环境特征, 从营养来源、栖息地、沉积物形成等多方面造就了海山独特的生境, 因此海山生态系统拥有比大洋更独特的生物学特征, 一直受到国内外学者的广泛关注。目前对于海山微型浮游生态学的研究还很少,仅在东太平洋亚北极Cobb海山报道了浮游纤毛虫的生物量和生态分布情况(Sime-Ngandoet al, 1992), 以及在东北大西洋的Seine和Seldo海山对微生物类群(包括 SYN、PRO、PEUK、HB、ANF和 HNF)群落结构和生物量进行了研究(Mendonçaet al, 2012)。对西太平洋海山区微食物网的研究还未见报道。
本研究首次报道了热带西太平洋Y3和M2海山微食物网主要类群(包括SYN、PRO、PEUK、HB和浮游纤毛虫)的生态分布情况, 比较了两座海山微食物网主要类群生态分布的异同点及其与环境因子的关系, 为系统研究热带西太平洋海山生态系统特征、丰富海山浮游生态学研究提供基础数据支持。
1 材料与方法
1.1 调查站位
2014年12月3日至2015年1月7日搭乘“科学号”调查船对热带西太平洋 Y3海山(8.7°—9.1°N,137.6°—138°E)(A、B 断面, 交叉于 Y3-0 站位)微食物网主要类群进行样品采集; 2016年3月1日至4月1日搭乘“科学号”调查船对热带西太平洋 M2海山(11.1°—11.5°N, 139.1°—139.6°E)(C、D 断面, 交叉于M2-01站位)微食物网主要类群进行样品采集(图1)。
1.2 样品采集及测定
各站用CTD(Sea-Bird SBE 9)测量温度和盐度垂直剖面, 并采用Niskin采水器(KC-Denmark公司, 丹麦)在每个站位分6—13层采集水样。
硝酸盐(NO3-)、亚硝酸盐(NO2-)和磷酸盐(PO43-)样品采集按照海洋调查规范(GB12763.6-2007)进行,分别在各水层采集水样250mL, 0.7µm滤膜(Whatman GF/F)过滤后装入250mL高密度聚乙烯瓶, 氯仿固定后于-20ºC冰箱中保存, 直至带回实验室用 QuAAtro连续流动分析仪(SEAL公司, 德国)分析测定。NO3-,NO2-和PO43-分别通过Cd-Cu还原法、盐酸-萘乙胺比色法和磷钼蓝法进行测定, 其检出限分别为 0.15,0.01和 0.01µmol/L (张文静等, 2016; 邢建伟等,2017)。
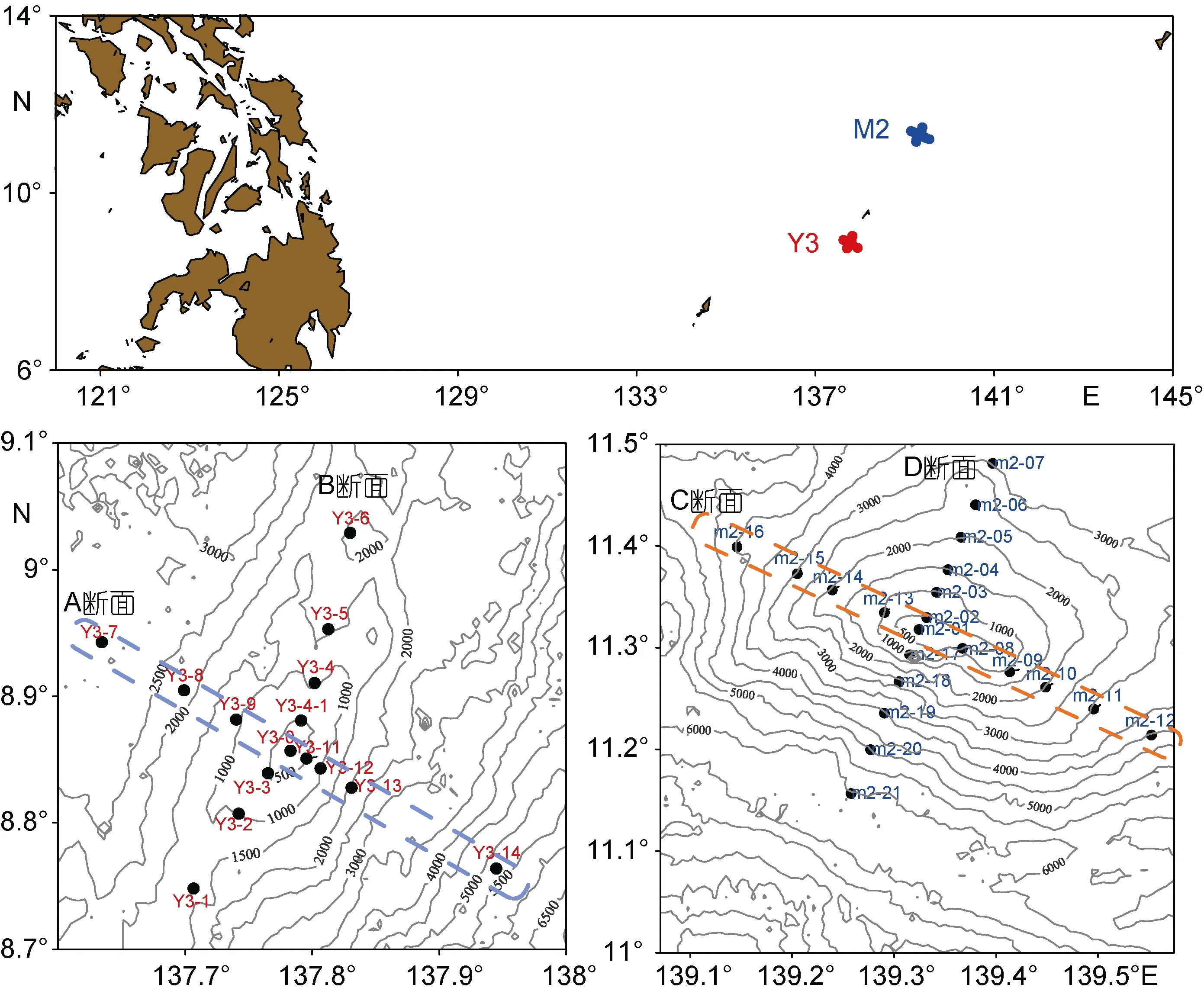
图1 西太平洋Y3(A、B断面)和M2海山(C、D断面)调查站位Fig.1 Sampling stations in the Y3 (Transect A and B) and M2 (Transect C and D) seamounts
微微型浮游生物(SYN、PRO、PEUK和HB)样品采集分别在各水层采集水样 4mL, 加入 10%多聚甲醛进行固定, 终浓度为 1%(V/V), 样品于室温下避光固定20min后置于液氮中冷冻保存, 直至带回实验室进行流式细胞仪分析。在实验室通过流式细胞仪(BD FACSJazz, 美国)检测SYN、PRO、PEUK和HB的丰度。对SYN、PRO和PEUK, 由于其具有自发荧光, 可直接进行检测。对于HB, 由于其丰度较高, 通常先取50μL 海水样品, 加入 250μL TE 缓冲溶液 (Tris-EDTA, 100mmol/L Tris-Cl, 10mmol/L EDTA, pH=8.0,Sigma, 美国)稀释, 并加入 4μL 核酸染料 SYBR GreenⅠ(Molecular Probes, 美国)避光染色 20min后上机检测。
浮游纤毛虫样品分别在各水层采集水样 1L, 用Lugol’s试剂(终浓度 1%)进行固定。样品分析按照Utermöhl方法进行, 沉降至少 48h后虹吸出上清液,只留下约20mL。将20mL样品全部置于沉降杯中沉降, 12h后于 Olympus IX71倒置显微镜下进行镜检(100×)(王超锋等, 2016)。
1.3 生物量转换
微微型浮游生物SYN、PRO、PEUK和HB生物量由其丰度乘以碳转换系数得出, 其转换系数分别为: 100、53、1500和 20fgC/cell (Campbellet al, 1997;Zubkovet al, 2000)。浮游纤毛虫生物量由体积乘以碳转换系数 0.19pgC/µm3得出(Puttet al, 1989)。
1.4 数据分析
采用Surfer 11和Origin 8.5软件绘制作图, 采用IBM SPSS Statistics 19软件进行相关性分析和逐步回归分析。分别对Y3和M2海山真光层各站位的环境参数和微食物网各类群丰度和生物量取算数平均值进行计算比较。
2 结果
2.1 Y3和M2海山水文环境垂直分布特征及比较
Y3海山为中层海山, 山顶距海面300m; M2海山为浅海山, 山顶距海面34m。比较Y3和M2海山真光层环境参数发现, Y3海山温盐、叶绿素a浓度和PO43-浓度平均值低于 M2海山, NO3-和 NO2-则相反(表1)。垂直尺度上, Y3(A和B断面)和M2海山(C和D断面)温度均由表层至底层逐渐降低, 其中Y3海山50m以浅水体混合均匀, 50m以深分层明显; M2海山75m以浅温度基本一致, 75m以深存在明显分层。Y3海山盐度高值在75—100m水层, 50m存在明显盐跃层; M2海山盐度高值在90-110m之间, 未发现明显盐跃层(图2)。Y3海山DCM层多在75—100m水层(张文静等, 2016); M2海山DCM层多集中在110m水层(代晟, 未发表数据)。
对营养盐浓度垂直分布(200m 以浅)进行比较发现, Y3和M2海山NO3-在75m以浅基本耗尽, 100—200m水层 NO3-逐渐升高且分层明显。Y3海山 30m以浅 NO2-基本耗尽, 高值多分布在 75—100m 水层,150m以深NO2-浓度较低; M2海山NO2-呈斑块状分布, 高值主要集中在 75—150m水层。Y3海山 75m以浅区域PO43-浓度基本不超过5µmol/L且分布均匀,75m以深逐渐升高, 且分层现象明显; M2海山150m以浅PO43-浓度较低且呈斑块状分布, 150m以深逐渐升高且出现分层(李学刚, 未发表数据)。
2.2 Y3和M2海山微食物网各类群丰度垂直分布特征及比较

表1 Y3和M2海山真光层水文环境及微食物网不同类群平均值比较Tab.1 Comparison of average environmental parameters,picoplankton and ciliate abundance and biomass between Y3 and M2 seamounts in the euphotic zone
Y3和M2海山均发现SYN、PRO、PEUK、HB和浮游纤毛虫的广泛存在, 其中PRO和HB在丰度上占有绝对优势。SYN、PRO和HB主要分布在真光层区域, 超过200m丰度极低, 可忽略不计。HB和浮游纤毛虫从表层到底层均有分布, 深水层丰度远远低于真光层区域(图 3)。比较真光层微食物网主要类群丰度和生物量平均值发现, Y3海山SYN和HB丰度和生物量平均值略低于M2海山, 而PRO和PEUK则相反(表 1)。
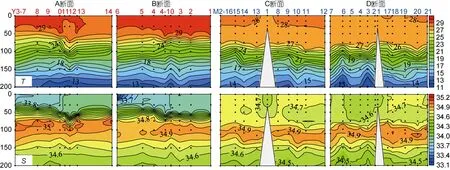
图2 Y3(A、B断面)和M2海山(C、D断面)温度(T, ºC)和盐度(S)垂直分布(200m以浅)Fig.2 Vertical distribution of temperature (ºC) and salinity in the Y3 (Transects A and B) and M2 seamounts (Transects C and D) (in the upper 200m)
由于自养型生物SYN、PRO和PEUK高值多集中在真光层区域, 因此仅对200m以浅区域垂直分布进行比较。微食物网主要类群丰度在两座海山的垂直分布不完全一致。Y3海山除Y3-4和Y3-5站位SYN高值出现在150m外, 其他高值多集中在100m以浅;M2海山SYN高值分布相对较深, 多集中在150m以浅。PRO和PEUK垂直分布基本一致, Y3海山PRO和PEUK高值多分布在50-100m水层, 丰度最大值基本集中在 75m水层, 与叶绿素a浓度分布相似; M2海山PRO和PEUK高值多分布在75—150m水层, 丰度最大值集中在 110m水层(即 DCM 层)。两座海山HB分布基本一致, 从表层至底层丰度逐渐降低, 其中 75m 以浅多呈斑块状分布, 75—150m出现分层,150m以深丰度较低且分布比较均匀。Y3和M2海山浮游纤毛虫分布情况相似, 高值呈斑块状分布于150m以浅海区, 150m以深丰度逐渐降低(图3)。
2.3 Y3和M2海山微食物网主要类群生物量及百分比比较
由于200m以深自养型生物SYN、PRO和PEUK丰度极低, 可忽略不计, 因此仅对 200m以浅区域进行比较。Y3海山微食物网主要类群总生物量平均值(8.99µgC/L)明显高于 M2 海山(6.61µgC/L)。两座海山均是HB生物量最高, 所占百分比超过60%, 自养型生物SYN、PRO和PEUK次之, 所占百分比约为30%,浮游纤毛虫不超过6%(图4)。
垂直尺度上, Y3海山微食物网主要类群总生物量最高值出现在75m水层, 高达18.63µgC/L, 最低值在200m水层, 仅为2.83µgC/L。30m以浅HB占绝对优势, 生物量超过 7.13µgC/L, 所占百分比约为 80%,自养型生物约为15%, 浮游纤毛虫约为5%。75m水层自养型生物占优势, 所占百分比为59.51%, HB生物量略有降低, 所占百分比降至38.07%。100m水层各类群生物量均有所降低, 所占百分比与 75m基本一致。150m各类群生物量明显降低, HB占优势, 百分比为68.52%, 自养型生物约为28.52%。200m水层HB所占百分比为94.11%, 其他各类群所占百分比极低(图5)。
M2海山与 Y3海山不完全一致, 总生物量最高值出现在 110m水层(即 DCM层), 高达 14.15µgC/L,最低值在200m水层, 仅为1.27µgC/L。50m以浅HB占绝对优势, 百分比超过 74.3%, 自养型生物约为20%, 浮游纤毛虫约为5%。75m水层 HB仍占优势,生物量略升高(5.06µgC/L), 但所占百分比降低(61.97%), 自养型生物百分比升至 34.38%。110m水层自养型生物占绝对优势, 百分比为 77.36%, HB为19.95%, 浮游纤毛虫百分比降至2.7%。150m水层各类群生物量明显降低, 但自养型仍占优势, 百分比为55.79%, HB百分比为37.83%。200m水层HB占优势,百分比为61.87%, 自养型百分比为32.76%(图5)。

图3 Y3(A、B断面)和M2海山(C、D断面)聚球藻(SYN, ×103 cells/mL)、原绿球藻(PRO, ×103 cells/mL)、微微型真核浮游生物(PEUK, ×103 cells/mL)、异养细菌(HB, ×105 cells/mL)和浮游纤毛虫丰度(Ciliate, ind./mL)垂直分布(0—200m)Fig.3 Vertical distribution of Synechococcus (SYN, ×103 cells/mL), Prochlorococcus (PRO, ×103 cells/mL), picoeukaryotes (PEUK,×103 cells/mL), heterotrophic bacteria (HB, ×105 cells/mL) and ciliate abundances (Ciliate, ind. /mL) in the Y3 (Transects A and B) and M2 (Transects C and D) seamounts (0—200m)

图4 Y3和M2海山微食物网不同类群生物量及百分比比较Fig.4 Comparison in biomass and the percentage between picoplankton and ciliate in the Y3 and M2 seamounts
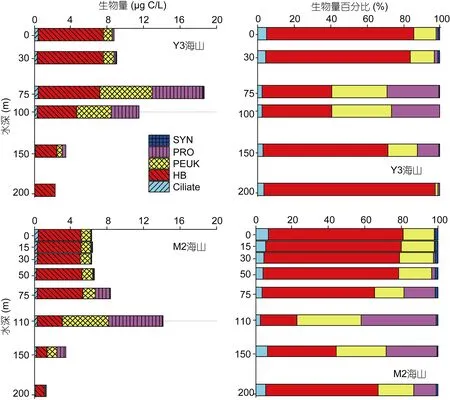
图5 Y3和M2海山在垂直尺度上对不同类群生物量及百分比比较Fig.5 Vertical comparison between picoplankton and ciliate in biomass and the percentage in the Y3 and M2 seamounts
2.4 Y3和M2海山微食物网主要类群丰度与环境因子的相关性分析
对Y3和M2海山所有数据(200m以浅)进行相关性分析, 发现环境因子与各类群之间均存在不同程度的相关性(表2)。Y3海山SYN、PRO、PEUK、HB和浮游纤毛虫之间互为极显著正相关。此外, SYN、HB和浮游纤毛虫均与温度呈极显著正相关, 与营养盐呈极显著负相关; PRO与盐度、叶绿素a和NO2-呈极显著正相关, 与NO3-呈极显著负相关; PEUK与温度、盐度和叶绿素a呈极显著正相关, 与 NO3-和PO43-呈极显著负相关。M2海山除 PRO外, SYN、PEUK、HB和浮游纤毛虫之间互为极显著正相关。此外, SYN、HB和浮游纤毛虫与温度呈极显著正相关,与营养盐呈极显著负相关; SYN、PRO、PEUK和浮游纤毛虫均与盐度和叶绿素a呈极显著正相关; PRO与 NO3-和 NO2-呈极显著负相关, 而 PEUK 与 NO3-和PO43-呈极显著负相关。
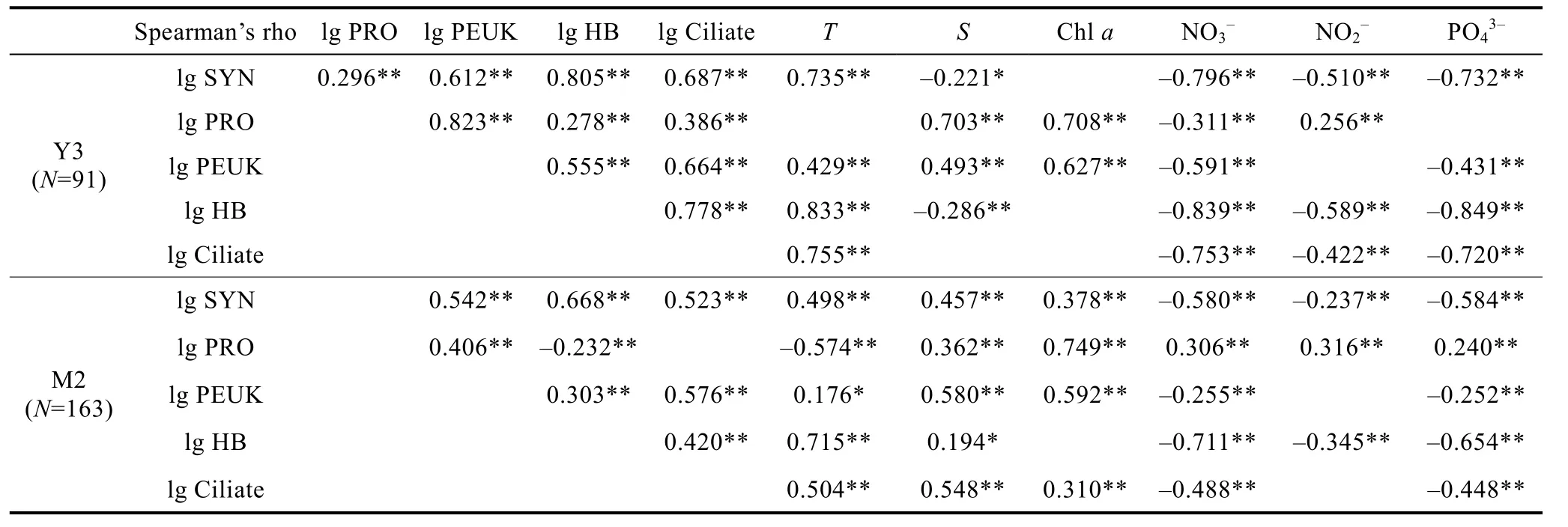
表2 微食物网主要类群与环境、生物因子之间的相关性分析Tab.2 Spearman’s rank correlation coefficient for picoplankton and ciliate in abundance and environmental and biological variables
用逐步回归分析对影响各类群分布的主要因素作进一步研究(表3)。结果表明, Y3海山温度是影响SYN和HB分布的最主要因素, 分别能够解释73.9%SYN和80.9% HB的变化; 盐度和PO43-是影响PRO和 PEUK分布的最主要因素, 分别能够解释 84%PRO和81.9% PEUK的变化; PO43-是影响浮游纤毛虫分布的主要因子, 能解释 70%浮游纤毛虫的变化。与Y3海山不完全一致, M2海山NO3-是影响SYN分布的最主要因素, 能够解释82.9% SYN的变化; 盐度和温度是影响 PRO分布的主要因子, 能解释55.8% PRO的变化; 盐度和NO3-是影响PEUK变化的主要因素, 能解释 77.3% PEUK 的变化; 温度是影响 HB分布的最主要因素, 能解释 92.9%的变化;SYN是影响浮游纤毛虫分布的重要因素, 能解释50.2%的变化。

表3 微食物网主要类群与环境、生物因子之间的逐步回归分析Tab.3 Multiple stepwise regression analysis between picoplankton and ciliate in abundance and environmental and biological variables
3 讨论
3.1 海山区微食物网主要类群垂直分布模式
Veldhuis等(2005)研究了北大西洋典型贫营养海域微微型浮游植物的垂直分布模式, 发现 SYN丰度表层最高, 50m以深丰度明显降低; PRO和PEUK丰度高值分别出现在75m和100m水层, 其他水层丰度明显降低。对海山区微食物网各类群垂直分布模式的研究还很少。本研究发现Y3和M2海山微食物网各类群存在明显的垂直分布特征, 且其分布特征与叶绿素a显著相关。浮游纤毛虫垂直分布呈现显著的“双峰型”模式, 即表层和 DCM 层均发现高丰度值;PRO和 PEUK垂直分布均呈现明显的“单峰型”模式, 即丰度最大值出现在 DCM 层, 其他水层丰度明显降低; SYN和HB峰型相对不显著, DCM层以浅丰度较高, DCM 层以深丰度明显降低(图 6)。Sime-Ngando对东太平洋亚北极Cobb海山浮游纤毛虫进行研究, 发现其丰度高值多集中在 25—50m 水层, 且与叶绿素a显著相关(Sime-Ngandoet al,1992)。尽管 Sime-Ngando未对其垂直分布模式进行总结, 我们可以看出, Cobb海山浮游纤毛虫多为“单峰型”模式。本研究中Y3和M2海山浮游纤毛虫的“双峰型”模式与Cobb海山的“单峰型”模式不完全相似, 但都与叶绿素a有密切关系。浮游纤毛虫在表层和DCM层出现丰度高值的原因, 推测主要与表层的高细菌生产力和DCM层的高初级生产力有关, 它们为浮游纤毛虫提供了丰富的食物饵料。具体原因已在其他文章中作过详细讨论, 在此不做赘述(王超锋等, 2016)。此外, Mendonça等调查了东北大西洋Seine和Sedlo海山不同季节微生物群落(包括SYN、PRO、PEUK、HB、ANF和 HNF)的分布情况, 发现夏季Sedlo海山PEUK丰度高值出现在DCM层(75m), SYN和PRO高值出现在DCM以浅的50m水层, 而Seine海山 SYN、PRO和 PEUK分布与叶绿素a不一致(Mendonçaet al, 2012)。本研究中Y3和M2海山微微型浮游生物垂直分布模式与夏季Sedlo海山比较相似,与Seine海山明显不同。
Y3和M2海山PRO和PEUK垂直分布呈现“单峰型”, DCM层丰度最高, 而SYN和HB峰型不显著, DCM 层以浅丰度较高。分析其原因, 可能是受到温度、光照和营养盐协同作用的影响。温度是影响微微型浮游生物分布的重要因素, 尤其是 PRO,当水温低于15ºC时, PRO生长受到限制(Bucket al,1996)。Y3和M2海山150m以浅水温不低于15ºC, 不会限制微微型浮游生物生长(图2)。光照是寡营养海区影响微微型浮游植物分布的另一重要因素。SYN对光照的依赖性比较强, 在真光层以下无法生长,因此其高丰度值多集中在 DCM 层以浅区域。PRO和 PEUK对光的依赖性相对较弱, 更适应在低光照环境中生长(Mooreet al, 1995; Blanchotet al, 1996)。此外, 营养盐是寡营养海区微微型浮游生物分布的主要限制因子。已有研究表明, 低浓度的 NO3-有利于 SYN 的快速生长, 尤其是当 NO3-浓度在 0.1—3µmol/L时, SYN会成为最重要的初级生产者(Liuet al, 1998; Matsumotoet al, 2004)。Y3和 M2海山DCM以浅区域, NO3-浓度极低, 更适合 SYN生长。对PEUK 来说, 由于个体相对较大, 所需 NO3-浓度相对更高, 因此更适宜在 NO3-浓度较高的环境中生存(李学刚, 未发表数据)。因此, 光照充足、营养盐贫乏的上层水体更适合SYN的生长, 光照相对较弱、营养盐相对丰度的DCM层, 更适合PRO和 PEUK的生长。

图6 Y3和M2海山200m以浅全部站位不同水层微食物网各类群丰度平均值垂直分布及比较Fig.6 Vertical distribution of the average abundance of picoplankton and ciliate in every sampling layer at all stations
3.2 海山区微食物网结构的垂直变化及影响因素
Y3和 M2海山微食物网结构的垂直变化不完全一致。Y3海山30m以浅和150m以深HB生物量占绝对优势, 所占百分比超过70%; DCM层(75—100m)自养型生物(PRO和 PEUK)占绝对优势, 所占百分比约为60%。M2海山75m以浅和200m以深HB占绝对优势, 所占百分比超过60%; 110—150m自养型生物占绝对优势(图5)。M2海山自养型生物占优势的水层(110—150m)要明显深于Y3海山(75—100m), 这可能与它们的海山类型有关。Y3海山山顶站位距海面300m, 上层水体稳定且有明显盐跃层, 真光层深度在150m以浅区域; M2海山为浅海山, 山顶站位距海面仅38m, 山顶区域的生物活动可能会影响到真光层的稳定性, 因此真光层能达到 150m以深区域, 满足PRO和PEUK的生存条件。此外, Y3和M2海山的调查时间分别为 12月和 3月, 季节上的差异也可能带来物理-化学-生物结构的差异, 因此两座海山微食物网结构的垂直变化还可能与采样季节的差异有关。
SYN被认为是寡营养海域初级生产力的重要贡献者, 在热带和温带海域其丰度约为103—105cells/mL, 对总初级生产力的贡献达到20%—80% (Liet al, 1983)。然而在热带西太平洋 Y3和 M2海山,SYN丰度最大值仅为2.31×103cells/mL, 明显低于其他热带和温带海区, 其生物量在各水层的百分比不超过 2%, 可忽略不计(图 5)。HB是微食物网结构的重要组成部分, Y3和M2海山真光层以浅区域, 强光照和寡营养环境不适宜 PEUK和PRO的生长, 而真光层以下自养型生物无法生存, 因此HB生物量百分比占优势。DCM 层适宜的温度、低光照条件以及丰富营养盐, 更适合自养型生物的生长, 因此自养型生物在DCM层占优势(图2, 图3)。由此可见, 海山区微食物网结构的垂直变化主要是受到温度、盐度、光照、营养盐等因素的共同影响。
3.3 海山区与大洋区微食物网结构比较
海山生态系统不同于大洋生态系统, 海山的存在可能改变海流结构如形成上升流或泰勒柱等, 因此海山区与大洋区微食物网的结构也可能存在差异。Blanchot等对赤道太平洋海区 SYN、PRO和 PEUK的垂直分布进行了研究, 结果表明 SYN在垂直尺度上峰型不显著, 从表层到 DCM 层分布基本一致,PRO和PEUK均为“单峰型”, 但PRO丰度高值出现在DCM层以上, PEUK丰度高值在DCM层, 与我们研究的海山区不完全一致(Blanchotet al, 2001)。王超锋等研究了Y3海山和西太平洋海区浮游纤毛虫丰度和生物量, 发现它们在垂直方向上的分布模式基本一致, 均呈现“双峰型”模式(王超锋等, 2016)。
目前对海山区微型浮游生态学的研究还太少,基础数据严重不足, 本研究中Y3和M2海山微食物网垂直分布模式能否代表整个西太平洋海山区, 还需要更多海山航次数据来进一步验证。
4 结语
本研究发现西太平洋Y3和M2海山微食物网各类群的垂直分布模式比较一致, 均与叶绿素a浓度密切相关, 其中浮游纤毛虫垂直分布为“双峰型”模式,丰度高值出现在表层和叶绿素最大值层(DCM); 原绿球藻和微微型真核浮游生物为“单峰型”模式, 丰度高值出现在DCM层; 聚球藻和异养细菌峰型相对不显著, DCM层以浅丰度较高, DCM层以深丰度较低。分析其原因, 可能是受到温度、光照和营养盐协同作用的影响。此外, 对Y3和M2海山微食物网结构的垂直变化进行分析发现, 真光层以浅区域和真光层以下区域异养细菌占优势, DCM 层自养型生物占优势, 主要是受到温度、盐度、光照、营养盐等因素的共同影响。目前对海山区微食物网的研究还太少,
基础数据不足, Y3和M2海山微食物网垂直分布模式能否代表整个西太平洋海山区, 还需要更多海山航次数据作进一步验证。
致谢本研究中温盐数据由中国科学院海洋研究所海洋专项办公室提供, 航次调查采样得到了“科学号”考察船全体工作人员的帮助和支持, 谨致谢忱。
王超锋, 赵 丽, 赵 苑等, 2016. 热带西太平洋浮游纤毛虫的垂直分布. 海洋与湖沼, 47(2): 429—437
邢建伟, 宋金明, 袁华茂等, 2017. 胶州湾夏秋季大气湿沉降中的营养盐及其入海的生态效应. 生态学报, 37(14): 4817—4830
张文静, 孙晓霞, 陈芸燕等, 2016. 2014年冬季热带西太平洋雅浦Y3海山浮游植物叶绿素a浓度及粒级结构. 海洋与湖沼, 47(4): 739—747
张武昌, 于 莹, 李超伦等, 2014. 海山区浮游生态学研究.海洋与湖沼, 45(5): 973—978
张武昌, 陈 雪, 李海波等, 2016. 海洋浮游微食物网对氮、磷营养盐的再生研究综述. 海洋通报, 35(3): 241—251
Blanchot J, André J M, Navarette Cet al, 2001.Picophytoplankton in the equatorial Pacific: vertical distributions in the warm pool and in the high nutrient low chlorophyll conditions. Deep Sea Research Part I:Oceanographic Research Papers, 48(1): 297—314
Blanchot J, Rodier M, 1996. Picophytoplankton abundance and biomass in the western tropical Pacific Ocean during the 1992 El Niño year: results from flow cytometry. Deep Sea Research Part I: Oceanographic Research Papers, 43(6): 877—895
Buck K R, Chavez F P, Campbell L, 1996. Basin-wide distributions of living carbon components and the inverted trophic pyramid of the central gyre of the North Atlantic Ocean, summer 1993. Aquatic Microbial Ecology, 10(3):283—298
Campbell L, Liu H B, Nolla H Aet al, 1997. Annual variability of phytoplankton and bacteria in the subtropical North Pacific Ocean at Station ALOHA during the 1991-1994 ENSO event. Deep Sea Research Part I: Oceanographic Research Papers, 44(2): 167—192
Garrison D L, Gowing M M, Hughes M Pet al, 2000. Microbial food web structure in the Arabian Sea: a US JGOFS study.Deep Sea Research Part II: Topical Studies in Oceanography,47(7—8): 1387—1422
Genin A, 2004. Bio-physical coupling in the formation of zooplankton and fish aggregations over abrupt topographies.Journal of Marine Systems, 50(1—2): 3—20
Li W K W, Rao D V S, Harrison W Get al, 1983. Autotrophic picoplankton in the tropical ocean. Science, 219(4582): 292—295
Liu H, Campbell L, Landry M Ret al, 1998.ProchlorococcusandSynechococcusgrowth rates and contributions to production in the Arabian Sea during the 1995 Southwest and Northeast Monsoons. Deep Sea Research Part II:Topical Studies in Oceanography, 45(10—11): 2327—2352
Matsumoto K, Furuya K, Kawano T, 2004. Association of picophytoplankton distribution with ENSO events in the equatorial Pacific between 145ºE and 160ºW. Deep Sea Research Part I: Oceanographic Research Papers, 51(12):1851—1871
Mendonça A, Arístegui J, Vilas J Cet al, 2012. Is there a seamount effect on microbial community structure and biomass? The case study of seine and Sedlo seamounts(Northeast Atlantic). PLoS One, 7(1): e29526
Moore L R, Goericke R, Chisholm S W, 1995. Comparative physiology ofSynechococcusandProchlorococcus:influence of light and temperature on growth, pigments,fluorescence and absorptive properties. Marine Ecology Progress Series, 116: 259—275
Putt M, Stoecker D K, 1989. An experimentally determined carbon: volume ratio for marine "oligotrichous" ciliates from estuarine and coastal waters. Limnology and Oceanography,34(6): 1097—1103
Rogers A D, 1994. The biology of seamounts. Advances in Marine Biology, 30: 305—350
Sime-Ngando T, Juniper K, Vézina A, 1992. Ciliated protozoan communities over Cobb Seamount: increase in biomass and spatial patchiness. Marine Ecology Progress Series, 89: 37—51 Veldhuis M J W, Timmermans K R, Croot Pet al, 2005.Picophytoplankton; a comparative study of their biochemical composition and photosynthetic properties. Journal of Sea Research, 53(1—2): 7—24
Zubkov M V, Sleigh M A, Burkill P Het al, 2000. Picoplankton community structure on the Atlantic Meridional Transect: a comparison between seasons. Progress in Oceanography,45(3—4): 369—386
