核盘菌Ⅲ型聚酮合酶基因SspksⅢ的克隆与生物信息学分析
2017-03-15李秀丽刘文豪
李秀丽,刘文豪,王 磊
宿州学院生物与食品工程学院,宿州,234000
核盘菌Ⅲ型聚酮合酶基因SspksⅢ的克隆与生物信息学分析
李秀丽,刘文豪,王 磊
宿州学院生物与食品工程学院,宿州,234000
首先利用RT-PCR技术扩增出一个核盘菌的Ⅲ型PKS基因,而后将该片段克隆到T载体中进行测序验证,并将其命名为SspksⅢ。同时,利用生物信息学技术,从理化性质、疏水性、亚细胞定位、跨膜结构域、磷酸化位点、信号肽、二级结构和三级结构等方面对该基因编码蛋白SpksⅢ进行预测和分析。结果表明:SspksⅢ蛋白的相对分子量为49031.8,理论等电点为5.54,氨基酸数目为456,亚细胞定位于细胞质,磷酸化位点主要在丝氨酸残基上,不含信号肽,为非分泌蛋白,在二级和三级结构分析中发现无规则卷曲和α螺旋占主要部分。
核盘菌;聚酮合酶;基因克隆;生物信息学
聚酮化合物是一大类重要的次级代谢产物,在植物、细菌、真菌等生物中广泛存在,具有多种重要的生物学功能。在生物体内,该类化合物由聚酮合酶(Polyketide synthases,PKS)催化合成[1]。PKS以脂酰CoA为底物,通过重复的脱羧缩合过程产生线性聚酮化合物或成环形成环状的酮内酯。根据结构,PKS可分为3种类型[2]:Ⅰ型 PKS,先由多个催化功能域共价连接组成模块,而后多个模块一起构成多功能酶复合物;Ⅱ型PKS,各个催化功能域为独立的蛋白;Ⅲ型PKS大多由酮缩合酶构成,并且在催化聚酮链的延伸过程中不依赖酰基CoA载体蛋白[2]。真菌中的PKS较早被人们认识的是由多个与Ⅰ型PKS相似的催化结构域所组成的巨型多功能蛋白,因此起初真菌的PKS被归为重复性Ⅰ型PKS[3]。2005年,在真菌米曲霉中首次发现了Ⅲ型PKS,接着在曲霉菌、稻瘟病菌、粗糙脉孢菌等真菌基因组中发现假定的Ⅲ型PKS基因[4]。研究表明,粗糙脉孢菌Ⅲ型PKS以长链酰基CoA为起始物,结合丙二酰基CoA进行延伸,合成吡喃酮、间苯二酚、二羟基苯甲酸[5]。相对于Ⅰ型PKSs来说,在真菌基因组中,Ⅲ型PKS在数量上要少得多,但这些酶类发挥着非常重要的作用[6]。因此,研究真菌中Ⅲ型PKS的功能,了解它在真菌代谢和致病过程中的作用,对防治由真菌引起的病害具有重要的意义。
本研究以油菜菌核病的病原菌核盘菌为研究对象。首先运用RT-PCR方法克隆到该菌基因组中一个Ⅲ型PKS基因,而后利用生物信息学技术对其编码蛋白的氨基酸组成、理化性质、疏水性与亲水性、亚细胞定位、跨膜结构域、磷酸化位点、信号肽、二级结构预测和三级结构预测等方面进行初步分析,从而为后续的基因功能研究奠定基础。
1 材料与方法
1.1 材料与试剂
试验以核盘菌NGA4(由安徽农业大学植保学院真菌研究室提供)无性发育阶段的菌丝为材料;实验中所用的大肠杆菌感受态细胞DG5α、TaqDNA聚合酶、pMD18-T vector、DNase I、逆转录酶M-MLV购自宝生物工程(大连)有限公司;提取RNA用的E.Z.N.A.TM Total RNA Kit I试剂盒由美国Omega Bio-Tek公司提供。
1.2 仪器与设备
Bio-Rad Power-Pac 300电泳系统(美国Bio-Rad公司);Eppendorf AG-22331 PCR自动系列化分析仪(德国Eppendorf公司);Milli-QBiocel纯水仪(美国Millipore公司);ZHWY-103B恒温培养震荡器(上海智成分析仪器制造有限公司);Bio-Rad PTC240 PCR仪(美国Bio-Rad公司)。
1.3 总RNA的提取与cDNA合成
将新鲜核盘菌NGA4菌丝在液氮中速冻,而后利用RNA提取试剂盒进行总RNA提取。将提取产物进行琼脂糖凝胶电泳检测后,用DNase酶切进行预处理,以充分去除污染的基因组DNA,而后将所得产物在逆转录酶M-MLV的催化下进行cDNA第一链的合成。
1.4 引物设计与基因SspksⅢ的克隆
用以下两个寡核苷酸作引物对基因SspksⅢ进行扩增,上游引物SspksⅢ-F:5′-ATGCCCAGCAATCCGCCAAC-3′,下游引物SspksⅢ-R: 5′-TTAGT CCAGCTGCACACCATT-3′。以核盘菌NGA4菌丝阶段的cDNA为模板,克隆基因SspksⅢ。扩增条件:预变性95℃ 5 min,94℃变性30 s,56℃退火30 s,72℃延伸70 s,32个循环,10℃保温。PCR产物经电泳检测并回收后,连接至pMD18-T vector上,并转化大肠杆菌感受态细胞DG5α,然后在具有氨苄青霉素抗性的LB培养基平板上过夜得到单菌落。对经PCR验证成功的阳性克隆进行测序。
1.5 基因SspksⅢ的生物信息学分析
对SspksⅢ基因编码蛋白SspksⅢ进行如下几个方面的分析预测。利用ProtParam(http://web.expasy.org/protparam/)进行氨基酸组成和理化性质分析,运用ProtScale(http://web.expasy.org/protscale/)进行疏水性分析,利用Protcomp9.0(http: //www.softberry.com/berry. phtml? topic=protcomppl&group= programs & subgroup=proloc)进行亚细胞定位分析,利用TMHMM Server软件(http://www.cbs.dtu.dk/services/TMHMM/)进行跨膜分析,利用NetPhos2.0(http:// www.cbs.dtu.dk/services/NetPhos/)进行潜在磷酸化位点分析,运用SignalP4.1(http://www.cbs.dtu.dk/services/SignalP/)进行信号肽分析,利用SOPMA(http://npsa-pbil.ibcp.fr/cgi-bin/npsa_automat.plpage=npsa_sopma.html)进行二级结构分析,利用Swiss-model(http: //swiss model.expasy.org/)工具进行三级结构分析。
2 结果与分析
2.1 核盘菌基因SspksⅢ的克隆
提取的RNA如图1所示。28S rRNA的亮度约为18S rRNA的两倍且没有降解,说明提取的RNA完整性较好,可用作为反转录模板,合成cDNA。然后以SspksⅢ-F、SspksⅢ-R为引物进行PCR扩增,琼脂糖凝胶电泳结果如图2。各泳道在大约1400 bp的位置都有清晰单一的条带,将条带回收,经大肠杆菌克隆后进行PCR验证,并选取阳性克隆进行测序验证。结果表明,所克隆到的基因序列同学界公布的序列一致(http://www.Broad institute.org/annotation/genome/sclerotinia_sclerotiorum/MultiHome.html)。

图1 总RNA电泳图
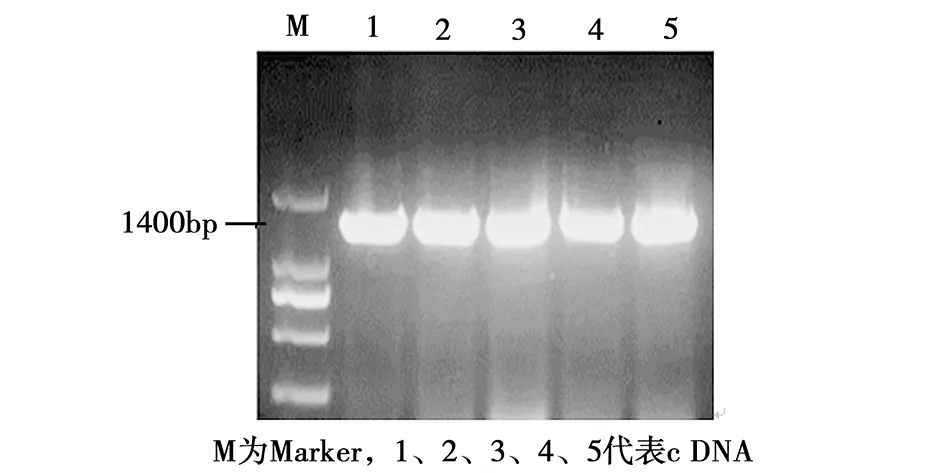
图2 SspksⅢ cDNA电泳图
2.2 蛋白SspksⅢ的生物信息学分析2.2.1 氨基酸组成与理化性质
SspksⅢ蛋白的氨基酸组成种类、数目、所占百分比如表1所示。由表1可知,SspksⅢ蛋白带负电荷残基数量(Asp+Glu)为57,带正电荷残基数量(Arg+Lys)为42;该蛋白质含有2137个碳原子,3436个氢原子,596个氮原子,674个氧原子和24个硫原子,分子式C2137H3436N596O674S24;相对分子量为49031.8,理论等电点为5.54,氨基酸数目为456,其中,酸性氨基酸(DE)占13.6%,碱性氨基酸(KR)占9.2%,中性氨基酸占78.2%。在组成SspksⅢ蛋白的20种氨基酸中,亮氨酸占比最高,达到9.9%,色氨酸占比最低,仅为0.9%。在酵母和大肠杆菌中表达的半衰期分别大于20 h和10 h,在溶液中的不稳定指数为41.78,提示该蛋白为不稳定;脂肪族氨基酸指数为87.46,总平均疏水指数为-0.171。
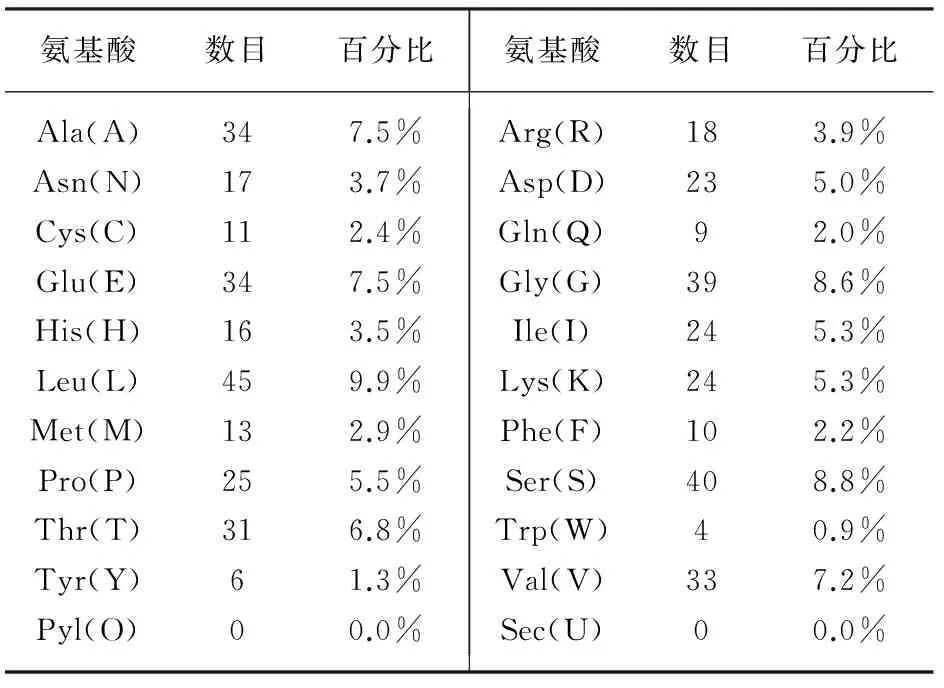
表1 SspksⅢ蛋白的氨基酸组成
2.2.2 疏水性
运用ProtScale软件分析得到SspksⅢ蛋白的疏水性特点,见图3。由图3可以看出SspksⅢ蛋白在152位氨基酸有一个亲水性峰,峰值为2.2,疏水性峰在440位氨基酸,峰值为-2.5,故推测该蛋白可能为疏水性较强的蛋白质。
2.2.3 亚细胞定位分析
利用Protcomp9.0对SspksⅢ蛋白的亚细胞定位进行分析(表2),由表2可知,该蛋白分布于细胞核(1.13)、细胞质(6.67)可能性较大,分布于细胞外、质膜、线粒体、溶酶体、高尔基体的可能性较小,而分布于细浆和液泡的可能性为0,因此推断SspksⅢ蛋白主要在细胞质中发挥生物学作用。
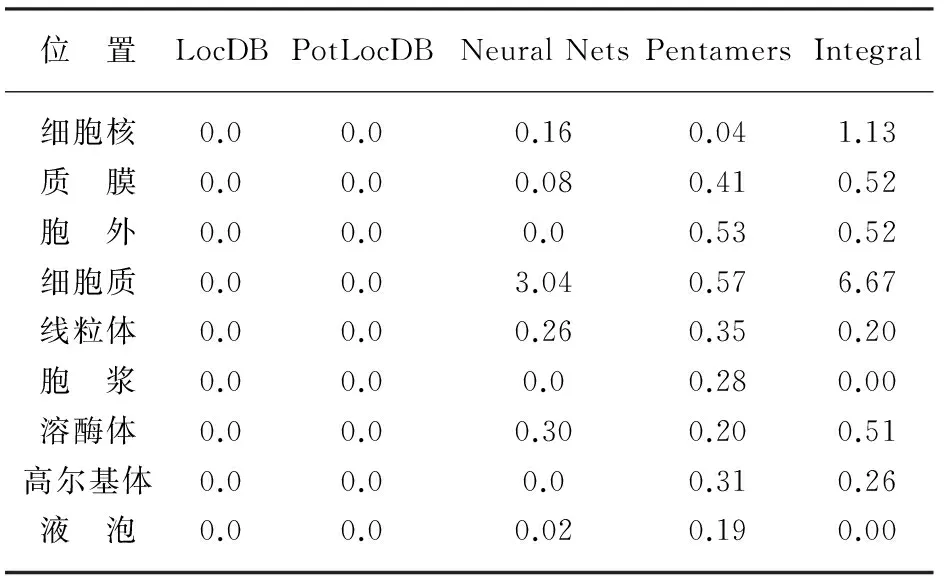
表2 SspksⅢ的亚细胞定位
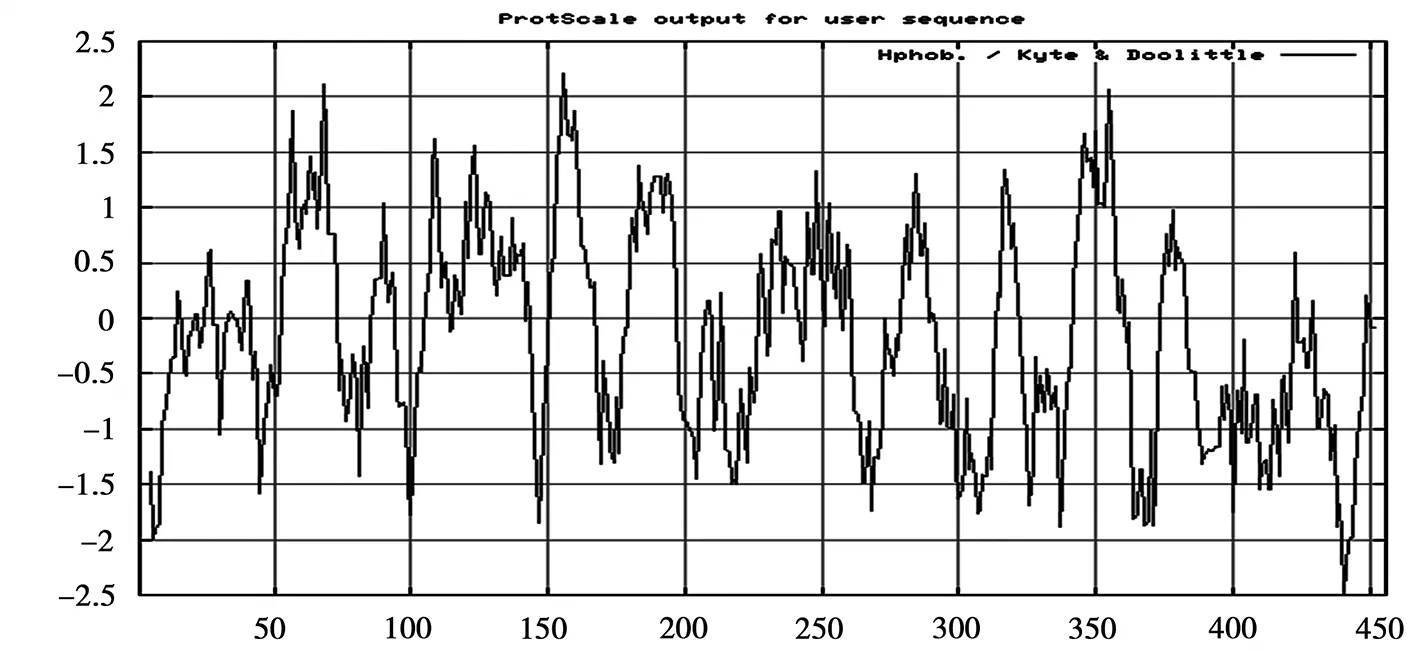
图3 SspksⅢ疏水性分析
2.2.4 跨膜区
利用TMHMM Server 2.0对SspksⅢ蛋白进行跨膜区分析,结果表明该蛋白不具有跨膜区,说明它不是跨膜蛋白,推测主要在基质中发挥作用。
图4 SspksIII蛋白的跨膜结构分析
2.2.5 磷酸化位点
磷酸化是蛋白翻译后的最普遍的修饰方式,SspksⅢ蛋白可能的磷酸化位点如图5。由图5可知,丝氨酸数目为21个,苏氨酸数目为8个,酪氨酸数目为3个,由此可以判断该蛋白的磷酸化位点主要在丝氨酸位点上,部分在苏氨酸和酪氨酸上。
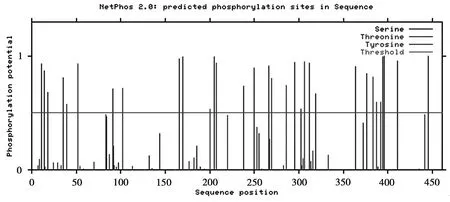
图5 SspksⅢ的磷酸化位点预测
2.2.6 信号肽分析
用Signal-P软件分析得出信号肽分析图(图6),可见信号肽分值都低于0.5,由此可推测该基因所编码的蛋白不存在信号肽,为非分泌蛋白。
图6 SspksⅢ的信号肽分析
2.2.7 二级结构
二级结构分析结果见表4。由4表可以看出,SspksⅢ蛋白主要由三种二级结构形式组成,其中,无规则卷曲占46.27%,α螺旋占36.62%,伸展链占17.11%,不含β折叠结构。

表3 SspksⅢ蛋白的二级结构
2.2.8 三级结构
SspksⅢ蛋白三级结构建模(图7)显示,SspksⅢ存在较多的无规则卷曲结构和α螺旋结构,同二级结构的预测结果一致。
4 讨 论
聚酮化合物具有结构和功能的多态性,其中不乏具有生物活性和重要药用价值的活性物质,比如抗真菌剂灰黄霉素和甲氧基丙烯酸酯类杀菌剂、抗寄生虫抗生素除虫菌素和莫能菌素、免疫抑制剂雷帕霉素等[7]。研究Ⅲ型PKS可以为人工合成某些次生代谢提供依据[8-9]。Ⅲ型PKS在植物、细菌、真菌中都存在,而现有的研究多集中于植物和细菌方面,而对真菌中的Ⅲ型PKS研究较少[10-11],故对该类蛋白进行深入的研究具有一定的理论意义和实践意义。
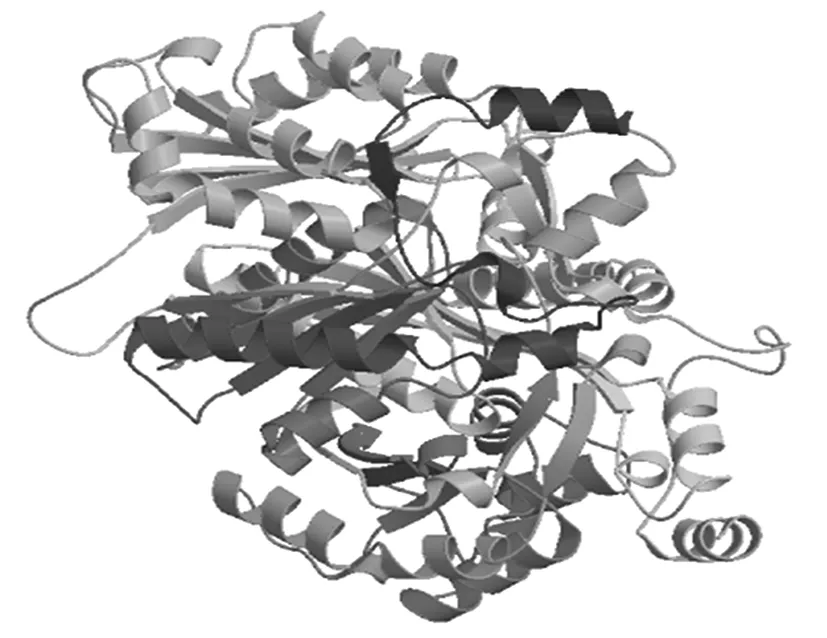
图7 SspksⅢ蛋白的三级结构
本研究利用RT-PCR方法对核盘菌假定PKS基因SspksⅢ进行克隆,并在此基础之上对该基因编码蛋白进行生物信息学分析。结果表明:SspksⅢ蛋白为疏水性较强的蛋白质,不具跨膜结构,不含有信号肽,主要分布于细胞质中,据此可以判定SspksⅢ蛋白主要在细胞质基质中发挥催化功能。另外,该蛋白的二级结构和三级结构表现为无规则卷曲、α螺旋占36.62%、伸展链占主要部分。
[1]Hopwood DA,Sherman DH.Molecular genetics of polyketides and its comparison to fatty acid biosynthesis[J].Annu Rev Genet,1990,24:37-66
[2]Shen B.Polyketide biosynthesis beyond the type Ⅰ,Ⅱ and Ⅲ polyketide synthase paradigms[J].Curr Opin Chem Biol,2003,7(2):285-295
[3]Kroken S L,Glass N L,Taylor J W,et al.Phylogenomic analysis of type I polyketide synthase genes in pathogenic and saprobic ascomycetes[J].Proc Natl Acad Sci USA,2003,100(26):15670-15675
[4]Seshime Y L,Juvvadi P R,Fujii I,et al.Discovery of a novel superfamily of type Ⅲ polyketide synthases in Aspergillus oryzae[J].Biochem Biophys Res Commun,2005,331(1):253-260
[5]Funa N,Awakawa T,Horinouchi S.Pentaketide resorcylic acid synthesis by type Ⅲ polyketide synthase from Neurospora crassa[J].J Biol Chem,2007,282:14476-14481
[6]Hashimoto M L,Nonaka T,Fujii I.Fungal type Ⅲ polyketide synthases[J].Nat Prod Rep,2014,31(10):1306-1317
[7]Austin M B,Noel J P.The chalcone synthase superfamily of type Ⅲ polyketide synthases[J].Nat Prod Rep,2003,20(1):79-110
[8]Katsuyama Y,Ohnishi Y.Type Ⅲ polyketide synthases in microorganisms[J].Methods Enzymol,2012,515:359-377
[9]Yu D,Xu F,Zeng J,et al.Type Ⅲ polyketide synthases in natural product biosynthesis[J].IUBMB Life,2012,64(4):285-295
[10]严少华,郭亮.真菌聚酮合酶分类进展及其应用[J].江西科学,2006,24(2):170-172
[11]许杨,魏康霞.真菌聚酮合酶基因的研究进展[J].食品与生物技术学报,2008,27(2):1-5
(责任编辑:汪材印)
10.3969/j.issn.1673-2006.2017.01.032
2016-08-28
宿州区域发展协同创新中心全国开放课题“夹沟香稻米的香味基因的克隆与序列特异性研究”(2014SZXTKF04ZD)。
李秀丽(1980-),女,山东济宁人,博士,讲师,研究方向:植物病原真菌病害。
Q71
A
1673-2006(2017)01-0117-05
