RUVBL2基因knock—down对Rad51和γ—H2AXfoci的影响
2017-03-27韩潆仪
韩潆仪

摘 要:用脂质体转染的方法将特定的RUVBL2 siRNA序列转染到真核细胞,并鉴定其在人肺纤维细胞(MRC5 SV)中的表达,以确保RUVBL2基因的knock-down效果。用Rad51和γ-H2AX的特异性抗体进行免疫荧光染色,应用荧光显微镜观察计数Rad51和γ-H2AX foci的数量。Western blot法证实了siRNA以及转染的效率,确定了RUVBL2基因的存在有利于诱导DNA双链断裂(DSB)条件下Rad51和γ-H2AX被招募到DNA损伤位点。
关键词:RUVBL2 siRNA RAD51 γ-H2AX foci
中图分类号:R113 文献标识码:A 文章编号:1672-3791(2017)01(b)-0200-03
在细胞生命活动过程中,基因组DNA常会受到内源和外源因子的攻击而发生不同类型的损伤。其中,对细胞最具伤害性的损伤是DNA双链断裂(double-strand break, DSB)。大分子蛋白复合体INO80就直接参与了DSB修复过程[1]。RUVBL蛋白是INO80复合体的一类重要亚基。RUVBL1(RuvB-like 1)和RUVBL2(RuvB-like 2)同属于ATPases(AAA+ ATPase)家族成員,是细菌DNA 解旋酶RuvB蛋白在哺乳动物中的同源蛋白[2]。
同源重组是DSB修复的一种重要修复方式,RAD51是一种重要的重组蛋白,主要寻找同源DNA的模板并且帮助在损伤DNA与未损伤DNA修复模板之间产生联系[3]。在酵母中,INO80复合体在染色体修饰和染色体重组过程中会将RAD51蛋白招募到DNA损伤位点[4],而在哺乳动物细胞里INO80复合体也会在染色体修饰和染色体重组过程中将RAD51招募到了DNA损伤位点[5]。哺乳动物INO80复合体通过DNA末端切除的作用介导了DSB的修复[6]。RUVBL蛋白的缺失会影响RAD51被招募到DNA双链断裂位点,从而影响到后续进行的DNA末端切除修复[7]。小鼠和人体细胞内,在含有RUVBL蛋白的TIP60复合体缺失或者活性降低的情况下,RAD51被招募至DNA双链断裂损伤位点的进程也会被削弱[8]。也就是说RUVBL蛋白在DNA损伤修复过程,尤其是末端切除修复中有着不可替代的作用。细胞在电离辐射等因素作用下直接诱导形成的DSB可诱导H2AX磷酸化成γ-H2AX,γ-H2AX是DNA 双链断裂的一个标志[9]。细胞在受到DNA损伤后,尤其是DNA双链断裂后,在断裂位点会出现由γ-H2AX形成的焦点。γ-H2AX对于其他蛋白募集到焦点起着非常关键的作用[10]。INO80复合体通过与γ-H2AX相互作用而使得INO80被招募到DNA双链断裂处[11]。在人类细胞中,RUVBL就参与了与γ-H2AX的相互作用[12]。由此可以推测RUVBL蛋白在哺乳动物细胞DNA损伤修复的过程中有着重要的作用。
RUVBL1作为INO80的亚基之一,在DSB修复过程中起到了重要的作用,主要体现在诱导H2AX磷酸化变成γ-H2AX,然后将Rad51修复蛋白招募至DSB位点[13]。由于RUVBL2和RUVBL1的同源性,我们猜测RUVBL2和RUVBL1也有相似的作用。为了探究RUVBL2是否也参与了DSB的修复,在人类细胞里用siRNA的方法降低RUVBL2蛋白表达,X射线处理诱导DSB的形成,探索在此种情况下Rad51和γ-H2AXfoci的形成是否会受到RUVBL2蛋白表达的影响。
1 材料与方法
1.1 材料与试剂
人胚肺成纤维细胞MRC5 SV细胞系为本实验室保存,脂质体Lipofectamine 2000购自Invitrogen公司,DMEM培养基购自Nissui公司,新生胎牛血清购自Biological Industries公司,兔抗RUVBL2(EPR4146)抗体购自Abcam 公司,HRP 标记山羊抗兔IgG(474-1516)购自KPL公司,β-Tubulin抗体(T8328)购自 Sigma公司,HRP标记抗鼠IgG (sc-2005)购自Santa Cruz Biotechnology公司,ECL 发光试剂盒购自Promega公司,DAPI细胞核荧光染剂购自WAKO公司,RAD51抗体(70-002)购自Bio Academia公司,γ-H2AX 抗体(05-636)购自MILLIPORE公司,鼠抗(Alexa Fluor 546)购自Invitrogen公司,兔抗(Alexa Fluor 488)购自Invitrogen公司,荧光封闭剂购自DAKO公司。
1.2 方法
1.2.1 细胞培养
人肺纤维细胞MRC5-SV细胞系用于进行RUVBL2 knock-down的实验。细胞系培养于DMEM培养基,内含有10% 胎牛血清,L-谷氨酰胺(584μg/mL)和卡那霉素(20 μg/mL)。细胞放置37°C,5% CO2培养箱中培养。
1.2.2 siRNA
MRC5-SV细胞培养于直径为10 cm的培养皿中,当细胞密度达到约50%左右,用Lipofectamine 2000(Invitrogen)转染试剂将RUVBL2 siRNA (2~30 nM) 转入细胞。1 d以后将细胞分装成4盘,3d以后收集细胞检测蛋白。siRNA序列为5-gctccacgcagtacatgaaggagta-3。
1.2.3 Western blot分析
细胞培养5 d后收集细胞,低温RIPA 缓冲液(25 mM Tris-HCl (pH 7.6),150 mM,NaCl,1% NP-40,1% sodium deoxycholate和0.1% SDS) 重悬细胞,补充1 mM PMSF 放置10min, 涡旋震荡2次,2 s/次,4°C,22,000 ×g离心20 min,取上清。蛋白浓度的测定用BCA protein assay kit (Pierce)。蛋白样品上样至7.5%SDS聚丙烯酰氨凝胶电泳,转膜仪Trans-Blot Turbotransfer system (BIO-RAD)的作用下蛋白被转移至PDVF 膜。TBST(20 mM Tris-HCl(pH 7.6),150 mM NaCl和0.1% Tween 20)清洗PDVF膜。 用含有1% BSA 的TBST封闭PDVF膜,封闭后用TBST清洗。加入RUVBL2抗体(1:5000稀释),4 ℃ 过夜; 次日用TBST洗3次,10 min/次;加入HRP 标记的山羊抗兔二抗(KPL 474-1516, 1:5000稀释),37 ℃,1 h; TBST 洗3 次,10 min/次。β-Tubulin作为对照蛋白,加入β-Tubulin抗体(1:5000稀释),4 ℃过夜;次日用TBST洗3次,10 min/次; 加入HRP 标记的抗鼠二抗(1:2000稀释)。用ECL显色,ChemiDoc XRS+ system(BIO-RAD)化学发光凝胶成像系统中检测杂交信号。目标条带密度将由Precision Plus Protein WesternC Standard(BIO-RAD)进行计算。
1.2.4 foci免疫荧光染色
细胞需要培养于盖玻片上:用镊子将高温灭菌后的盖玻片(MATSUNAMI S2112 microslide glass)放进直径为10 cm的培养皿中,每片盖玻片承载500 uL的细胞悬液(细胞悬液的浓度约为5×105个细胞/mL)。细胞培养箱培养30 min,细胞于盖玻片附着稳定后,加入10 mL培养基,培养箱中培养24 h。24 h后,真空泵吸除细胞培养基,10 mPBS清洗两次。将盖玻片浸泡于冰甲醇里,冰箱放置20 min,取出盖玻片,移除残留甲醇后,在冰丙酮里浸泡7 s后迅速取出干燥。加入300 uL封闭液封闭盖玻片,室温反应20 min。加入150 uL γ-H2AX/ RAD51抗体(10%BSA 15μL,γ-H2AX 1:150,RAD51 1:400稀釋于PBS),4 ℃过夜;次日用PBS 洗3次,10 min/次;加入二抗150 uL(10%BSA 15μL,抗鼠二抗1:500,抗兔二抗1:1000稀释于PBS),37 ℃,1 h;PBS 洗3次,10 min/次。将盖玻片放入有0.05% tween20%PBS 的罐子里避光放置5 min。取出盖玻片吸干PBS。将100 uL细胞核染液(DAKO:DAPI=1:500)均匀夹在固定有细胞的盖玻片上。将盖玻片放置在载玻片上,用指甲油封住边缘处。最后用荧光显微镜(Olympus FluoView)进行观察。
2 结果与分析
2.1 siRNA效果检测
siRNA序列作用于RUVBL2 mRNA。siRNA被转染进入MRC5-SV细胞, 用Western blot分析法分析了转染3 d后siRNA对RUVBL2 knock-down的效果。最适knock-down浓度为10 nM siRUVBL2,如图1所示。
2.2 RUVBL2knock-down对γ-H2AX的影响
如图2所示,没有抑制RUVBL2蛋白表达时,经X射线照射诱导了DSB形成后,γ-H2AX的foci数量很多,但是抑制了RUVBL2蛋白的表达,再经X射线处理,γ-H2AX foci数量明显要比RUVBL2表达没有被抑制时的γ-H2AX的foci数量要少,这说明RUVBL2有可能参与;了DSB诱导γ-H2AX的形成。
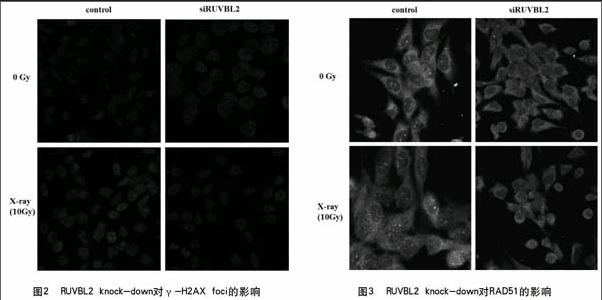
2.3 RUVBL2的knock-down对RAD51 foci的影响
没有抑制RUVBL2蛋白表达时,经X射线照射诱导了DSB的形成,RAD51的foci数量很多,但是抑制了RUVBL2蛋白的表达,再经X射线处理,RAD51 foci数量明显要比没有被knock-down的细胞RAD51的foci数量要少(图3),这说明siRUVBL2影响了RAD51被招募到细胞核,间接说明RUVBL2参与了将RAD51招募到DSB损伤位点处的作用。
3 结论
从荧光免疫的实验来看,我们可以说RAD51和γ-H2AX的foci是受到了RUVBL2 knock-down的影响。对于对照处理细胞来说,10Gy X射线处理过的细胞里RAD51和γ-H2AX的foci数量明显要比没有被射线照射的细胞多。由此我们可以初步得出以下结论:RUVBL2参与了招募RAD51至损伤位点的进程,一旦RUVBL2缺失,将会阻碍DNA损伤触发细胞周期中的信号传导等一系列级联反应,阻碍RAD51被招募到DNA损伤位点。
因此,这个结果揭示了抑制RUVBL2的表达会影响DSB诱发的RAD51分配失调进而阻碍DNA损伤修复应答的进程。
参考文献
[1] 周建杰,陈学峰.ATP依赖型染色质重塑复合物在DNA双链断裂修复中的作用[J].生命科学,2016(4):452-463.
[2] 郑英,谢林俊,张露萍,等.人RUVBL2真核表达载体的构建及其在HeLa 细胞中的表达[J].细胞与分子免疫学杂志,2012, 28(6):586-587.
[3] 乔贵宾.Rad51在同源重组中作用及其与肿瘤关系的研究进展[J].中华肿瘤防治杂志,2010,17(4):306-310.
[4] T.Tsukuda,A.B.Fleming,J.a Nickoloff,et al.Chromatin remodelling at a DNA double-strand break site in Saccharomyces cerevisiae[J].Nature,2005,438(7066):379-383.
[5] T.Ikura,V.V Ogryzko,M.Grigoriev,et al. Involvement of the TIP60 histone acetylase complex in DNA repair and apoptosis[J],Cell,2000,102(4):463-473.
[6] 董立国.人INO80染色质重塑复合物调控基因的分析与基因间调控网络的构建[D].吉林大学,2016.
[7] Gospodinov A,Tsaneva I,Anachkova B. RAD51 foci formation inresponse to DNA damage is modulated by TIP49[J].International Journal of Biochemistry & Cell Biology,2009,41(4):925-933.
[8] 杜凤霞.Tip60磷酸化和p400对Tip60活性及DNA双链断裂修复通路的调控[D].中国科学院北京基因组研究所,2013.
[9] 宾萍,郑玉新.γ-H2AX与DNA双链断裂关系的研究进展[J].卫生研究,2007,36(4):520-522.
[10] 余艳柯,陆源,余应年,等.γH2AX:DNA双链断裂的标志[J].中国药理学与毒理学杂志,2005,19(3):237-240.
[11] A.J.Morrison,J.Highland,N.J.Krogan,et al.INO80 and gamma-H2AX interaction links ATP-dependent chromatin remodeling to DNA damage repair[J].Cell,2004,119(6):767-775.
[12] 樊嵘.Tip60蛋白对细胞DNA损伤反应的调节作用及机制研究[D].中国人民解放军军事医学科学院,2011.
[13] Gospodinov A,Tsaneva I,Anachkova B.RAD51 foci formation in response to DNA damage is modulated by TIP49[J].International Journal of Biochemistry & Cell Biology,2009,41(4):925-933.
