河北雾灵山森林景观蛾类组成与多样性分析
2017-03-14唐慎言潘昭任卫红项亚飞孟祥普王圆圆任国栋
唐慎言,潘昭,任卫红,项亚飞,孟祥普,王圆圆,任国栋
(1.河北大学 生命科学学院 河北省动物系统学与应用重点实验室,河北 保定 071002;2.河北省森林病虫害防治检疫站 检疫科,河北 石家庄 050000;3.河北雾灵山国家级自然保护区 资源科,河北 承德 067300)
河北雾灵山森林景观蛾类组成与多样性分析
唐慎言1,潘昭1,任卫红2,项亚飞3,孟祥普3,王圆圆3,任国栋1
(1.河北大学 生命科学学院 河北省动物系统学与应用重点实验室,河北 保定 071002;2.河北省森林病虫害防治检疫站 检疫科,河北 石家庄 050000;3.河北雾灵山国家级自然保护区 资源科,河北 承德 067300)
2015年5—9月,通过对河北雾灵山国家级自然保护区7个样点进行森林昆虫多样性普查,初步鉴定得到蛾类昆虫23科436种.针对该结果,使用Simpson多样性指数、Shannon-Wiener多样性指数、Pielou均匀度指数、Whittaker多样性指数和Margalef丰富度指数对其中分布较广的11科进行群落多样性分析,结果发现:雾灵山不同生境下蛾类昆虫多样性由大到小依次为针阔混交林带、农田带、寒温性针叶林带;不同海拔条件下物种多样性大小顺序为1 000~1 500 m、500 m以下、500~1 000 m、1 500 m以上;不同月份的物种多样性大小顺序为7月、8月、6月和5月.通过对五岔沟不同月份的物种多样性进行单独分析,发现该样点物种多样性情况与保护区整体不同,多样性大小顺序为7月、6月、8月.对不同的环境因素进行β多样性分析,发现蛾类昆虫的多样性受生境影响最大.综上,雾灵山蛾类昆虫的物种多样性和均匀度在各因素下差别不大,表明保护区整体环境良好,受人为干扰影响较小.
雾灵山;物种组成;多样性指数;Pielou均匀度指数;Margalef丰富度指数
雾灵山自然保护区位于河北省承德市兴隆县境内,居燕山山脉中段,地理坐标为40°29′~40°38′N,117°17′~117°35′E.该山海拔起伏较大,主峰歪桃峰高2 118 m,至连易寨一带仅为450 m,大部分山峰的海拔均在1 600 m以上.保护区总面积14 310.2 hm2,森林覆盖率达到80.3%.雾灵山属暖温带湿润大陆性季风气候区,年平均气温在7.6 ℃,5-10月日平均气温稳定超过10℃,年平均降水量在763 mm[1].雾灵山地处暖温带落叶阔叶林带和温带针阔混交林带的过渡带,同时也是动物地理区划上中日界和古北界的交界处[2],而在中国动物地理区划上处在蒙新区与华北区的交界处[3].这种复杂的地理环境和生境类型,造就了雾灵山植被类型多样化、生态系统类型繁多、生物多样性丰富的现状,使得雾灵山被广大学者誉为“物种基因宝库”.
雾灵山生物物种多样性研究始于20世纪30年代,日本植物学家中井猛之、本田正次、北川正夫对该区的植物进行了考察并于1936年总结出版《第一次满蒙学术调查研究报告》[4],至今已有80年的历史,但绝大多数研究都集中在植被与脊椎动物上[5],而对于生物界所占比重最大、种类最为丰富的昆虫来说,仅张培毅的《雾灵山昆虫生态图鉴》记载的昆虫较为全面[6],共收录雾灵山昆虫837种.对于观赏价值、经济价值极高的鳞翅目昆虫,仅牛浩和梁文琴[7]进行过整理,列举了雾灵山鳞翅目昆虫346种;鳞翅目中比例最大的夜蛾类,相关研究仅见迟美妍等[8]记录夜蛾种类44种.按照生物物种多样性比例来估算,这些研究对于雾灵山蛾类的物种统计还远远不够,物种本底可谓完全不清楚,并且对物种多样性的分析也还是空白.这对于一个国家自然保护区而言,无疑是巨大的遗憾.
有鉴于此,河北大学任国栋教授团队应河北省林业厅森防总站和雾灵山国家级自然保护区管理局邀请,于2015年5-9月对雾灵山进行了大规模昆虫物种多样性考察,以求摸清雾灵山昆虫物种资源本底,推进对保护区昆虫资源保护工作的开展.本文选取雾灵山蛾类为研究对象,在完善该地区的蛾类物种名录基础上,对蛾类种类与周围环境的关系进行分析,通过多样性指数之间的比较,得到对蛾类昆虫影响最大的环境因素,从而对保护区蛾类的防治和保护提出建议.
1 材料与方法
1.1 材料来源
研究标本全部于雾灵山自然保护区采集所得,现保存于河北大学博物馆.
1.2 研究方法
1.2.1 标本采集
依照保护区不同海拔高度和不同生境,共设置7个采集样点,于2015年5—9月间在这些样点进行周期性采集,详见表1和图1,其中,将代表针阔混交林植被带的五岔沟作为驻点样地,从6月1日开始,每周灯诱1次,其他6个样点随机采集.

表1 雾灵山保护区7个灯诱样点采集信息

图1 雾灵山保护区7个灯诱样点分布
采集方法为夜间使用450 W高压汞灯、2 m×2.5 m的白色幕布对蛾类进行灯光诱集,收集幕布周围5 m×5 m内的所有蛾类,时间为从晚20点至次日凌晨5点.
1.2.2 标本鉴定
标本鉴定主要参考文献[9-26]进行.
1.2.3 数据分析
数据分析采用的指数有[27]
优势度指数D′=Nmax/Nt,其中Nmax为最多个体数,Nt为全部个体数;
Shannon-Wiener多样性指数H′=-∑PilnPi;
Margalef丰富度指数E=(S-1)/lnS;
Whittaker多样性指数βw=S/A-1[29];
其中,Pi为第i种的个体数与总个体数的比值;S为物种数;A为群落环境梯度上发现的物种的平均数.
数据分析软件采用SPSS 20.0.
2 结果
2.1 物种组成
经统计,2015年共在雾灵山保护区采集蛾类23科308属436种(表2),其中蛾类物种最丰富的是夜蛾科,共144种,约占蛾类物种总数的33.02%,占有明显优势;其次为尺蛾科,共71种,占总数的16.28%.
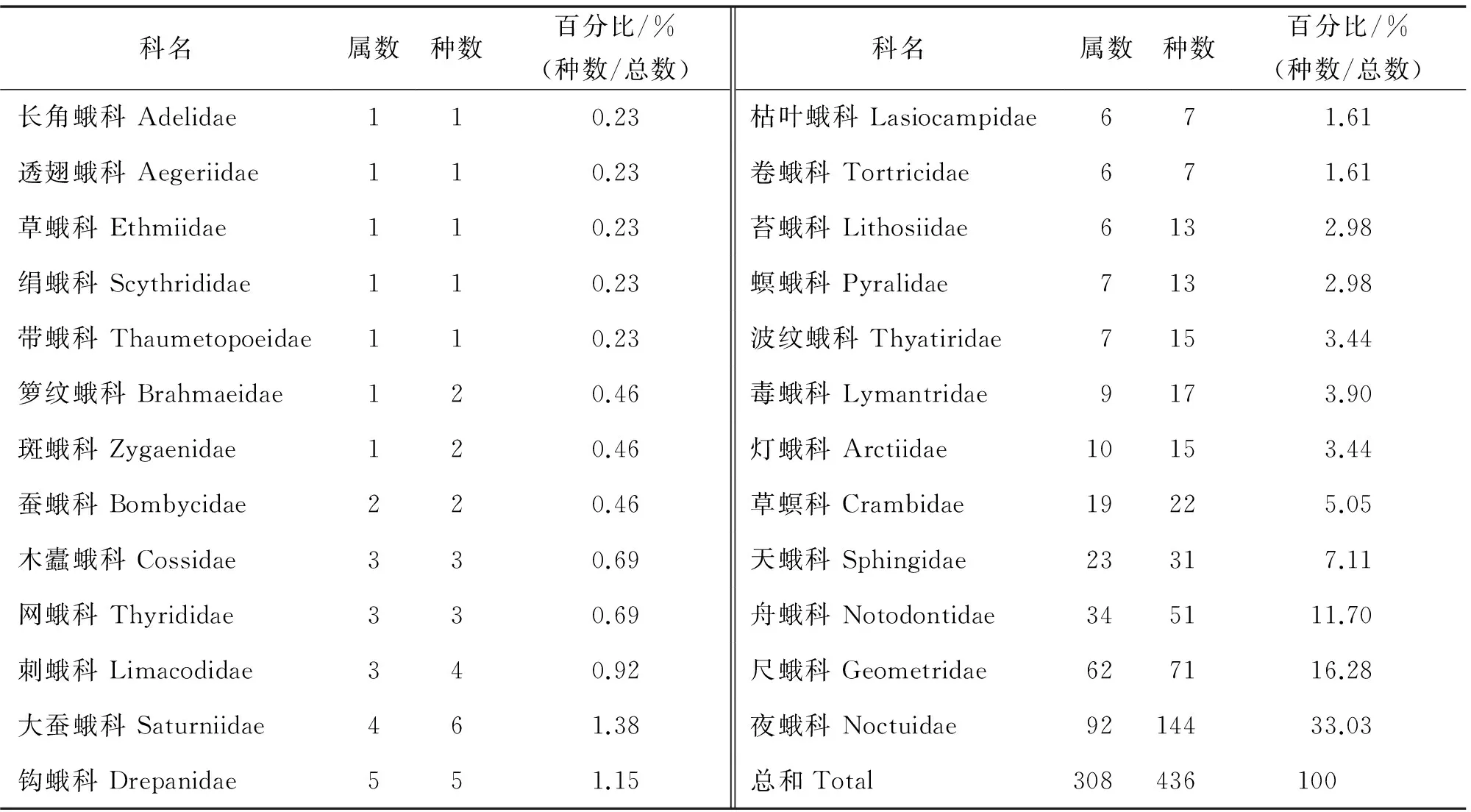
表2 雾灵山蛾类昆虫种类组成比例
2.2 雾灵山蛾类昆虫的α多样性
2.2.1 不同植被类型的蛾类昆虫群落多样性分析
雾灵山的灯诱样地涉及的植被类型有针阔混交林带、寒温性针叶林带和农田带(表1),选取在蛾类发生量较大的7、8月份诱集过的流水沟(针阔混交林带)、莲花池(寒温性针叶林带)和刘寨子(农田带)作为样点,调查并分析其蛾类物种多样性,以探究不同类型的生境对蛾类多样性的影响.
由表3可见,物种的Simpson多样性指数与Shannon-Wiener多样性指数变化趋势一致,D与H′由大到小的顺序为针阔混交林带、农田带、寒温性针叶林带.多样性指数H′最高的针阔混交林带的植被类型丰富,主要有天然油松林、少量核桃楸林和大叶白蜡林等,存在一定的人为干扰,除森林植被外还存在一些低山落叶阔叶灌丛、灌草丛等[1],致使取食此类植物的蛾类幼虫出现集中的现象.

表3 不同生境中蛾类昆虫多样性指数
Pielou均匀度指数反映了群落空间分布均匀程度,指数越大群落空间分布越均匀.由表3可见,不同生境下的Pielou均匀度指数之间差异较小,其中寒温带针叶林带的多样性指数最低但是均匀度为最高,而多样性指数最高的针阔混交林带的均匀度指数低于另两者,可见在生境的比较上多样性指数和均匀度呈反比.
Margalef丰富度指数分析中农田带的丰富度指数最高,寒温性针叶林带次之,针阔混交林带的丰富度指数最低,这与之前多样性指数与均匀度指数的变化趋势有所不同.
2.2.2 不同海拔高度蛾类昆虫群落多样性分析
随着海拔高度的升降,温度、湿度和植被类型等因素也随之发生变化,进而对蛾类昆虫的物种多样性产生影响.
将主要采集点以垂直500 m为间隔,分为500 m以下、500~1 000 m、1 000~1 500 m和1 500 m以上4个梯度(表4).比较各海拔梯度之间的多样性指数的顺序,并进行分析,由表4可见,物种多样性指数在1 000~1 500 m处具1明显峰值,而在1 500 m以上多样性指数最低,且1 500 m以上的多样性指数远小于其他海拔梯度.

表4 不同海拔高度蛾类昆虫多样性指数
比较不同海拔的Pielou均匀度指数,发现1 000~1 500 m的均匀度最高,群落分布更为均匀,而1 500 m以上的均匀度最低,可见不同海拔的Pielou均匀度指数与多样性指数呈正比.
1 000~1 500 m处的Margalef丰富度指数也最高,海拔低于1 500 m的栏地丰富度指数随海拔的增加依次升高,但1 500 m以上丰富度指数最低.
2.2.3 不同月份蛾类昆虫群落多样性动态变化分析
随着时间的变化,物种的生活周期及植物种类也发生了变化,蛾类昆虫群落多样性也随之发生变迁.对雾灵山蛾类昆虫及其发生期进行统计分析,由表5可知,不同月份蛾类的Simpson多样性指数和Shannon-Wiener多样性指数从高到低为7月、8月、6月和5月,可以看出随着夏季气温的升高,蛾类的物种多样性快速上升;但5月份的Pielou均匀度指数最高,8月份的Pielou均匀度指数最低,这与不同月份的多样性指数不呈线性相关.Margalef丰富度指数在7月份最高,5月最低.
以科级阶元进行分析,5月份以舟蛾科的种类最为丰富;6—9月则以夜蛾科的种类最多,其中8—9月夜蛾科的种数和个体数量都呈现出大幅度增长;7月份舟蛾科和尺蛾科的物种种数与夜蛾科相似(表6);保护区整体与五岔沟的7月多样性指数均为月份中最高.

表5 不同月份的蛾类昆虫多样性指数

表6 雾灵山5-8月灯下蛾类数量分布
以五岔沟为例,对6-9月各月份灯下蛾类的优势种进行统计,结果见表7,其中紫光盾天蛾的活动时间最长,从5月持续到8月.对该地单独进行多样性指数分析(表8),将结果与整个保护区进行对比,发现保护区6月份多样性小于8月份,而五岔沟6月份的多样性大于8月份,指数的差异也更明显.五岔沟均匀度指数明显大于大生境下的均匀度指数,且由大到小依次为6月、8月和7月,与多样性指数也不呈线性相关.

表7 五岔沟6-9月灯下蛾类优势种分布
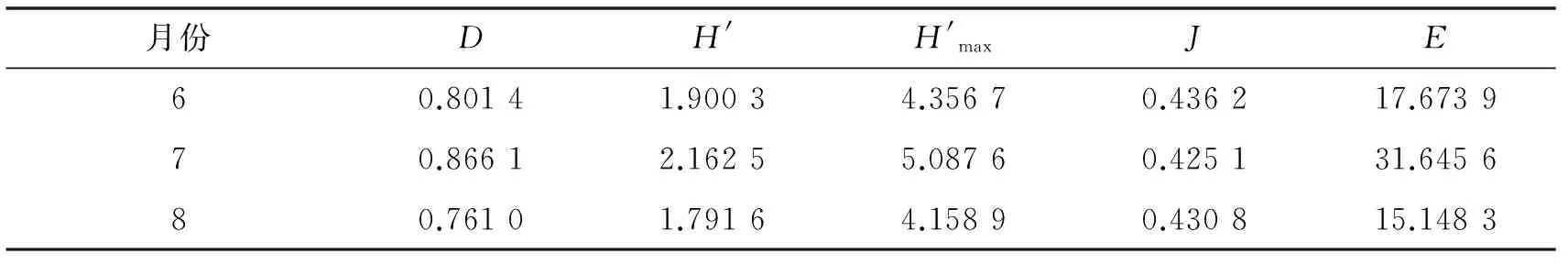
表8 五岔沟不同月份灯下蛾类昆虫多样性指数
2.3 雾灵山蛾类昆虫群落的β多样性分析
β多样性为沿着环境梯度的变化物种替代的程度,不同群落或某环境梯度上不同点之间的共有种越少,β多样性越大[30].本文使用Whittaker多样性指数对雾灵山蛾类群落组成沿环境梯度上的变化进行计算.
沿生境梯度βw=293/((47+133+146)/3)-1=1.696 5,
沿海拔梯度βw=375/((200+226+235+46)/4)-1=1.121 6,
沿月份梯度βw=371/((41+168+228+191)/4)-1=1.363 1.
由结果可知,保护区β多样性指数的大小顺序从高到低依次为沿生境梯度、沿月份梯度、沿海拔梯度,可见对雾灵山蛾类物种多样性影响最大的因素是生境类型,其次为月份,海拔影响最小.
3 讨论
从现有标本来看,雾灵山蛾类物种数量最多的是夜蛾科,物种数量较少的科分别是透翅蛾科、绢蛾科、带蛾科和草蛾科,分析其主要原因:一是这些种类多为白天活动,较难捕捉,二是小蛾类标本保存与鉴定比较困难.综上,雾灵山保护区的蛾类物种多样性研究还有很大的探究空间.在分析雾灵山保护区蛾类物种α多样性时发现,在分别使用Simpson多样性指数和Shannon-Wiener多样性指数时,两者在描述多样性的变化趋势时较为一致.一个样地多样性的大小常由多因素影响,如位于高海拔的寒温性针叶林带多样性小,是因为其海拔在1 500 m以上,气温较低,能在其环境中生活的植物种类较为单一,因此以植物为取食对象的蛾类的物种种类更少.
在对海拔因素的影响进行分析时发现寒温性针叶林带物种多样性指数与另外2个植被带的差别十分显著,可能是由于寒温性针叶林的物种组成主要为华北落叶松、硕桦等 ,植被类型较为单一造成的.在对海拔因素进行分析时发现海拔梯度与多样性指数之间并不呈线性关系.
1 000~1 500 m处多样性指数最高可能由于该海拔区间是保护区的核心区,样点所在的生境为针阔混交林带,地区植被类型复杂,所以蛾类的种类最为丰富.多样性指数方面,在6月与8月时,五岔沟的多样性指数小于整个保护区,在7月时,五岔沟的多样性超过了整体.比较后发现,五岔沟的蛾类发生时间更为集中,发生量也更大.
雾灵山保护区的均匀度分析上各因素的差别不明显,五岔沟位于海拔尺度中均匀度最高的1 000~1 500 m,在对其进行月份的均匀度分析时发现其与全保护区月份的均匀度相比更高,说明小的生境物种分布相比大尺度更为均匀.
比较Simpson多样性指数、Shannon-Wiener多样性指数和Pielou均匀度指数在植被、海拔、月份因素的变化曲线,多样性指数与均匀度指数在植被、海拔与月份3个因素的比较中出现了正相关、负相关和不相关(图2),证明多样性指数与均匀度指数间不一致.刘文萍和邓合黎[31]同样发现不同生境的蝶类群落多样性指数与均匀度不一致,并据此提出种-多度关系表现为生态位优先占领假说.而尤平等[32]对北大港湿地蛾类的研究也得到了相同的结论.
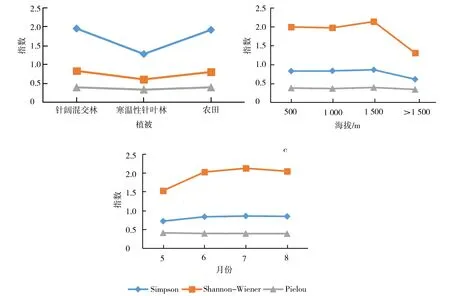
图2 群落多样性指数与均匀度指数在不同划分尺度中的动态
雾灵山保护区蛾类β多样性的大小顺序从高到低分别为沿生境梯度、沿月份梯度、沿海拔梯度.在对不同种群的研究中结果也是相似的[33-34],但也有文献表明,生境中树木种类的多样性比整体植被对物种多样性的影响更大[35],Zou等[36]也支持这个观点.对雾灵山蛾类影响最大的因素也是生境,其次为月份,影响最小的是海拔,而海拔和月份是通过改变植物群落的外貌及生境中的小环境如温度、湿度等间接影响蛾类昆虫的发生及分布.
通过对以上多样性的研究可以发现,雾灵山的蛾类多样性较为平均,种类丰富,雾灵山地区蛾类昆虫不存在明显的优势种,均匀度也较为平均,表明现在雾灵山的生态环境较为稳定,自雾灵山1984年建立自然保护区以来,生境保护良好,核心区多为天然林,植被种类较多,为蛾类昆虫的繁育生长提供了一个十分多样化的环境,而农田带的多样性与针阔混交林带差别不大,说明在雾灵山地区人为因素对蛾类昆虫的多样性影响较小.
致谢:本研究由河北省林业厅林业有害生物普查项目资助.雾灵山国家级自然保护区管理局在采集过程中提供了人力、物力及交通等方面的支持.本文多样性指数分析部分得到河北大学生命科学学院徐海云博士的指点和帮助.
[1] 王德艺,李东义,冯学全.暖温带森林生态系统[M].北京:中国林业出版社,2003.
[2] HOLT B G,LESSARD J P,BORREGAARD M K,et al.An update of Wallace’s zoogeographic regions of the world[J].Science,2013,339(6115):74-78.DOI:10.1126/science.1228282.
[3] 张荣祖.中国动物地理[M].北京:科学出版社,1999.
[4] 满蒙学术调查研究团.第一次满蒙学术调查研究报告[M].东京:早稻田大学理学部,1936.
[5] 李春秋,吴跃峰,武明录,等.雾灵山、小五台山自然保护区陆生脊椎动物研究[M].北京:中国科学技术出版社,1996.
[6] 张培毅.雾灵山昆虫生态图鉴[M].哈尔滨:东北林业大学出版社,2013.
[7] 牛浩,梁文琴.雾灵山自然保护区鳞翅目昆虫调查研究[J].安徽农业科学,2012,40(4):2049-2053.DOI:10.13989/j.cnki.0517-6611.2012.04.006 NIU H,LIANG W Q.Investigation of order Lepidoptera insects in Wulingshan Nature Reserve[J].Journal of Anhui Agricultural Science,2012,40(4):2049-2053.DOI:10.13989/j.cnki.0517-6611.2012.04.006
[8] 迟美妍,韩辉林,高强,等.以雾灵山夜蛾科为材料评估进化模型对DNA 条码分类的影响[J].昆虫学报,2012,55(10):1193-1204. CHI M Y,HAN H L,GAO Q,et a.Effects of different evolution models on DNA barcoding evaluated with the Noctuidae from Wuling Mountain,Hebei,northern China[J].Acta Entomologica Sinica,2012,55(10):1193-1204.
[9] 中国科学院动物研究所.中国蛾类图鉴Ⅰ[M].北京:科学出版社,1981.
[10] 中国科学院动物研究所.中国蛾类图鉴Ⅱ[M].北京:科学出版社,1982.
[11] 中国科学院动物研究所.中国蛾类图鉴Ⅲ[M].北京:科学出版社,1982.
[12] 中国科学院动物研究所.中国蛾类图鉴Ⅳ[M].北京:科学出版社,1983.
[13] 朱弘复,王林瑶.中国动物志·昆虫纲第三卷[M].北京:科学出版社,1991.
[14] 朱弘复,王林瑶.中国动物志·昆虫纲第五卷[M].北京:科学出版社,1996.
[15] 朱弘复,王林瑶.中国动物志·昆虫纲第十一卷[M].北京:科学出版社,1997.
[16] 薛大勇,朱弘复.中国动物志·昆虫纲第十五卷[M].北京:科学出版社,1999.
[17] 陈一心.中国动物志·昆虫纲第十六卷[M].北京:科学出版社,1999.
[18] 方承莱.中国动物志·昆虫纲第十九卷[M].北京:科学出版社,2000.
[19] 赵仲苓.中国动物志·昆虫纲第三十卷[M].北京:科学出版社,2003.
[20] 武春生,方承莱.中国动物志·昆虫纲第三十一卷[M].北京:科学出版社,2003.
[21] 赵仲苓.中国动物志·昆虫纲第三十六卷[M].北京:科学出版社,2004.
[22] 刘友樵,武春生.中国动物志·昆虫纲第四十七卷[M].北京:科学出版社,2006.
[23] 韩红香,薛大勇.中国动物志·昆虫纲第五十四卷[M].北京:科学出版社,2011.
[24] 刘友樵,白九维.中国经济昆虫志第十一册[M].北京:科学出版社,1977.
[25] 李后魂,王淑霞.河北动物志鳞翅目小蛾类[M].北京:中国农业科学技术出版社,2009.
[26] 虞国跃.北京蛾类图谱[M].北京:科学出版社,2014.
[27] 马克平.生物群落多样性的测度方法:Ⅰα多样性的测度方法(上)[J].生物多样性,1994,2(3),162-168.
[28] PIELOU E C.The measurement of diversity in different types of biological collections[J].Journal of Theoretical Biology.1966:131-144.DOI:10.1016/0022-5193(66)90013-0.
[29] WHITTAKER R H.Evolution and measurement of species diversity[J].Taxon,1972,21(2):213-251.DOI:10.2307/1218190.
[30] 赵志模,郭依泉.群落生态学原理与方法[M].重庆:科学技术出版社重庆分社,1990.
[31] 刘文萍,邓合黎.木里蝶类多样性的研究[J].生态学报,1997,17(3):266-271. LIU W P,DENG H L.The butterfly diversities in muli,sichuan province[J].Acta Ecologica Sinica,1997,17(3):266-271.
[32] 尤平,李后魂,王淑霞.天津北大港湿地自然保护区蛾类的多样性[J].生态学报,2006,26(4):999-1004. YOU P,LI H H,WANG S X.The diversity of the moth community in the north dagang wetland nature reserve,Tianjin[J].Acta Ecologica Sinica,2006,26(4):999-1004.
[33] 于晓东,周红章,罗天宏.云南西北部地区地表甲虫的物种多样性[J].动物学研究,2001,22(6):454-460.
[34] 王松,鲍方印,梅百茂,等.安徽鹞落坪国家级自然保护区蝶类的垂直分布及其群落多样性[J].应用生态学报,2009,20(9):2262-2270.DOI:10.13287 / j.1001-9332 2009.0354. WANG S,BAO F Y,MEI B M,et al.Vertical distribution and community diversity of butterflies in Yaoluoping National Nature Reserve,Anhui,China[J].Chinese Journal of Applied Ecology,2009,20(9):2262-2270.DOI:10.13287 / j.1001-9332 2009.0354.
[35] BREHM G,HOMEIER J,FIEDLER K.Beta diversity of geometrid moths (Lepidoptera:Geometridae) in an Andean montane rainforest[J].Diversity and Distributions,2003,9:351-366.DOI:10.1046/j.1472-4642.2003.00023.x.
[36] ZOU Y,SANG WG,BAI F,et al.Relationships between Plant Diversity and the Abundance and α-Diversity of Predatory Ground Beetles (Coleoptera:Carabidae) in a Mature Asian Temperate Forest Ecosystem[J].Plos One,2013,8(12):1-7.DOI:10.1371/journal.pone.0082792.
(责任编辑:赵藏赏)
Constitution and biodiversity of moths in the forest at Wuling Mountain of Hebei Province,China
TANG Shenyan1,PAN Zhao1,REN Weihong2,XIANG Yafei3,MENG Xiangpu3,WANG Yuanyuan3,REN Guodong1
(1.The Key Laboratory of Zoological Systematics and Application,College of Life Sciences,Hebei University,Baoding 071002,China;2.The Quarantine Department,The Forest Pests Control and Quarantine Inspection Bureau of Hebei Province,Shijiazhuang 050000,China;3.The Resources Division, Wulingshan National Nature Reserve,Chengde 067300,China)
There are 436 moth species belonging to 23 families identified from the specimens which were collected through field biodiversity investigation of the forest insects at seven locations of the Wulingshan National Nature Reserve of Hebei Province,China from May to September,2015.Eleven widely distributed families were chosen to analyze biodiversity using Simpson index,Shannon-Wiener index,Pielou index,Whittaker index and Margalef index.The results showed that the order of different factors from the highest to lowest are:1) the habitats range from mixed broadleaf-conifer forest,farmland and coniferous forest;2) elevational range from 1 000-1 500 m,under 500 m,500-1 000 m and above 1 500 m;3) time range from July,August,June and May.Focus on Wuchagou only,the species diversity is most in July,then is June and August,different from the result of the whole reserve.Pielou index of Wuchagou is the highest than that of the whole reserve,this result proved that the species distribution equilibrium of microhabitat is higher than that of macrohabitat.To analyzeβdiversity of different environmental factors,we considered that the habitat is the main factor on component of diversity of moths in the reserve,the next is month,and the minimally influencing factor is altitudinal distribution.Focus on family taxa,Notodontidae is the largest species family in May,Noctuidae is the largest from June to September,and numbers of both species and individuals of Nocuidae rapidly increase from August to September.In July,the species amount of Notodontidae and Geometridae are close to that of Noctuidae.In conclusion,Wuling Mountain is a moth species-rich region,there is not a conspicuous moth sociales at Wuling Mountain which means ecological environment at Wuling Mountain is stable,the effect of man-made interference is not serious.
Wuling Mountain;species constitution;diversity index;Pielou index;Margalef index
10.3969/j.issn.1000-1565.2017.01.010
2016-09-25
河北省重点实验室运行补助费资助项目(14967611D)
唐慎言 (1992—),女,湖南衡阳人,河北大学在读硕士研究生. E-mail:tangshenyan913@163.com
任国栋 (1955—),男,宁夏中卫人,河北大学教授,博士生导师,主要从事昆虫系统及进化学研究. E-mail:gdren@hbu.edu.cn
O175.12
A
1000-1565(2017)01-0062-09
