粳稻柱头外露率QTL定位
2017-02-20李威圣忠华朱子亮魏祥进石磊邬亚文唐绍清王建龙胡培松
李威圣忠华朱子亮魏祥进石磊邬亚文唐绍清王建龙,∗胡培松,,∗
(1湖南农业大学农学院,长沙410128;2中国水稻研究所,杭州310006;#共同第一作者;∗通讯联系人,E-mail:wjl9678@126.com)
粳稻柱头外露率QTL定位
李威1,#圣忠华2,#朱子亮1魏祥进2石磊1邬亚文2唐绍清2王建龙1,∗胡培松1,2,∗
(1湖南农业大学农学院,长沙410128;2中国水稻研究所,杭州310006;#共同第一作者;∗通讯联系人,E-mail:wjl9678@126.com)
【目的】柱头外露是影响杂交水稻制种的一个重要性状。本研究旨在初步鉴定控制柱头外露率的QTL。【方法】以高柱头外露率粳稻两系不育系大S×低柱头外露率热带粳稻品系D50的F2群体及海南陵水、浙江杭州两地重复种植的F2:3群体为材料,考查F2群体及陵水、杭州两地两次重复F2:3群体单边柱头外露率(PSES)、双边柱头外露率(PDES)和柱头总外露率(PES)。【结果】相关分析表明,单边柱头外露率、双边柱头外露率和柱头总外露率之间存在极显著的正相关。根据构建的大S×D50 F2群体遗传连锁图谱,QTL分析检测到水稻柱头外露率QTL 21个,分布于第1、2、3、4、6、7和12染色体上,贡献率变幅为0.1%~57.6%,除个别贡献率极低的位点外,绝大部分QTL加性效应来自于高值亲本大S。其中位于第4染色体和第12染色体的两个位点qPDES4,q PES12在F2和两次重复F2:3群体中均检测到,贡献率分别达17.86%和16.98%。位于第3染色体的qPES3位点在陵水、杭州两次F2:3群体中重复检测到,贡献率高达25.6%。【结论】检测到的QTL,有些与其他学者报道的处于同一位置,有些则是新的柱头外露率QTL。这些水稻柱头外露率QTL为今后柱头外露率基因精细定位、克隆、以及利用分子标记辅助选择(MAS)选育高柱头外露率粳型不育系提供理论依据。
柱头外露率;QTL定位;分子标记辅助选择(MAS);粳稻
水稻是我国最主要的粮食作物之一。水稻单产的高低事关我国粮食安全。近半个世纪以来,三系和两系杂交水稻体系相继研制成功,大大提高了我国水稻单产,保障了我国的粮食安全[1,2]。然而杂交水稻在发展初期也受诸多因素困扰,其中尤为突出的是制种产量低、异交结实差,从而导致制种成本居高不下,限制了杂交水稻尤其是杂交粳稻的大面积推广[3]。在水稻的驯化进程中,水稻的天然异交结实率由野生稻的100%降至栽培稻的2%~4%[4]。Ying等[5]考查了2065份非洲稻种资源的柱头外露率情况,发现柱头外露率从斑点野生稻(100%)、长药野生稻(85.7%)、非洲野生稻(54.9%)、巴蒂野生稻(34.8%)、杂草型野生稻(20.0%)、亚洲栽培稻(1.0%)依次降低。柱头外露率在水稻籼粳亚种间差异显著,尤其籼粳不育系之间,一般而言,籼稻品种的柱头外露率普遍高于粳稻品种[6]。
粳稻品种柱头外露率低,特别是低柱头外露率的粳稻不育系,异交结实差,制种产量低,严重制约着杂交粳稻的发展。目前对于控制杂交水稻异交特性,尤其是水稻柱头外露率的分子机理研究尚不够深入;国内外学者大多利用籼型高柱头外露率水稻与粳型低柱头外露率水稻品种构建临时性或永久性群体,筛选检测控制水稻柱头外露率的QTL[7]。邓应德等[8]利用柱头外露率差异显著的优ⅠB和糯5号构建F2群体,检测到3个分别位于第2、5、8染色体的柱头外露率QTL:q PES-2、q PES-5、q PES-8,贡献率分别为10.1%,11.1%,9.0%,另外,他们利用高柱头外露率亲本Ⅱ-32B和低柱头外露率亲本冈46B构建F2定位群体,采用单标记法成功筛选到分别位于第1、2、5、8染色体上的4个柱头外露率QTL,其中位于第5染色体的位点贡献率最高,达到16.65%,被认为是主效QTL[9]。王嫘等[10]利用高柱头外露率品种DS和低柱头外露率品种C9083构建的F2定位群体,共检测到16个分别位于第1、3、7、9染色体上的柱头外露率相关QTL,其中qPSES-1a的LOD达到12.18,贡献率为37.62%, q PDES-3a的LOD达到14.99,贡献率为31.18%, q PES-3b的LOD达到19.69,贡献率为41.91%;增效基因均来自于DS,是迄今为止鉴定的贡献率最高的QTL。冯玲玲等[11]以高柱头外露率的不育系50S作母本,低柱头外露率的连B为父本,构建F2定位群体,共检测到分别位于第3、9、12染色体上的3个QTL(q PES-3,q PES-9,q PES-12);增效基因均来自于不育系50S。岳高红等[12]利用旱稻保持系沪旱1B与Ⅱ-32B构建临时F2群体,检测到位于第3、7、9染色体的3个单边外露率位点,位于第3、9染色体的2个双边外露率位点和位于第3、4、7、9染色体上的5个总柱头外露率位点,其中位于第3染色体RM6932-RM6959区间的q PES-3同时控制单边外露、双边外露和总柱头外露。Miyata等[13]利用IR24和Koshihikari构建的F2群体定位到第3染色体上的柱头外露率位点q PES-3,该位点成功地在水稻新育品种中增加了20%的外露率。同时,研究表明,GS3也位于该区间内,GS3在增长粒型的同时增加了柱头的细胞数目,导致柱头伸长,进而提高柱头外露率。
对于临时性F2群体检测到的QTL存在遗传不稳定性,同时受环境影响,因而其检测准确性不高。永久性遗传群体,由于遗传稳定,可多年多点重复实验,故检测到的QTL相对可靠。尹成等[14]使用岳早籼9号和Ⅱ-32B构建的重组自交系群体,共检测到16个分别位于第1、3、5、6、7、9染色体上的QTL,其中只有第1、9染色体上的位点在两年的重复数据中被稳定检测到。沈圣泉等[15]使用协青早B×密阳46构建的RIL群体和相应的分子遗传图谱,设置海南和浙江两点试验,检测到一个LOD为28.16,贡献率为14.14%的柱头外露率QTL。此外,Hu等[16]利用保持系珍汕97B和IRAT109构建重组自交系,在正常和干旱胁迫条件下分别检测到3个和4个柱头外露率QTL,其中仅有q PES-1在两种环境下都能检测到。李勇等[17]用川香29B× Lemont的F5群体,检测到3个柱头外露率QTL,分别位于3条染色体上,命名为q PES-1、q PES-2、qPES-10。李文宏等[18]利用籼稻品种窄叶青8号和粳稻品种京系17构建的DH群体,成功地在第2、3染色体上分别定位到2个QTL:qPES-2和q PES-3,同时认为总柱头外露率和单边柱头外露率的QTL位点一致。由于水稻柱头外露率特性属数量性状,遗传不稳定,受环境因素影响较大,故不同学者采用不同的定位群体检测到的水稻柱头外露率QTL位点差异较大。
本研究以高柱头外露率的粳稻两系不育系大S为材料,通过与低柱头外露的热带粳稻品种D50杂交构建F2及F2:3遗传群体,对不同世代群体柱头外露性状重复调查分析,旨在初步鉴定稳定遗传的控制大S高柱头外露率的QTL。研究结果为进一步探究控制水稻柱头外露率基因的克隆,基因功能分析等分子机理提供理论指导。同时对育种家利用分子标记辅助选择(MAS)选育高柱头外露的粳型不育系,突破杂交粳稻制种瓶颈,促进杂交粳稻发展提供技术支持。
1 材料与方法
1.1 供试材料
大S为高柱头外露率的粳稻两系不育系;D50为低柱头外露率的热带粳稻品种;大S×D50 F2群体包含177个正常可育单株,F2:3群体包含F2群体衍生的177个株系。
1.2 群体构建及农艺性状考查
2013年夏以大S为母本、D50为父本杂交,收F1;2013年冬季种植F1于海南陵水,去杂后收F1种子;2014年夏季种植F2群体于杭州。在抽穗期,从中随机选择177个正常可育单株编号,待其开花完全后取各单株一个主穗及分蘖穗分别考查单边柱头外露率(PSES)、双边柱头外露率(PDES)及柱头总外露率(PES)。F2群体成熟时按单株编号收种即为F2:3株系。各单株种子分成两份。2014年冬,将亲本及177个F2:3株系种植于海南陵水,待开花完全后每株系随机取6株主穗考查单边柱头外露率、双边柱头外露率及柱头总外露率;2015年夏季,将亲本与另一份177个F2:3株系种植于杭州,待开花后每株系随机选取6株主穗,考查单边柱头外露率,双边柱头外露率及柱头总外露率。柱头外露率计算方法如下:单边柱头外露率(%)=单边柱头外露颖花数/总颖花数×100%;双边柱头外露率(%)=双边柱头外露颖花数/总颖花数×100%;柱头总外露率(%)=单边柱头外露率+双边柱头外露率。
1.3 SSR分子标记连锁图谱构建及QTL定位分析
SDS法提取亲本及177个F2单株DNA[19]。利用本实验室的556对以及擎科生物公司合成的83对均匀分布于水稻12条染色体上的SSR标记筛选大S和D50之间的多态性。
利用筛选出的均匀分布在水稻12条染色体上且在大S和D50间具有多态性的131对SSR标记对177个F2单株进行基因型分析,采用Map Maker 3.0软件构建连锁图谱。遗传连锁图谱结合各项表型数据,采用Win QTL cartographer 2.5软件,取LOD≥2.5进行QTL分析[20]。采用McCouch命名系统对QTL命名[21]。
2 结果与分析
2.1 大S×D50 F2、F2:3群体及亲本柱头外露率表型分析
大S为高柱头外露率粳稻不育系,D50为低柱头外露率热带粳稻品种,其表型见表1和图1。大S ×D50 F2和F2:3群体,两代三次实验的柱头外露率表型统计参数和表型分布见图表1和图2。F2及F2:3群体柱头外露率表型分离明显,呈连续正态分布(图2),显示出数量性状的特征,并有超亲分离现象;其中单边柱头外露率在两年三次重复中的偏值小于-1;双边柱头外露率与柱头总外露率偏值基本在±1以内(表1),表明基本呈正态分布,符合QTL作图要求。单边柱头外露率、双边柱头外露率及柱头总外露率表型频率分布在两代三次实验中基本一致,显示出较好的重复性。

表1 大S×D50 F2与F2:3群体及亲本的柱头外露率表型Table 1.The stigma exsertion rate of F2and F2:3 populations of Da S×D50 combination.

图1 大S和D50柱头外露表型Fig.1.Phenotype of stigma exsertion of Da S and D50.
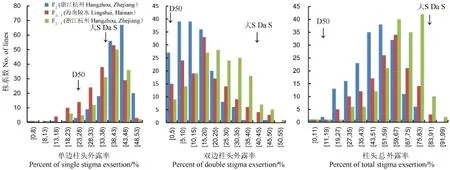
图2 大S×D50 F2和F2:3群体柱头外露率频率分布Fig.2.The frequency distributions of stigma exsertion rate of F2and F2:3populations of Da S×D50 combination.
2.2 大S×D50 F2、F2:3群体柱头外露率性状间相关性显著
对大S×D50的F2和F2:3群体柱头外露率性状间相关性分析(表2)表明:单边柱头外露率、双边柱头外露率及柱头总外露率三个性状之间在两代三次实验中均表现出极显著的正相关,表明水稻柱头外露率性状之间关系密切,互相影响。
2.3 大S×D50 F2、F2:3群体柱头外露率的QTL分析
利用131对在大S与D50之间表现多态性的SSR标记,采用Map Maker 3.0构建了遗传连锁图谱,图谱总长为1865.6 c M,平均距离为14.6 c M。采用Win QTL cartographer 2.5进行QTL区间作图分析(表3和图3),两代3次重复共检测到21个柱头外露率QTL,分别位于第1、2、3、4、6、7、12染色体上,其中单边柱头外露率QTL 6个,双边柱头外露率QTL 7个,柱头总外露率QTL 8个;至少在两次重复中检测到的QTL有5个,分别为q PSES 3.1,q PSES12,q PDES4,q PES4和q PES12,其中qPSES12、qPDES4和qPES12在F2及2个F2:3群体中均被检测到,且其加性效应均来自于高值亲本大S,表明这些QTL受环境影响相对较小,不同条件下都能稳定遗传表达。
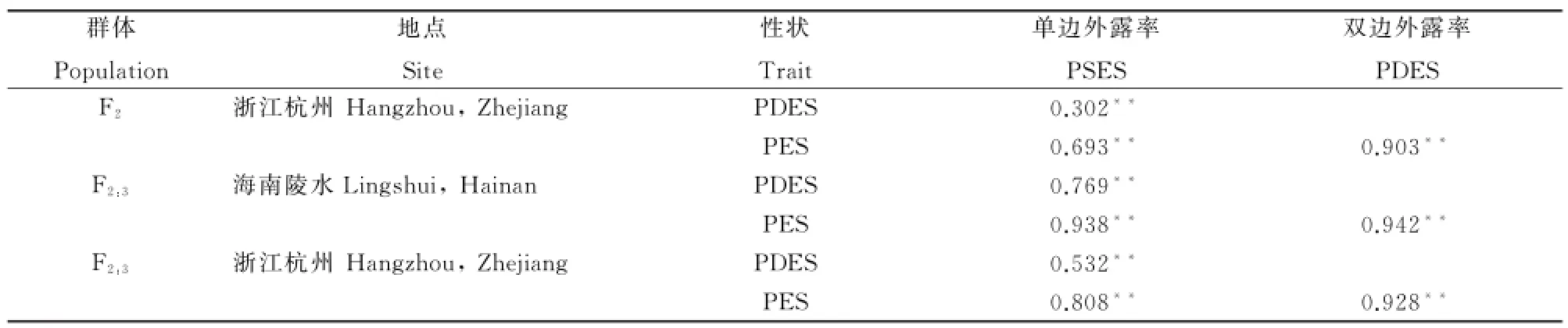
表2 大S×D50 F2、F2:3群体柱头外露率性状间的相关分析Table 2.Correlation analysis among stigma exsertion rate in F2and F2:3 populations of Da S×D50 combination.
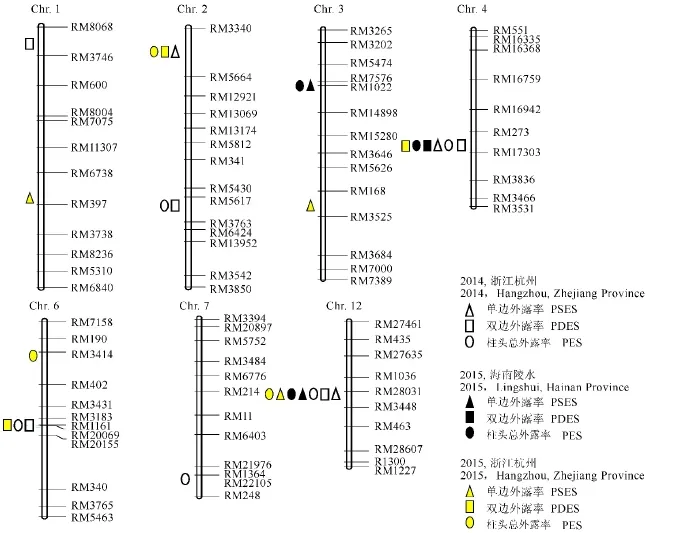
图3 大S×D50 F2与F2:3群体检测到的柱头外露率QTLFig.3.The stigma exsertion rate QTLs detected in F2and F2:3 populations of Da S×D50 combination.

表3 大S×D50 F2与F2:3群体检测到的柱头外露率QTL分布Table 3.The QTL distributions of stigma exsertion rate detected in F2and F2:3 populations of Da S×D50.
在大S×D50 F2群体,以及陵水、杭州的F2:3群体中检测到的单边柱头外露率QTL共有6个,分别为q PSES1、q PSES2、q PSES3.1、q PSES3.2、q PES4、q PSES12,贡献率为0.12%~57.67%(表3和图3)。其中q PSES12在F2群体,以及陵水、杭州F2:3群体中均被检测到,该位点在F2群体的加性效应来自于D50,但贡献率极低;在陵水和杭州F2:3群体中被检测的加性效应来自于大S,贡献率达16.98%。q PSES3.1在陵水、杭州F2:3群体中均检测到,贡献率达25.6%,来自大S的等位基因可解释表型变异的5.58%。在这些被检测到的单边柱头外露率QTL中,绝大部分位点的加性效应均来自大S。
3 讨论
本研究利用高柱头外露率的粳稻两系不育系大S为研究材料所定位到的水稻柱头外露率QTL,部分与其他学者利用高柱头外露率的籼稻品种为研究对象所得到的研究结果相似,同时也检测到新的稳定遗传的柱头外露率QTL(表3)。本研究共检测到21个水稻柱头外露率QTL,位于10个标记区间。其中位于同一区间的控制单边外露率的q PES3.1和总外露率的q PES3,在尹成等[14]、李海滨等[22]和王嫘等[10]的研究中均见报道,且贡献率较高;在王嫘等[10]的研究中贡献率更是高达31.18%;本研究在两次重复的F2:3群体中检测到,其贡献率分别达到21.87%和25.60%。本研究检测到的单边柱头外露率位点q PSES1与李勇等[17]、李海滨等[22]和Wen等[23]的报道中类似。本研究控制双边外露率的qPDES1与李海滨等[22]所报道的q PES-1,Li等[24]所报道的qPDES-1位于同一个区间;另外,本研究同时控制单边、双边和柱头总外露率的QTL区间有3个,分别是第2染色体的RM3340-RM5664、第4染色体的RM16942-RM17303和第12染色体的RM1036-RM3448,其中,除第4染色体的定位区间外,第2染色体和第12染色体所定位区间分别与邓应德[9]等检测到的与RM3188紧密连锁的QTL,Li等[24]报道的qSSE-12位置相近。此外,本研究中q PDES2.1、q PES2.1与李勇等[17]报道的q PES-2,陈爱国等[25]检测到的q PGCES-2两个位点同属RM5430-RM6424区间。与标记RM22105紧密连锁的q PES7和尹成等[14]所报道的q PDES-7-2位置非常接近。不同学者检测水稻柱头外露率QTL位点存在相似性,表明确实存在控制水稻柱头外露的QTL及基因,值得进一步的基因挖掘与应用。
本研究检测到的柱头外露率QTL,绝大多数的表型变异来自大S,表明大S确实携带控制水稻高柱头外露率的QTL。同时本研究检测到的q PSES12在杭州F2群体,以及陵水、杭州F2:3群体中均被检测到,而该位点在F2群体的加性效应来自D50,但贡献率极低;在陵水和杭州种植的F2:3群体中被检测的加性效应来自大S,其可能原因是水稻柱头外露特性受遗传特性、环境影响较大,不同的环境条件、不同世代,单株及株系柱头外露率表型值波动较大,影响检测结果[26]。q PSES3.1在陵水、杭州F2:3群体中均检测到,而在F2群体中未被检测到,表明某些效应值较小的QTL,随着世代的增加其效应值有可能增大。另外,q PSES4、q PDES4、q PES4位点在F2群体,以及陵水、杭州两次重复F2:3群体中稳定检测到,贡献率较大,且位于同一定位区间,可能是一个新的影响柱头外露率的QTL,目前已构建相应的次级分离群体,以期将该控制水稻外露率的QTL精细定位甚至克隆。
在F2和F2:3群体,两代三次实验中也检测到新的控制柱头外露率的QTL,如:q PSES4、q PDES4、q PES4,值得进一步深入研究。此外,我们在杭州F2:3群体中还检测到一个新的贡献率高达57%的有待进一步确认的柱头单边外露率QTL qPSES3.2。针对已报道的且贡献率较大的QTL以及最新检测到的稳定遗传的QTL,一方面可构建次级分离群体进一步深入研究,将控制柱头外露率的基因进行克隆、功能分析,阐明控制柱头外露高低的分子机理。另一方面,精细定位柱头外露率QTL,开发相应的与柱头外露率紧密连锁的功能分子标记[27],利用分子标记辅助选择选育高柱头外露率的粳稻类型的保持系、不育系等,进而促进杂交粳稻发展。
[1] 岳高红,潘彬荣,许立奎,楼钰,梅捍卫,罗利军.水稻柱头外露率的研究进展.种子,2009,28(1):47-50.Yue G H,Pan B R,Xu L K,Lou Y,Mei H W,Luo L J.Progress of study on rice stigmaexsertion.Seed,2009,28(1): 47-50.(in Chinese)
[2] 袁隆平.杂交水稻学.北京:中国农业出版社,2002:37-40.Yuan L P.Hybrid Rice.Beijing:China Agriculture Press, 2002:37-40.
[3] Marathi B,Jena K K.Floral traits to enhance outcrossing for higher hybrid seed production in rice:Present status and future prospects.Euphytica,2015,201(1):1-14.
[4] 杜雪树,戚华雄.水稻柱头外露率研究进展.湖北农业科学, 2015,54(16):3841-3843,3850.Du X S,Qi H X.Progress on the stigmaexsertion in rice.Hubei Agric Sci,2015,54(16):3841-3843,3850.(in Chinese with English abstract)
[5] Ying C S,Zhang S Q.Studies on the character of stigma exsertion among some of Oryza species.Chin J Rice Sci,1989, 3(2):62-66.
[6] 许克农,李泽炳,李成荃.光(温)敏核不育水稻的育性和开花习性研究.安徽农业科学,1992,20(4):293-301.Xu K N,Li Z B,Li C Q.Studies on fertility and flowering habits of photoperiod(thermo-)sensitive genic male-sterile rice.J Anhui Agric Sci,1992,20(4):293-301.(in Chinese with English abstract)
[7] 邓华凤,何强,舒服,张武汉,杨飞,荆彦辉,东丽,谢辉.中国杂交粳稻研究现状与对策.杂交水稻,2006,21(1):1-6.Deng H F,He Q,Shu F,Zhang W H,Yang F,Jing Y H, Dong L,Xie H.Status and technical strategy on development on japonica hybrid rice in China.Hybrid Rice,2006,21(1):1-6.(in Chinese with English abstract)
[8] 邓应德,应杰政,石媛媛,肖层林,张海清.控制水稻柱头外露率的数量性状基因座初步分析.湖南农业大学学报:自然科学版,2010,36(4):373-376.Deng Y D,Ying J Z,Shi Y Y,Xiao C L,Zhang H Q.Mapping of QTL for percentage of exserted stigma in rice.J Hunan Agric Univ:Nat Sci,2010,36(4):373-376.
[9] 邓应德,肖层林,邓化冰,张海清,邓晓建,刘玉龙.用近等基因池法定位与水稻柱头外露率相关的QTL.农业现代化研究, 2011,32(2):230-233.Deng Y D,Xiao C L,Deng H B,Zhang H Q,Deng X J,Liu Y L.Detection of QTL related to stigmaexsertion rate(SER) in rice(Oryza sativa L.)by bulked segregant analysis.Res Agric Mod,2011,32(2):230-233.(in Chinese with English abstract)
[10]王嫘.水稻光温敏不育系DS的遗传分析与基因定位.天津:天津科技大学,2012.Wang L.Genetic analysis and gene mapping of the photoperiodthermo sensitive genic male sterile rice line DS.Tianjin:Tianjin University of Science&Technology,2012.(in Chinese with English abstract)
[11]冯玲玲,荆彦辉,黄成,徐正进,陈温福.水稻柱头外露率的QTL分析.北方水稻,2010,40(3):20-22.Feng L L,Jing Y H,Huang C,Xu Z J,Chen W F.QTL analysis for percentage of exserted stigma in rice(Oryza sativa L.).North Rice,2010,40(3):20-22.(in Chinese with English abstract)
[12]岳高红,梅捍卫,潘彬荣,楼钰,李明寿,罗利军.杂交旱稻保持系沪旱1B柱头外露率的QTL定位.浙江农业学报,2009, 1(3):241-245.Yue G H,Mei H W,Pan B R,Lou Y,Li M S,Luo L J.Mapping of QTL affecting stigma exsertion rate of Huhan 1B as a CMS maintainer of upland hybrid rice.Acta Agric Zhejiang, 2009,1(3):241-245.(in Chinese with English abstract)
[13]Miyata M,Yamamoto T,Komori T,Nitta N.Marker-assisted selection and evaluation of the QTL for stigmaexsertion under japonica rice genetic background.Theor Appl Genet,2007,114:539-548.
[14]尹成,李平波,高冠军,张庆路,罗利军,何予卿.水稻柱头外露率QTL定位.分子植物育种,2014,12(1):43-49.Yin C,Li P B,Gao G J,Zhang Q L,Luo L J,He Y Q.QTL analysis of percentage ofexserted stigma in rice.Mol Plant Breeding,2014,12(1):43-49.(in Chinese with English abstract)
[15]沈圣泉,庄杰云,包劲松,舒庆尧,夏英武.水稻柱头外露率QTL定位及其互作分析.生物数学学报,2006,21(4):610-614.Shen S Q,Zhuang J Y,Bao J S,Shu Q Y,Xia Y W.Analysis on QTL with main,epistasis and G×E interaction effects of stigma extruding trait in rice.J Biomath,2006,21(4):610-614.(in Chinese with English abstract)
[16]Hu SP,Zhou Y,Zhang L,Zhu X D,Wang Z G,Li L,Luo L J,Zhou Q M.QTL analysis of floral traits of rice(Oryza sativa L.)under well-watered and drought stress conditions.Gene Genom,2009,31(2):173-181.
[17]李勇.水稻柱头几个数量性状的研究与QTL遗传定位分析.成都:四川师范大学,2009.Li Y.Several quantitative traits in rice research and QTL genetic analysis.Chengdu:Sichuan Normal University,2009.
[18]李文宏,董国军,胡新民,滕胜,郭龙彪,曾大力,钱前.水稻柱头外露率的QTL分析.遗传学报,2003,30(7):637-640.Li W H,Dong G J,Hu X M,Teng S,Guo L B,Zeng D L, Qian Q.QTL analysis for percentage ofexserted stigma in rice (Oryza sativa L.).Acta Genet Sin,2003,30(7):637-640.
[19]Orjuela J,Garavito A,Bouniol M,Arbelaez J D,Moreno L, Kinball J,Wilson G,Rami J,Tohme J,Mclouch SR.A universal core genetic map for rice.Theor Appl Genet,2010,120: 563-572.
[20]Wang S,Basten C J,Zeng Z B.Windows QTL cartographer 2.5 department of statistics.Raleigh,USA:North Carolina State University,2006.
[21]McCouch S R.Gene nomenclature system for rice,Rice, 2008,1(1):72-84.
[22]李海滨,高方远,曾礼华,李勤修,陆贤军,李治军,任鄄胜,苏相文,任光俊.长药野生稻导入系柱头性状的QTL分析.分子植物育种,2010,8(6):1082-1089.Li H B,Gao F Y,Zeng L H,Li Q X,Lu X J,Li Z J,Ren J S, Su X W,Ren G J.QTL analysis of rice stigma morphology using an introgression line from Oryza longistaminata L.Mol Plant Breeding,2010,8(6):1082-1089.(in Chinese with English abstract)
[23]Yan W G,Li Y,Agrama H A,Luo D G,Guo F Y,Lu X J, Ren G J.Association mapping of stigma and spikelet characteristics in rice(Oryza sativa L.).Mol Breed,2009,24(3): 277-292.
[24]LiP B,Feng F C,Zhang Q L.Genetic mapping and validation of quantitative trait loci for stigma exertion rate in rice.Mol Breeding,2014,34:2131-2138.
[25]陈爱国,华泽田,王嫘,李志彬,苏玉安,肖艳云,王彦荣.水稻柱头外露性状的遗传分析及QTL定位.沈阳农业大学学报, 2011,02:142-146.Chen A G,Hua Z T,Wang L,Li Z B,Su Y A,Xiao Y Y, Wang Y R.Inheritance analysis and detection of QTL for exserted stigma rate in rice.J Shenyang Agric Univ,2011,42 (2):142-146.(in Chinese with English abstract)
[26]史开兵,邓华凤.中国杂交粳稻科技创新研讨会在三亚召开.杂交水稻,2004,19(3):76.Shi K B,Deng H F.The National symposium of Chinese japonica hybrid rice research held at Sanya,Hainan.Hybrid Rice,2004,19(3):76.
[27]邱福林,庄杰云,华泽田,王彦荣,程式华.北方杂交粳稻骨干亲本遗传差异的SSR标记检测.中国水稻科学,2005,19(2): 101-104.Qiu F L,Zhuang J Y,Hua Z T,Wang Y R,Cheng S H.Inspect of genetic differentiation of main parents of japonica hybrid rice in the northern China by simple sequence repeats (SSR).Chin J Rice Sci,2005,19(2):101-104.(in Chinese with English abstract)
QTL Mapping of japonica Rice Stigma Exsertion Rate
LI Wei1,#,SHENG Zhonghua2,#,ZHU Ziliang1,WEI Xiangjin2,SHI Lei1,WU Yawen2,TANG Shaoqing2, WANG Jianlong1,∗,HU Peisong1,2,∗
(1College of Agronomy,Hunan Agricultural University,Changsha 410128,China;2China National Rice Research Institute, Hangzhou 310006,China;#The authors contributed equally to this work;∗Corresponding author,E-mail:wjl9678@126.com)
【Objective】Stigma exsertion is one of the important traits which contribute to the efficient improrement of commercial seed production in hybrid rice.The objective of the study was to understand the genetic factors involved in the sigma exsertion.【Method】The F2and F2:3populations of Da S×D50 planted in Lingshui,Hainan Province and Hangzhou,Zhejiang Province,respectively were used and the single stigma exsertion rate(PSES),double stigma exsertion rate(PDES)and total stigma exsertion rate(PES)were measured.【Result】Correlation analysis indicated that there were significant relationships among the PSES,PDES and PES.Further,the genetic linkage map of Da S× D50 F2population was constructed,then,21 stigma exsertion rate QTL were located on chromosome 1,2,3,4,6,7 and 12,respectively,with the contribution rate ranging from 0.1%to 57.6%.The additive effect of most QTLs were contributed by high value parent Da Sexcept for some QTL that with extremely low contribution rate.There were two QTLs on Chromosome 4 and Chromosome 12,q PDES4 and q PES12,both detected in F2and F2:3populations with contribution rate as high as 17.86%and 16.98%,respectively.The q PES3 located on Chromosome 3 with high contribution rate were detected in F2:3population for two times.【Conclusion】Some stigma exsertion rate QTL shared the same location with those reported by other researchers,but there were also some new QTLs which provide theoretical support for further fine-mapping,gene cloning,and breeding new japonica male sterile lines with high stigma exsertion rate by using molecular marker-assistant selection.
stigma exsertion rate;QTL mapping;marker-assistant selection(MAS);japonica rice
Q343.1+5;S511.0351
A
1001-7216(2017)01-0023-08
2016-03-14;修改稿收到日期:2016-07-19。
国家自然科学基金资助项目(31501285);浙江省自然科学基金资助项目(LY14C130009);浙江省公益技术应用研究计划资助项目(2015C32045)。
