高CO2条件下温度和光照对铜藻吸收N、P的短期影响
2017-02-27杨安强史定刚陈少波王铁杆关万春
杨安强,史定刚,陈少波,张 鹏,王铁杆,关万春*
(1.温州医科大学 生命科学学院,浙江 温州 325035; 2.浙江省海洋水产养殖研究所,浙江 温州 325005)
高CO2条件下温度和光照对铜藻吸收N、P的短期影响
杨安强1,史定刚1,陈少波2,张 鹏2,王铁杆2,关万春1*
(1.温州医科大学 生命科学学院,浙江 温州 325035; 2.浙江省海洋水产养殖研究所,浙江 温州 325005)
探讨在2种CO2浓度(0.76和1.96 μg·L-1)条件下,温度和光照对铜藻(Sargassumhorneri)吸收N、P速率的影响。结果表明,培养液中通入空气(LC,Low CO2)的情况下,铜藻对N、P的吸收速率与时间的关系符合指数衰减方程y=a+b×e-x/c。吸收速率在培养6 h后趋于稳定,且该趋势未受到温度、光强的影响。通入高浓度CO2(HC,High CO2)后,铜藻对N、P的吸收速率也符合LC的规律,但HC处理1 h,可显著提高铜藻对营养盐的吸收速率,随后HC的促进作用逐渐消失。HC条件下,20 ℃培养的铜藻对营养盐的吸收速率最高,10 ℃培养的铜藻对营养盐N、P的吸收分别被抑制46.03%和73.93%,30 ℃培养的铜藻仅对P的吸收造成抑制,抑制率为31.46%。光照强度为70 μmol·m-2·s-1培养的铜藻营养盐的吸收速率最高;升至280 μmol·m-2·s-1后,吸收速率分别下降65.51%(N)和28.04%(P)。因此,高浓度的CO2短期处理铜藻,可显著提高铜藻对营养盐的吸收速率,该促进作用在试验开始的1 h内最显著。
CO2; 光强; 铜藻; 温度
在全球气候变化的背景下,人类活动造成的CO2浓度升高是目前海洋面临的生态环境问题之一[1]。随着化石燃料燃烧和森林砍伐的日益加剧,大气CO2浓度已由工业革命前的0.55 μg·L-1升至目前的0.76 μg·L-1左右,约高出40%,而大气中30%的CO2是由海洋通过生物泵吸收的。据IPCC数据统计,预测到2100年,大气CO2浓度将达到约1.96 μg·L-1[2]。这将导致海洋表层海水pH下降0.3~0.4个单位(7.8~7.9),造成海水酸化,从而对具有光合固碳能力的藻类产生显著影响。大型海藻对环境中无机碳的利用机制存在明显种间差异,因此大型海藻对高CO2的响应也不尽相同[3]。高CO2培养条件可促进条斑紫菜(Porphvrayezoensis)[4]、酵母状节荚藻(Lomentairaarticulata)[5]的生长,也有发现高CO2培养降低了紫菜属中的P.leucostica[6]与P.linearis[7]的生长速率。但高CO2并未显著提高石莼(Ulva)[8-9]和江蓠属(Gracilariagaditana)[10]的生长。所以,高CO2培养对大型海藻的影响仍存在较大争议,可能与不同大型海藻的种间差异和环境因子有关。有研究表明,高CO2提高了铜藻(Sargassum.horneri)的生长,且这一过程受到温度和光强的调节[11]。因此,本文针对这一现象,探讨铜藻在高CO2条件下对N、P的利用情况,并结合温度和光强来分析高CO2对铜藻吸收营养盐存在何种调节作用。
1 材料与方法
1.1 材料
铜藻经合子培养成苗后在室内水池培养成1~2 m长的藻植株。选择健康藻体,除去表面附着物,藻体截成约3 cm小段,保持每段假茎长度相近,且在中部有2~3个假叶节,置于水箱中适应培养。适应期水温(20±0.5)℃,盐度24 PSU,光照强度50 μmol·m-2·s-1,光暗周期L∶D为12∶12,每3 d更换1次培养液,适应培养6 d。培养液为过滤的天然海水,灭菌后以F/8培养基的溶解无机氮(dissolved inorganic nitrogen,DIN)和溶解无机磷(dissolved inorganic phosphorus,DIP)浓度加富海水(DIN 431.0±57.7 μmol·L-1;DIP 9.0±0.66 μmol·L-1)。藻体经饥饿处理24 h后开始试验。
1.2 方法
1.2.1 高浓度CO2气体调节
设定CO2培养箱(MGC-450BP,上海一恒)内的CO2浓度约为1.96 μg·L-1(水体pH值约为7.80),该浓度约为2100年大气中CO2的浓度[2]。将所有藻体分置于培养箱中。一组置于CO2培养箱,通入培养箱中高CO2浓度气体,定义为HC(High CO2,CO2浓度约为1.96 μg·L-1,pH为7.8左右);另一组置于普通培养箱中(GTX-430E,江南仪器),通入室外新鲜空气,定义为LC(Low CO2,CO2浓度约为0.76 μg·L-1, pH为8.2左右),通过此法可调节2种不同处理铜藻的培养液pH值,相差0.3~0.4。
1.2.2 高浓度CO2条件下温度对铜藻N、P吸收的短期影响
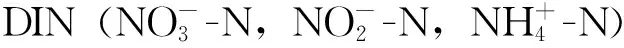
1.2.3 高浓度CO2条件下光照对铜藻N、P吸收的短期影响
选取大小均一、长势健康的铜藻,分成6组(每组重复2个,每重复鲜重为5±0.1 g),分别置于12个3 L锥形瓶中进行充气(30 mL·s-1)培养,温度20 ℃。其余试验处理为LLHC、MLHC、HLHC、LLLC、MLLC、HLLC,LL表示70 μmol·m-2·s-1低光,ML表示140 μmol·m-2·s-1中光, HL表示280 μmol·m-2·s-1高光,HC表示CO21.96 μg·L-1,LC表示CO20.76 μg·L-1。试验开始后于0、1、3、6、12 h取样,每处理分别取200 mL培养液,用于测定DIN和DIP。
1.2.4 DIN、DIP测定和计算

U=(Co-Ct)·V/(t·G)。
式中,U为营养盐吸收速率(μmol·g-1Fw·h-1),Co为试验开始时海水中营养盐含量(μmol·L-1),Ct为试验开始t时海水中营养盐含量(μmol·L-1),V为海水体积(L),t为试验时间(h),G为铜藻鲜重(g)。
1.3 数据处理与分析
采用单因素方差分析(one-way ANOVA)对数据进行分析,以P<0.05作为差异显著水平。
2 结果与分析
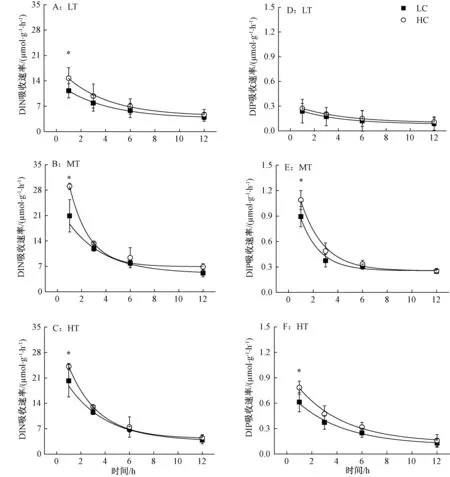
铜藻利用N和P的吸收动力学曲线符合指数衰减方程y=a+b×e-x/c(R2>0.87,a、b、c为调节参数),经过6 h基本达到稳定,且该规律不受到温度、光强和CO2浓度的影响(图1~2),但温度和光强可影响其绝对吸收速率。
2.1 温度对铜藻DIN、DIP吸收的短期影响
由图1可知,温度20 ℃时,铜藻对氮和磷的吸收最快,1 h内分别达到20.92和0.89 μmol·g-1·h-1(图1中B、E)。且提高CO2浓度则显著提升铜藻对氮和磷的吸收速率,1 h内可分别提升38.77%和22.47%。而当温度降至10 ℃或升至30 ℃后,铜藻对氮的吸收速率分别下降46.03%和3.49%,对磷的吸收速率分别下降73.93%和31.46%。这一现象在高CO2条件下也存在,铜藻对氮的吸收速率分别下降49.74%和20.82%,对磷的吸收速率分别下降74.87%和27.47%。高CO2对铜藻利用营养盐的促进作用在3 h后消失。而LT条件下,HC对吸收磷的促进作用不明显。
2.2 光强对铜藻DIN、DIP吸收的短期影响
由图2~3可知,随着光强的增加,铜藻对氮的吸收速率都存在下降趋势。在HC培养1 h、光强为LL时,对氮的吸收速率达到29.86 μmol·g-1·h-1,比ML和HL分别高出30.13%和189.59%;对磷的吸收速率达到0.88 μmol·g-1·h-1,比HL高出38.32%,而与ML相比差异不大。LC时也存在类似情况。HC处理1 h后,显著提高了铜藻吸收氮的速率,对比3种光强,分别提高55.55%(LL)、42.68%(ML)、26.27%(HL)(图2中A,B,C),对磷的吸收速率分别提高22.59%(LL),33.59%(ML)和12.84%(HL)。而HC对铜藻利用N、P的促进作用,随着培养时间的延长,经过3 h后逐渐消失,并趋于稳定。
*代表差异显著(图2同)图1 2种CO2条件下铜藻的氮(A,B,C)、磷(D,E,F)吸收速率曲线对温度的响应
3 讨论
铜藻吸收氮和磷的速率受到温度和光强的影响,且高CO2处理1 h后,除低温(10 ℃)条件下铜藻对P的吸收以外,HC对铜藻利用营养盐的促进作用极其显著,LC仅是HC的0.72~0.88倍,而随后这个比值逐渐接近于1。
温度可通过影响藻体酶的活性来影响细胞对营养盐的吸收。20 ℃时,铜藻吸收氮磷营养盐的速率最快,这是因为藻体培养温度即为20 ℃,与该藻的最适生长温度(22~24 ℃)较接近[12]。此时,细胞相关酶的活性高[13-14],所以吸收速率最高。试验设定的低温(10 ℃)抑制了光合作用暗反应相关酶的活性,减少了ATP的生成,从而导致供给N、P代谢所需的能量减少[15];而试验设定的高温(30 ℃)会造成酶和膜载体蛋白失活[16],也同样影响营养盐的代谢速率。
光照强度主要通过光合磷酸化调控ATP的转换,从而影响N、P的吸收速率,适宜的光照强度能增加海藻中的还原酶活性,促进藻体对N、P的吸收[13-14,17]。而过强的光照会使海藻光系统Ⅱ中的D1蛋白失活、裂解,从而降低光合速率,并间接导致N、P吸收速率的减慢[18]。本试验所用的铜藻培养于50 μmol·m-2·s-1的光照条件下,适应低光环境,当短期提高光照强度(140和280 μmol·m-2·s-1),会对藻体造成光抑制,从而降低藻体
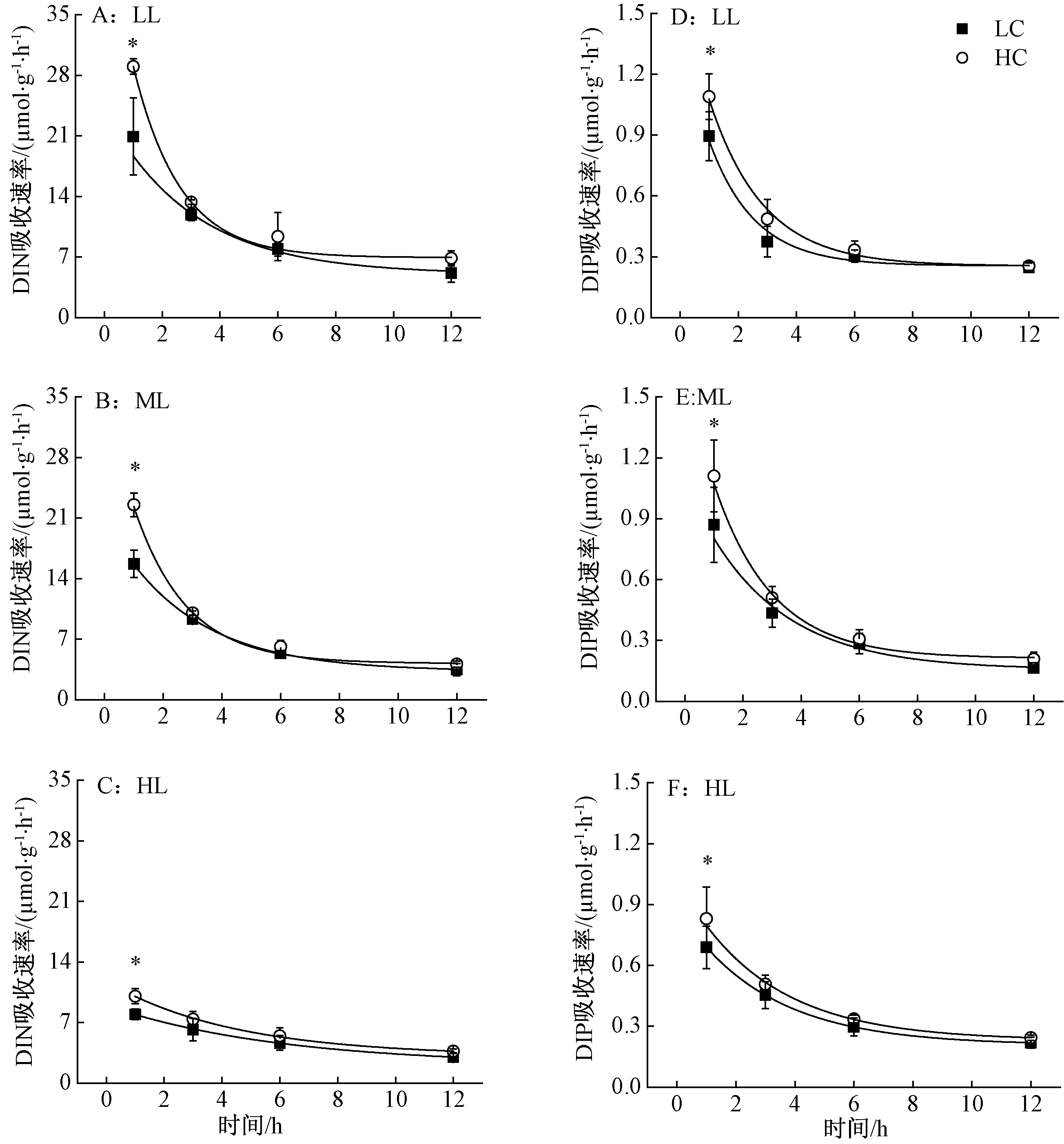
图2 2种CO2条件下铜藻的氮(A,B,C)、磷(D,E,F)吸收速率曲线对光的响应
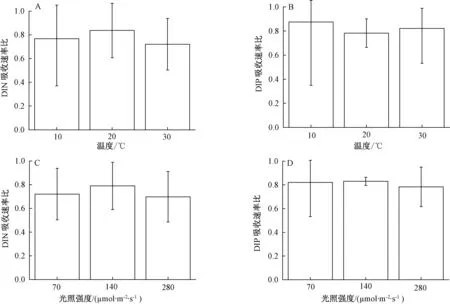
图3 高CO2条件下1 h内温度、光强对铜藻氮(A,C)、磷(B,D)吸收速率比的影响
对N、P的吸收速率。但光背景对藻体营养盐代谢的影响可以通过一定时间的光适应而消除或减弱。
短期提高CO2浓度,显著提升了铜藻对N、P的吸收速率,LC/HC<1。这可能是因为HC条件下短期内增加了藻体可利用的无机碳的缘故,光合速率加快,从而在短期内提高了营养盐代谢速率。而CO2的促进作用在培养3 h后消失,是因为短期高CO2浓度并未造成N和P代谢相关酶数量和活性变化的缘故。然而在测定的12 h内,藻体对N、P的吸收速率在培养1 h时,吸收速率最高,然后随着培养时间延长,逐渐下降,最后趋于稳定,这是因为藻体经24 h饥饿处理的缘故。处于饥饿状态的藻体对营养盐的吸收会存在被动扩散的过程,以便迅速补充细胞内部营养库的严重亏空[19]。所以,在开始的1 h可以看到吸收速率最高。而随后藻体对营养盐的吸收速率下降,并达到稳定阶段,这是由于藻体内营养库已经充盈,对营养盐的吸收速率产生了反馈调节作用[20],这种现象在龙须菜和羊栖菜的研究中也存在相似规律[13-14]。
铜藻的营养盐代谢受到温度和光强的显著调节,且短期升高CO2浓度,对铜藻的营养盐代谢也存在一定得促进作用。
[1] DONEY S C,FABRY V J,FEELY R A,et al. Ocean acidification:the other CO2problem[J]. Marine Science,2009,1:169-192.
[2] STOCKER T F,QIN D,PLANTTNER G K,et al. IPCC,2013:Climate change 2013:The physical science basis.contribution of working group I to the fifth assessment report of the intergovermrntal panel on climate change[M]//Cambridge,U K. and New York:Cambridge Univisity Press,2013:525-527.
[3] BEER S,KOCH E. Photosynthesis of marine macroalgae and seagrasses in globally changing CO2environments[J]. Marine Ecology-Progress Series,1996,141:199-204.
[4] GAO K,ARUGA Y,ASADA K,et al. Enhanced growth of the red algaPorphyrayezoensisUedain high CO2concentrations[J]. Journal of Applied Phycology,1991,3(4):355-362.
[5] KBLER J E,JOHNSTON A M,RAVEN J A. The effects reduced and elevated CO2and O2on the seaweedLomentariaarticulate[J]. Plant Cell Envir,1999,22:1303-1310.
[6] MERCADO J M,JAVIER F,GORDILIO L,et al. Effects of different leverls of CO2on photosynthesis and cell components of the red algaPorphyraleucosticte[J]. Journal of Applied Phycology,1999,11:455-461.
[7] ISRAEL A,KATZ S,DUBINSKY Z,et al. Photosynthetic inorganic carbon utilization and growth ofPorphyralinearis(Rhorophyta)[J]. Journal of Applied Phycology,1999,11:447-453.
[8] 邹定辉,高坤山,阮祚禧. 高CO2浓度对石莼光合作用及营养盐吸收的影响[J]. 青岛海洋大学学报,2001,31(6):877-882.
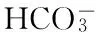
[10] ANDRIA J R,VERGARA J J,PEREZ-LLORENS J L. Biochemical responses and photosynthetic performance ofGracilariansp. (Rhodophyta) from Cadiz,Spain,cultured under different inorganic carbon and nitrogen levels[J]. Journal of Phycology,1999,34:497-504.
[11] 包楠欧,史定刚,关万春,等. CO2及光强对南麂列岛铜藻生长的影响[J]. 浙江农业学报,2014,26(3):649-655.
[12] 张玉荣,刘峰,单体锋,等. 利用叶绿素荧光技术揭示人工培育的铜藻幼苗对胁迫温度、光照和盐度的反应[J]. 南方水产科学,2009,5(2):1-9.
[13] 徐智广,邹定辉,高坤山,等. 不同温度、光照强度和硝氮浓度下龙须菜对无机磷吸收的影响[J]. 水产学报,2011,35(7):1023-1029.
[14] 徐智广,李美真,孙福新,等. 温度、光强和营养史对羊栖菜无机磷吸收的影响[J]. 南方水产科学,2013,9(3):8-13.
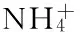
[16] LONG S P,WOODWARD F I. Plants and temperature[M]. Cambridge,United Kingdom:Company of Biologists,1988.
[17] 严国安,李益健,王志坚,等. 固定化栅藻对污水的净化及其生理特征的变化[J]. 中国环境科学,1995,15(1):10-13.
[18] 荆玉祥,匡延云,李德葆. 植物分子生物学:成就与前景[M]. 北京:科学出版社,1995.
[19] 邹定辉,夏建荣. 大型海藻的营养代谢及其与近岸海域富营养化的关系[J]. 生态学杂志,2011,30(3):589-595.
[20] PEDERSON M F,PALING E I,WALKER D I,et al. Nitrogen uptake and allocation in the seagrassAmphibolisAntarctica[J]. Aquatic Botany,1997,56:105-117.
(责任编辑:张瑞麟)
2016-08-09
国家级大学生创新创业训练计划(2014103343007);浙江省科技计划项目(2014F10005);温州医科大学大学生科研课题资助(wyx201501099)
杨安强(1994—),哈尼族,本科生,E-mail: yanganqiang0426@126.com。
关万春,副教授,博士,从事藻类学研究工作,E-mail: gwc@wmu.edu.cn。
10.16178/j.issn.0528-9017.20170236
Q949.2
A
0528-9017(2017)02-0303-05
文献著录格式:杨安强,史定刚,陈少波,等. 高CO2条件下温度和光照对铜藻吸收N、P的短期影响[J].浙江农业科学,2017,58(2):303-307.
