两种地被植物对干旱胁迫的生理生化响应
2017-03-08孙虹豆王贝贝张芝蕊张雪薇骆建霞
孙虹豆,王贝贝,张芝蕊,张雪薇,骆建霞
我国北方地区水资源匮乏,干旱少雨、土壤干燥,因此研究植物的抗旱性对园林绿化具有重要意义。匍枝委陵菜(Potentilla flagellaris)和荆芥(Nepeta cataria)是近年来在国内绿化实践中逐渐被广泛应用的新型地被植物,具有适应能力强、对土壤要求不严格、绿叶期长、生长快、繁殖能力较强等优点,可广泛地应用于城市的园林绿化。关于园林植物耐旱性研究[1-3]及委陵菜[4-5]的研究多有报道,但尚未见到匍枝委陵菜和荆芥的耐旱性比较的研究报道。本试验通过对匍枝委陵菜和荆芥在干旱胁迫下生理生化指标的测定,旨在了解两种植物的耐旱能力及差异,为其在园林绿化中的应用和制定苗木栽培管理措施提供理论指导。
1 材料和方法
1.1 材料
培育匍枝委陵菜和荆芥扦插苗,幼苗株高8~10cm时,移栽到盛有1800g培养土(m园土︰m珍珠岩=4︰1)的花盆中,进行缓苗生长,当植株高度生长至15~20 cm左右时,进行干旱胁迫处理。
1.2 方法
采用盆栽试验法,设置轻度胁迫(土壤相对含水量(SRW)为50%)、中度胁迫(SRW为35%~40%)、重度胁迫(SRW为25%~30%)3个干旱胁迫处理,以正常浇水为对照(SRW保持在60%~80%),称重法监测土壤含水量。试验开始时将每盆浇透水,之后不浇水让土壤水分持续降低,当降至设置的干旱胁迫处理时,继续胁迫24 h后取样进行各项生理生化指标的测定。采用随机区组设计,重复4次,每个处理至少25株。
1.3 测定指标及方法
选取新梢中部有代表性的成熟叶片进行生理生化指标测定[6]。采用蒽酮法测定可溶性糖含量;磺基水杨酸提取茚三酮显色法测定脯氨酸含量;考马斯亮蓝法测定可溶性蛋白含量;氯化硝基四氮唑蓝(NBT)法测定超氧化物歧化酶(SOD)活性;愈创木酚显色法测定过氧化物酶(POD)的活性;氯化三苯基四氮唑(TTC)法测定根系活力;硫代巴比妥酸(TBA)法测定丙二醛(MDA)含量。
1.4 数据处理
利用SPSS17.0统计软件对试验数据进行方差分析,SSR法进行多重比较。
2 结果与分析
2.1 参试植物的渗透调节物质在干旱胁迫下的变化
对干旱胁迫下两种植物的可溶性糖、脯氨酸及可溶性蛋白含量进行测定,结果见表1。
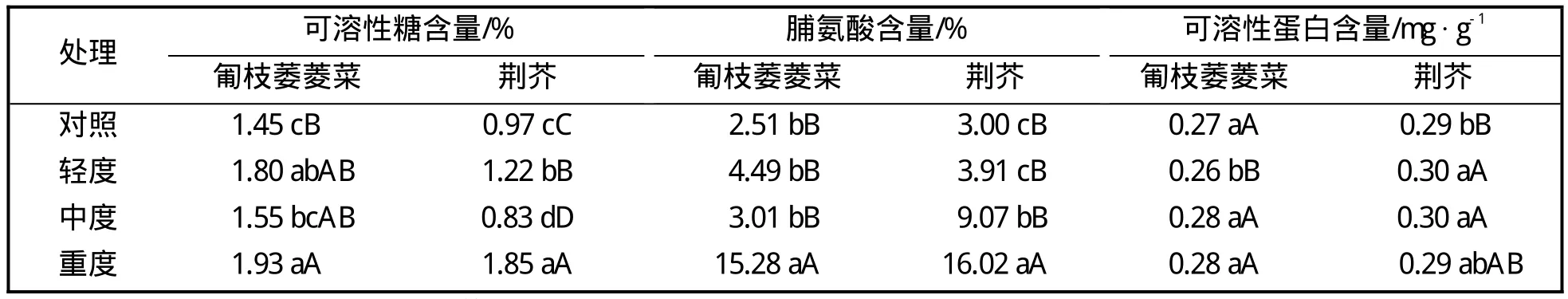
表1 参试植物在干旱胁迫下的渗透调节物质的变化%
由表1表明,两种地被植物的可溶性糖含量随着干旱胁迫程度的加深基本呈上升趋势,在干旱胁迫至中度时,匍枝委陵菜保持在对照水平,荆芥则低于对照;匍枝委陵菜的脯氨酸含量在轻度和中度干旱胁迫时均与对照差异不显著,重度干旱时极显著性上升,而荆芥脯氨酸含量则在轻度胁迫时保持在对照水平,中度时显著上升。两种地被植物的可溶性蛋白含量在干旱胁迫下的变化幅度较小。
2.2 参试植物叶片中的抗氧化酶活性在干旱胁迫下的变化
对干旱胁迫下两种植物的超氧化物歧化酶(SOD)和过氧化物酶(POD)活性进行测定,结果见表2。
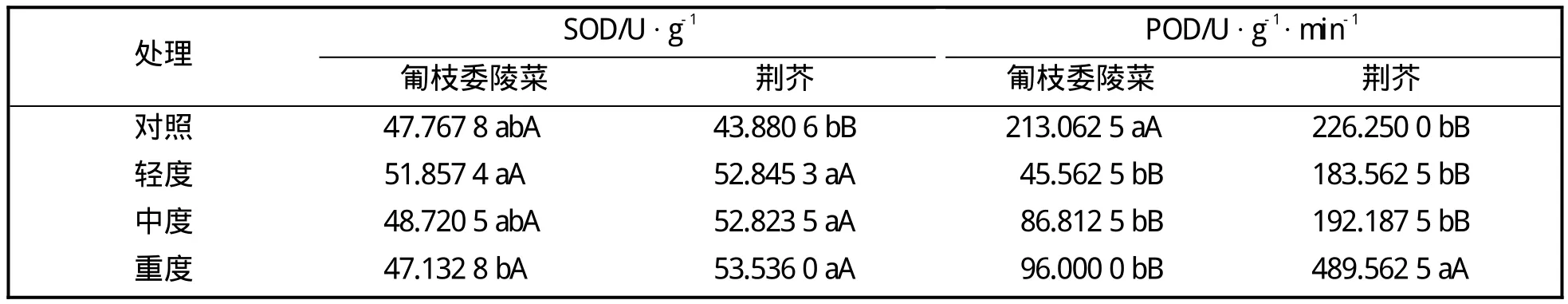
表2 干旱胁迫下两种植物SOD和POD活性的变化
表2表明,匍枝委陵菜的SOD活性在干旱胁迫处理下变化很小,POD活性则在干旱胁迫下极显著下降;荆芥的SOD活性在干旱胁迫时极显著上升,POD活性在轻、中度胁迫时保持在对照水平,在重度胁迫时极显著上升。上述结果表明,两种地被植物的SOD和POD活性在干旱胁迫下表现不同,可能是其对抵御干旱做出的响应有差异。
2.3 参试植物的根系活力、丙二醛含量对干旱胁迫的响应
对干旱胁迫下两种植物的根系活力、丙二醛含量进行测定,结果见表3。
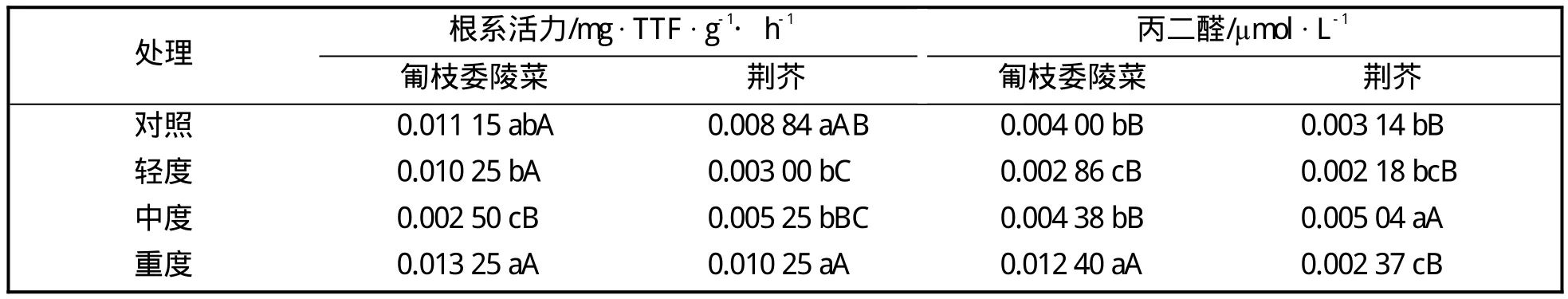
表3 匍枝委陵菜和荆芥的根系活力和丙二醛含量在干旱处理下的变化
表3表明,两种地被植物的根系活力均随干旱胁迫程度的加深呈先降后升趋势,在重度胁迫时均能恢复至对照水平。荆芥在轻度胁迫时、匍枝委陵菜在中度胁迫时才极显著下降,表明匍枝委陵菜在干旱胁迫下比荆芥能保持更高的根系活力。匍枝委陵菜的丙二醛含量在轻、中度胁迫时较对照无极显著差异,在重度胁迫时极显著增加,表明此时细胞受到较严重损害,而荆芥的丙二醛含量则在中度干旱胁迫时极显著高于对照。
3 讨论与结论
渗透调节是植物忍耐水分亏缺的重要生理机制,植物在感受到水分亏缺时,通过调节可溶性糖和可溶性蛋白等渗透调节物质抵抗干旱,它们积累的多少与植物抗逆性有关,可作为抗逆性筛选的指标[7]。本试验中,匍枝委陵菜和荆芥的可溶性糖和可溶性蛋白含量随着干旱程度的加深呈上升趋势,表明其能通过积累渗透调节物质来提高植株的吸水保水能力以抵御干旱逆境,这与试验中测定的根系活力变化趋势基本吻合。中度干旱胁迫时,匍枝委陵菜的可溶性糖和可溶性蛋白含量均能恢复到对照水平,能通过自身生理调节来抵御干旱,这与刘艳等[8]对甘草的研究结果一致。
脯氨酸作为胞质渗透调节剂,在植物遭受干旱后会积累,其积累的多少与植物抗逆性有关。研究认为,脯氮酸含量的积累是植物正面抵御干旱的结果,积累的数量与抗旱性呈正相关[9]。也有研究认为,脯氨酸含量的变化是干旱胁迫下的一种响应,其积累是植物受伤害的结果[10-12]。本试验中,随干旱胁迫程度的加深,参试植物叶片游离脯氨酸含量均有所增加,而荆芥脯氨酸积累的幅度更大,积累速度更快,对干旱更敏感,此结果支持卢少云等[11]、刘娥娥等[12]的观点,即在同一胁迫条件下,耐旱性强的品种脯氨酸积累较少,而敏感品种脯氨酸积累则较多。
干旱胁迫会造成植物细胞膜系统破坏,而植物体内过剩自由基的毒害是造成膜系统损伤的重要原因,严重时可导致植物细胞死亡[6]。超氧化物歧化酶(SOD)和过氧化物酶(POD)是膜脂过氧化防御系统的主要保护酶,超氧化物歧化酶能催化活性氧发生歧化反应,产生无毒分子氧和过氧化氢,过氧化物酶可去除生理系统中的过氧化氢,从而使植物免遭伤害[7]。本试验中,在干旱胁迫处理下,匍枝委陵菜SOD活性未发生显著变化,POD活性则极显著下降,这与蔷薇猪毛菜和粗枝猪毛菜在干旱胁迫下的SOD活性变化趋势一致[13]。荆芥的SOD活性在干旱胁迫时极显著增加,POD活性在轻、中度胁迫时保持在对照水平,未发生显著变化,在重度干旱胁迫时极显著增加,支持了时丽冉等[14]的研究结果。两种植物表现出的SOD和POD活性对干旱胁迫的响应差异,可能与植物的内部结构、调节方式、响应机制不同有关。荆芥的抗氧化酶活性在干旱胁迫下的增加幅度较匍枝委陵菜大也说明了荆芥对干旱胁迫更加敏感。
根系作为植物吸收水分和盐类的器官,同时参与多种物质的同化、转化和合成。根系活力反映了植物适应逆境、抵抗干旱环境的能力[15]。两种地被植物的根系活力随干旱程度的加深呈先降后升趋势,匍枝委陵菜比荆芥保持更高的根系活力,这与本试验中两种植物在干旱胁迫期间和复水解除干旱胁迫后植株的形态表现结果相吻合。在轻度干旱胁迫时匍枝委陵菜叶片出现萎蔫现象,而荆芥叶片萎蔫现象不明显,这可能与其表面覆绒毛、水分散失慢致萎蔫较慢有关,但在中度和重度干旱胁迫下两种植物萎蔫程度均逐渐增大。复水后,两种植物均可恢复正常,但匍枝委陵菜因保持较高的根系活力,故恢复正常表现所需时间比荆芥短。丙二醛(MDA)含量是膜脂过氧化过程中的分解产物,反映细胞膜的受害程度,进而反映植物的抗逆性[6]。有研究表明,丙二醛的含量随干旱胁迫时间的延长出现先增后降的趋势,说明植物组织在干旱胁迫至一定程度下遭到破坏,而这个峰值出现的越晚,其忍耐水分胁迫的能力越强[16]。本试验中,两种植物的丙二醛含量均随干旱胁迫的加深而增加,荆芥在中度干旱胁迫时达到峰值,而匍枝委陵菜则在重度干旱胁迫时才出现峰值。
综合各测试指标及对植株的观察结果认为,两种植物均有较强的耐旱能力,匍枝委陵菜的耐旱能力更强。
[1]韩玉林,孙桂弟,黄苏珍,等.干旱胁迫对鸢尾属5种观赏地被植物部分生理代谢的影响[J].北方园艺,2006(6):96-98.
[2]包卓,孟祥英,张晓松,等.干旱胁迫对5种园林绿化植物生理生化的影响[J].经济林研究,2010,28(1):46-50.
[3]Egilla J N,Davies F T,Drew M C.Effect of potassium on drought resistance ofHibiscus rosa-sinensiscv.Leprechaun:Plant growth,leaf macro-and micronutrient content and root longevity[J].Plant and Soil,2001,229(2):213-224.
[4]赵婵璞,王卫军,刘冬云,等.水分胁迫对匍枝委陵菜形态及生理特性的影响[J].河北农业大学学报,2013,26(5):34-38.
[5]李高,杜玉玲,赵宏斌.几种地被植物的抗逆性比较[J].内蒙古农业大学学报,2011,32(2):42-47.
[6]张治安,张美善,蔚荣海.植株生理学试验指导[M].北京:中国农业出版社,2004.
[7]鲁松.干旱胁迫对植物生长及其生理的影响[J].江苏林业科技,2012,39(4):51-54.
[8]刘艳,陈贵林,蔡贵芬,等.干旱胁迫对甘草幼苗生长和渗透调节物质含量的影响[J].西北植物学报,2011,31(11):2259-2264.
[9]许祥明,叶和春,李国凤.脯氨酸代谢与植物抗渗透胁迫的研究进展[J].植物学通报,2000,17(6):536-542.
[10]史燕山,骆建霞,王煦,等.5种草本地被植物的抗旱性研究[J].西北农林科技大学学报,2005,33(5):130-134.
[11]卢少云,陈斯平,陈斯曼,等.三种暖季型草坪草在干旱条件下脯氨酸含量和抗氧化酶活性的变化[J].园艺学报,2003,30(3):303-306.
[12]刘娥娥,宗会,郭振飞,等.干旱、盐和低温胁迫对水稻幼苗脯氨酸含量的影响[J].热带亚热带植物学报,2000,8(3):235-238.
[13]闻志彬,张明理.干旱胁迫对2种光合类型C4荒漠植物叶片光合特征酶和抗氧化酶活性的影响[J].西北植物学报,2015,35(9):1815-1822.
[14]时丽冉,陈红艳,崔兴国.干旱胁迫对地被菊膜脂过氧化和抗氧化酶活性的影响[J].园林花卉,2010(9):96-98.
[15]徐兴友,张风鹃,龙茹,等.6种野生耐旱花卉幼苗叶片脱水和根系含水量与根系活力对干旱胁迫的反应[J].水土保持学报,2007,21(1):180-184.
[16]顾文毅,王得祥,刘小利.干旱胁迫对四种野生地被植物抗性生理指标的影响[J].北方园艺,2015(18):81-83.
