DRD2基因TaqIA多态性与同伴侵害对青少年早期抑郁的交互作用*
2017-02-01
(山东师范大学心理学院, 济南 250014)
1 问题提出
抑郁(depression)是一种常见的情绪障碍和心理健康问题。世界卫生组织预测, 到2030年抑郁将成为引发死亡和导致疾病负担的首要因素(Lépine& Briley, 2011)。青少年早期是抑郁发生的关键期。抑郁的发病率和严重性程度在青少年早期开始迅速增加(Avenevoli, Swendsen, He, Burstein, & Merikangas,2015; Hankin, 2015), 且追踪研究显示青少年抑郁在14 ~ 15岁左右达到顶峰(Natsuaki, Biehl, & Ge,2009; Twenge & Nolen-Hoeksema, 2002)。该时期的抑郁不仅会损害青少年的心理社会适应(Verboom,Sijtsema, Verhulst, Penninx, & Ormel, 2014), 其消极影响还会持续至成年阶段, 致使个体罹患重性抑郁的风险显著增加(McLeod, Horwood, & Fergusson,2016)。因此, 探究青少年早期抑郁的影响因素及其作用机制是该研究领域的重要任务。
抑郁具有重要的遗传基础并受环境因素的影响。近十几年来, 研究者对导致青少年抑郁的分子遗传作用机制进行了大量的研究。继 Caspi等人(2003)里程碑式的研究之后, 越来越多的研究发现基因×环境交互(G×E)影响抑郁的发生发展(Banny,Cicchetti, Rogosch, Oshri, & Crick, 2013; Chen, Li,& McGue, 2012, 2013; W.X.Zhang et al., 2015; L.L.Zhang et al., 2016)。如Chen等(2013)采用双生子样本检验了 BDNF基因(brain-derived neurotrophic factor, 脑源性神经营养因子)Val66Met多态性与压力性生活事件对青少年抑郁的交互效应, 结果发现,相比MetMet纯合子携带者, 携带ValVal/ValMet基因型的青少年更可能受到压力性生活事件的影响而具有较高的抑郁水平。
根据抑郁的单胺缺陷假说(the monoamine deficiency hypothesis), 多巴胺(dopamine, DA)受体功能的缺陷及其合成或储存障碍是导致抑郁的主要原因之一(Belmaker & Agam, 2008)。DRD2基因(dopamine receptor D2, D2型多巴胺受体)TaqIA多态性是众多多巴胺候选基因中的一种功能性多态性位点, 它影响大脑纹状体区(情绪和奖赏敏感性相关脑区)的功能, 因而备受研究者关注。既有研究也表明, DRD2基因TaqIA多态性与抑郁密切相关(Dunn et al., 2011; Zhang et al., 2015)。该多态性位于11号染色体的q2.2~2.3区, DRD2基因下游10kbp处, 编码D2型多巴胺受体。该位点处谷氨酸(T或A1)到赖氨酸(C或 A2)的置换, 导致大脑纹状体区域D2受体密度降低30% ~ 40% (Ritchie & Noble,2003), 并且中枢神经系统的多巴胺活性较低(Mihara et al., 2003)。此外, 研究显示相比A1等位基因携带者, A2A2基因型携带者在面对消极反馈时表现出喙部扣带回区域(rostral cingulate zone)激活水平的相对增加(Klein et al., 2007), 而喙部扣带回区域的异常激活与重性抑郁相关(Liberg et al.,2013)。由此, DRD2基因可能与个体对环境的敏感性差异有关, 即DRD2基因与环境因素可能存在交互效应。
越来越多的研究显示, DRD2基因与环境因素交互作用于个体的抑郁症状。譬如, Elovainio等人(2007)对1611名成年人的研究显示, DRD2基因与压力性生活事件交互影响个体抑郁, 相比 A1等位基因携带者, 携带DRD2 A2A2基因型的个体在经历了较多的压力性生活事件后报告了更高的抑郁水平。Hayden等人(2010)的研究也发现携带DRD2基因A2A2纯合子的个体在经历了较多的消极教养后报告了更高的焦虑、抑郁。此外, van Roekel,Goossens, Scholte, Engels和Verhagen (2011)对孤独感的研究显示, 在携带 A2A2基因型的青少年中,低水平的父母支持可以正向预测青少年孤独感水平, 而在 A1等位基因携带者中, 父母支持与青少年孤独感水平无关。这些研究较为一致的发现,DRD2基因与环境经历(如消极教养、压力性生活事件、低水平的父母支持)交互影响抑郁及其相关特质。但是, 迄今有关DRD2基因与环境对抑郁的交互效应研究所选取的环境指标均是集中于家庭环境和负性生活事件, 如亲子关系、父母教养行为、童年期虐待等(参见综述Dunn et al., 2011)。然而,为了更全面的揭示基因×环境交互作用过程, 有必要关注其他环境背景。
同伴环境是青少年发展的重要微观环境变量之一。尤其在进入青春期后, 青少年的人际关系开始由父母中心向同伴中心过渡(Steinberg, 2014)。同时, 伴随父母监控的减少、同伴群体规模和多样性的增加, 使得青少年暴露于消极同伴环境的风险增加(Sugimura & Rudolph, 2012), 由此增加了青少年罹患抑郁等情绪障碍的风险。其中, 同伴侵害(peer victimization)是青少年抑郁症状的重要预测源(见元分析Hawker & Boulton, 2000)。同伴侵害是指个体遭受同伴的身体、言语、财务以及人际关系方面攻击的经历(Mynard & Joseph, 2000)。身体侵害(physical victimization)与关系侵害(relational victimization)是同伴侵害的两种典型类型, 前者是指儿童遭受同伴身体攻击或伤害的经历, 具有直接性和公开性的特点; 后者是指儿童在人际关系与社会互动方面受到同伴的蓄意伤害和攻击的经历(如遭受谣言中伤,被排斥在同伴活动之外), 具有间接性和隐蔽性的特点(Crick & Bigbee, 1998)。同伴侵害能够通过降低青少年在获得社会目标方面的自我效能感、引发青少年消极的自我认知, 进而导致焦虑、抑郁(Cole et al., 2014; Troop-Gordon & Ladd, 2005)。既有研究一致表明, 不同类型的同伴侵害(身体侵害和关系侵害)均能增加青少年罹患抑郁的风险(Crick & Bigbee,1998; Leadbeater, Thompson, & Sukhawathanakul, 2014;Schwartz, Lansford, Dodge, Pettit, & Bates, 2015)。值得指出的是, 迄今为止, 多数同伴侵害与抑郁的关联研究主要依赖于横断研究设计(参见元分析Hawker & Boulton, 2000)或者短期追踪设计(如采用6个月或1年的追踪设计, 参见元分析Reijntjes,Kamphuis, Prinzie, & Telch, 2010)。然而, 同伴侵害不仅影响当前抑郁, 而且对抑郁具有长时性影响(Leadbeater et al., 2014)。因此, 本研究采用间隔两年的追踪研究设计, 考察同伴侵害与DRD2基因对抑郁的长期效应, 旨在更清楚的解释抑郁的基因×环境交互作用机制。
同伴侵害与抑郁的关联依赖于个体的遗传素质。如Sugden等人(2010)的研究发现5-HTTLPR基因(serotonin-transporter-linked polymorphic region, 5-羟色胺转运体关联的多态性位点, 能够影响5-羟色胺神经递质系统活性)能够调节同伴侵害与青少年情绪问题间的关联, 携带S等位基因的个体更容易受到同伴侵害的影响表现出更多的情绪问题。此外,Benjet, Thompson 和 Gotlib (2010)和 Iyer, Dougall和 Jensen-Campbell (2013)等人对青少年抑郁的研究也发现了类似的结果。值得指出的是, 上述研究考察的均是 5-HTTLPR基因(5-羟色胺系统基因),但是, 目前尚未有研究考察多巴胺系统基因与同伴环境对抑郁的交互效应。既有研究提示, 多巴胺系统基因也可能调节消极同伴环境对青少年行为的影响。Goodnight, Bates, Newman, Dodge和Pettit(2006)的研究显示具有较高奖赏敏感性的儿童青少年更容易受到行为不良同伴的消极影响, 而个体过度的奖赏敏感性与多巴胺系统候选基因密切相关(Blum et al., 2011)。我们推测DRD2基因亦有可能与同伴侵害交互作用于青少年的抑郁。因此, 本研究采用同伴身体侵害和关系侵害作为环境指标, 考察其与DRD2基因TaqIA多态性对青少年抑郁的潜在交互作用。
对于男女青少年, 遗传因素和环境因素对抑郁交互作用可能存在不同模式。一些研究显示基因×环境仅能预测男性或者女性抑郁水平, 如有研究显示MAOA基因(monoamine oxidase A, 单胺氧化酶A, 参与代谢多巴胺、5-羟色胺等单胺类神经递质)rs6323多态性与母亲支持性教养行为交互作用于女青少年的抑郁, 母亲支持性教养显著负向预测GG基因型女青少年的抑郁, 但对TT基因型女青少年抑郁的预测作用并不显著(曹丛等, 2016)。另一些研究发现基因×环境对两性抑郁均存在预测作用,但作用模式或敏感基因型不同, 如 Brummett等(2008)发现携带5-HTTLPR S等位基因的女性在面临压力性生活事件(如亲属重病、低社会经济地位)时抑郁水平更高, 而携带5-HTTLPR L等位基因的男性在面临压力性生活事件时抑郁水平较高。对于DRD2基因, 目前仅有极少数几项研究对这一问题进行了考察, 且研究结论存在分歧。一些研究发现DRD2基因与环境的交互效应仅与女生抑郁存在显著关联。来自美国国家青少年健康追踪研究的报告显示, DRD2基因TaqIA多态性仅能够调节暴力侵害对非裔美国女性抑郁的影响, 携带 A1等位基因的非裔女性比携带A2A2基因型的非裔女性更容易受到暴力侵害的消极影响而产生抑郁, 在男性中,这一基因×环境交互作用不存在(Vaske, Makarios,Boisvert, Beaver, & Wright, 2009)。另一些研究则发现 DRD2基因与环境的交互效应与男孩抑郁的关联更加密切, 且与 Vaske等(2009)研究发现的抑郁易感基因型截然相反。如Berman等人(2003)对146名欧裔青少年的研究发现, 在父母酗酒的家庭风险环境中, 携带A2A2基因型的男青少年抑郁水平高于 A1等位基因携带者, 而在女青少年中, 两种基因型携带者的抑郁水平不存在显著差异。此外, 仅选取男性被试考察抑郁相关人格特质的间接研究证据显示:父母酗酒和外向性的关系受到DRD2基因的调节, 在酗酒家庭中, 携带 A2A2基因型的男孩外向性水平具有低于A1等位基因携带者的倾向(Ozkaragoz & Noble, 2000), 而低外向性与高抑郁水平有关(Watson, Stasik, Ellickson-Larew, & Stanton,2015)。上述研究在不同种族的被试中发现了截然不同的结果, 但是这些研究结果无法进行直接比较,也不能推广到中国被试样本中。此外, 仅选取男青少年为被试的研究(Ozkaragoz & Noble, 2000)也无法回答 DRD2基因与环境的交互作用是否存在性别差异以及具体的性别差异模式如何的问题。基于此, 有必要进一步考察DRD2基因与同伴环境对抑郁的作用机制是否存在性别差异以及具体的差异模式。
综上, 本研究采用DRD2基因TaqIA多态性为基因指标, 身体侵害与关系侵害为环境指标, 综合运用回归分析和方差分析考察基因与环境因素对青少年早期抑郁的影响, 并主要考察以下研究问题:(1)TaqIA多态性是否与同伴侵害交互预测青少年早期抑郁; (2)TaqIA多态性与同伴侵害对抑郁作用机制的性别差异。
2 研究方法
2.1 被试
本研究数据来自国内一项大型追踪项目。该追踪项目以班级为单位随机选取山东省济南市 14所小学(6所省级规范化小学, 8所普通小学)39个班级的 2133名小学三年级儿童作为被试(首次测评为2006年, 被试平均年龄9岁), 并且每年春季学期对这些儿童进行一次追踪测评。因为本研究关注青少年抑郁发展高峰期(14~15岁), 且旨在考察同伴侵害的长期效应, 因此参照既有文献资料(Leadbeater et al., 2014; Perren, Ettekal, & Ladd, 2013)采用间隔两年的追踪设计, 选取了该项追踪项目中的青少年早期的两个时间点(2009年和2011年的数据)。2009年测量青少年抑郁症状及同伴侵害(包括身体侵害和关系侵害)和家庭社会经济地位, 2011年收集青少年的基因数据并再次测评抑郁, 2011年时, 这些被试分布在8所初中的92个班级中。鉴于工作量和研究经费问题, 2011年仅随机选取1063名被试测评其 DRD2基因型, 男生 528人(49.7%), 女生535人(50.3%), 2011年被试的平均年龄为 14.32 ±0.47岁, 具有基因数据的样本(1063人)和最初样本(2133人)在基本人口学变量(性别、父母职业及受教育水平) (χ2s < 9.88,ps > 0.05)、抑郁及同伴侵害得分上(ts < 1.42,ps > 0.05)均不存在显著差异。母亲受教育水平在本科及本科以上者占 34.6%, 本科以下且高中以上者占53.7%, 高中以下者占11.7%。父亲受教育水平在本科及本科以上者占 45.6%, 本科以下且高中以上者占46.1%, 高中以下者占8.3%。儿童家庭月收入在 1000元以下的占 2.0%, 1000~3000元之间的占21.2%, 3000~6000元之间的占47.9%,6000元以上的占28.9%。
本研究经过山东师范大学伦理委员会审核并获得批准。此外, 本研究在进行数据收集前, 将问卷施测及唾液提取、DNA分型流程等相关信息告知青少年所在学校、青少年监护人及青少年本人,得到三方的知情同意后进行了研究。本研究以班级为单位采集青少年的唾液样本以对 DRD2基因TaqIA多态性进行基因分型, 并通过问卷测评的方式分别测量青少年的抑郁水平及同伴侵害水平。所有采样与施测程序均由经过严格培训的心理学研究生完成。唾液样本采集严格按照规范进行, 要求被试在采样前30分钟不能进食、喝水、嚼口香糖、吸烟, 发高烧 38°C以上的被试此次不予采集, 在采样完成后现场逐个检查样本质量, 对于不符合标准的被试后续进行重新采集。
2.2 研究工具、仪器与材料
2.2.1 同伴侵害
本研究采用Mynard和Joseph编制的多维同伴侵害量表(Multidimensional Peer Victimization Scale,MPVS; Mynard & Joseph, 2000)中的身体侵害、关系侵害两个分量表测量青少年的同伴侵害水平。本研究对选取的身体侵害和关系侵害进行了中文版修订。其中, 身体侵害的分量表包括 3个题目, 如“在这一学期, 别的同学打过我”, 关系侵害分量表包括8个题目, 如“在这一学期, 别的同学故意在背后说我的坏话、造谣”。被试的回答采用4点计分, 0代表“未发生过”, 1代表“很少发生”, 2代表“有时发生”, 3代表“经常发生”, 得分越高说明遭受同伴侵害越严重。本研究中, 身体侵害和关系侵害分量表的Cronbach’s α系数分别为0.79和0.89。为验证身体侵害和关系侵害两个因子的区分效度, 本研究对单因素模型和两因素模型进行比较, 发现 Satorra-Bentler LRT检验显著, △χ2=129.80,df=1,p<0.01, 两因素模型的其他各项指标(χ2=308.27,df=43, RMSEA=0.077, CFI=0.91, TLI=0.88)也优于单因素模型(χ2=477.24,df=44, RMSEA=0.097,CFI=0.85, TLI=0.81), 且两因素模型本身拟合指数良好, 说明身体侵害与关系侵害的区分效度较好。
2.2.2 青少年抑郁
采用儿童抑郁量表(Children’s Depression Inventory,CDI; Kovacs, 1992)测量青少年抑郁情绪。该量表在抑郁相关研究(尤其在非临床样本)中得以广泛应用(Chen, Li, Natsuaki, Leve, & Harold, 2014), 具有良好的信效度。CDI量表共包含27个题目, 要求被试评价近两周内的抑郁水平(如“我偶尔不高兴”, “我经常不高兴”, “我总是不高兴”), 分别采用0、1、2记分, 计算被试的抑郁均分, 得分越高, 表明青少年的抑郁水平越高。本研究中两个时间点青少年抑郁的 Cronbach’s α 系数均为 0.88。
2.2.3 家庭社会经济地位
家庭社会经济地位(SES)由父母文化程度、父母职业及家庭收入合成。其中父母职业根据职业的专业技术性程度将职业归为三类:“农民或下岗失业人员”、“蓝领”和“专业或半专业性人员”, 分别记为1~3分。父母受教育水平包括“小学或小学以下”、“初中(含初中未毕业)”、“高中或中专(含高中未毕业)”、“大专(含夜大、电大)”、“大学本科”和“研究生(硕士或博士)”六类, 每一类分别记1~6分。家庭月收入从“1000元以下”到“8000元以上”每隔 1000元划分为一类, 分别记1~9分。参照有关研究(范兴华, 方晓义, 刘杨, 蔺秀云, 袁晓娇, 2012), 采用因子分析法合成SES指标。SES得分越高, 代表家庭社会经济地位越高。本研究采用合成后的 SES变量作为控制变量。被试的SES平均数为0.00, 标准差为1.00。
2.2.4 DNA样本采集、提取、分型程序
在征得青少年、青少年监护人及合作学校同意的前提下, 以班级为单位采集学生被试的唾液样本,每人采集2~5 ml, 最终共获取有效样本1063个。采用唾液采样的方式提取青少年的DNA样本。利用Sequenom (San Diego, CA, USA)芯片基质辅助激光解吸/电离飞行时间(MALDI-TOF)质谱平台对DRD2基因TaqIA多态性进行基因分型。该多态性的引物为 F: 5’-CGGCTGGCCAAGTTGTCTAA-3’,R: 5’-AGCACCTTCCTGAGTGTCATCA-3’。PCR反应条件为:94°C 15 min; 94°C 20 s, 56°C 30 s,72°C 1 min, 共 45 个循环; 最终 72°C 3 min。随后经历单碱基延伸反应, 并由 MassARRAY Typer软件系统(版本号3.4)完成基因分型分析。
2.3 共同方法偏差的控制
由于本研究中同伴侵害和青少年抑郁均是采用自我报告法收集数据, 可能导致共同方法偏差效应。因此, 根据相关研究的建议(周浩, 龙立荣,2004), 本研究不仅在研究设计与测量过程中采取严格的控制措施, 在数据分析之前, 进一步对共同方法偏差进行统计检验。测量过程中的控制措施具体包括:(1)在数据收集阶段, 本研究所使用的各变量(抑郁, 同伴侵害)均不是接连排版, 并且各问卷的指导语、题目记分的编排方式、量尺不同。(2)选取的同伴侵害和青少年抑郁量表均为信效度较高的成熟量表, 以尽量减少问卷法的局限性给研究结果带来的消极影响; (3)采用严格合理的施测程序减小被试对测量目的的猜度; (4)部分条目使用反向计分。数据分析前, 进一步采用 Harman单因子检验对共同方法偏差进行估计, 结果表明, 特征值大于1的因子共有15个, 且第一个因子解释的变异量为20.03%, 小于40%的临界标准, 说明共同方法偏差不明显。
2.4 数据处理与分析
采用SPSS 22.0进行数据处理与分析。由于身体侵害、关系侵害和抑郁均是正偏态分布(skewness:1.28 ~ 2.45; kurtosis:1.68 ~ 7.17), 可能影响回归的研究结果。参照同类研究(DiLalla, Bersted, & John,2015)对非正态的变量进行了平方根转换, 转换后较好的校正了偏态的问题(skewness:0.06 ~ 0.82;kurtosis:?1.04 ~ 0.13), 随后, 分性别进行分层回归分析, 控制基线抑郁(2009年)和 SES, 分性别考察身体侵害、关系侵害与DRD2基因TaqIA多态性对 2011年时间点青少年抑郁水平的影响。采用这种时间顺序的纵向设计, 能够保证同伴侵害发生在抑郁之前, 并且排除早期抑郁的影响。另外, 为了减少多重共线性问题, 本研究对身体和关系侵害进行标准化处理, 并且不在同一个模型中考察 DRD2基因与两种侵害的交互作用(Benoit, Lacourse, &Claes, 2013)。由于携带A1等位基因的个体大脑纹状体区域的多巴胺 D2型受体密度小于携带 A2A2纯合子个体(Ritchie & Noble, 2003), 本研究参照同类研究的常规做法(van Roekel et al., 2011), 在后续的分析中将A1A1与A1A2基因型携带者进行了合并, 统称为A1等位基因携带者。为控制I类错误,本研究对交互效应的显著性进行 B-H程序校正(Benjamini & Hochberg, 1995)。若交互项显著, 则进一步对两种基因型分别进行分组回归分析。
此外, 由于身体侵害、关系侵害和抑郁均是正偏态分布, 本研究进行了补充分析:将同伴侵害作为类别变量进行分析, 多维同伴侵害量表测量的是个体遭受侵害的频次, 得分为0表示个体从未遭受过侵害(从未遭受过身体侵害的男生占 37%, 女生占51%; 从未遭受过关系侵害的男生占35%, 女生占49%), 得分大于0表示曾遭受过侵害。因此我们将同伴侵害重新编码为 2个类别(受侵害组和无侵害组), 分性别进行 2(同伴侵害:受侵害组 vs.无侵害组) × 2(基因型:A1等位基因vs.A2A2基因型)的方差分析。
3 研究结果
3.1 DRD2基因TaqIA位点的基因型分布
在本研究中, DRD2基因TaqIA位点基因型分布见表 1, 基因型的观测值与期望值吻合良好(χ2=1.18,df=1,p> 0.05), 符合Hardy-Weinberg平衡定律。DRD2基因型分布不存在显著的性别差异(χ2=2.21,df=2,p> 0.05), 将A1A1和A1A2基因型合并后, 基因分布亦不存在性别差异(χ2=0.54,df=1,p> 0.05)。
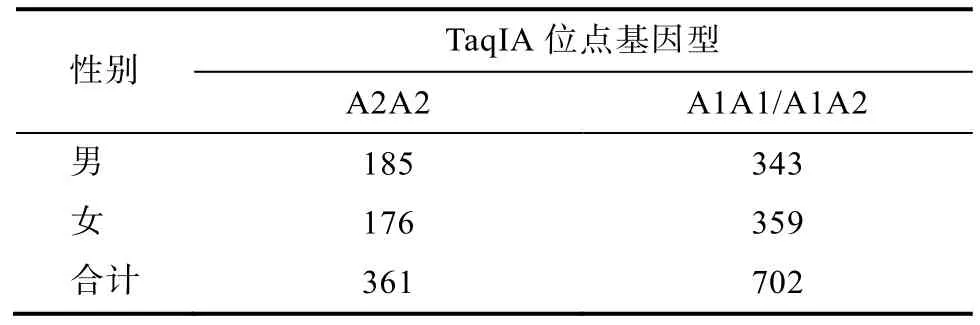
表1 TaqIA多态性位点基因型在男女青少年中的分布

表2 各变量的描述统计量及相关分析结果
3.2 变量的描述统计量与相关分析结果
各变量的平均数、标准差及相关系数见表2。对男女青少年而言, TaqIA多态性与身体侵害和关系侵害的相关均不显著(r=0.03 ~ 0.07,ps > 0.05), 排除了基因环境相关的可能。TaqIA多态性与两个时间点的抑郁症状之间也不存在显著关联(r=? 0.02 ~0.05,ps > 0.05), 表明TaqIA多态性对青少年抑郁无直接效应。同伴身体侵害和关系侵害之间存在显著正相关(r男=0.70,r女=0.51)。身体侵害、关系侵害与青少年两个时间点的抑郁均存在显著正向关联(r=0.15~0.55,ps < 0.001)。两个时间点的抑郁显著正相关(r男=0.49,r女=0.58), 表明青少年抑郁具有一定的稳定性。此外, 独立样本t检验显示,在两个时间点上, 青少年早期抑郁水平均不存在显著的性别差异(ts < 1.72,ps >0.05)。在2009年时间点时, 男青少年的身体侵害(t(1049)=6.46,p<0.001)和关系侵害得分(t(1049)=4.38,p< 0.001)均高于女青少年。

表3 DRD2基因TaqIA多态性与同伴侵害对青少年早期抑郁的影响

图1 DRD2基因TaqIA多态性与身体侵害对男青少年早期抑郁的影响
3.3 DRD2基因TaqIA多态性与同伴侵害对男女青少年早期抑郁的影响
以 2011年时间点抑郁为因变量, 控制基线水平的抑郁和SES, 以TaqIA多态性、同伴身体或者关系侵害, 以及两者的交互项为预测变量分性别进行分层回归分析。结果显示(见表3), 在控制基线水平的抑郁后, 身体侵害和关系侵害不能显著预测青少年的抑郁; TaqIA多态性对青少年早期抑郁亦无显著主效应; TaqIA多态性与同伴身体或关系侵害对男青少年的抑郁具有显著交互作用, 经过B-H统计矫正后结果依然显著; TaqIA多态性与同伴侵害的交互作用对女青少年抑郁的不存在显著的预测作用。
随后, 采用传统的简单斜率检验方法, 对携带不同基因型的男青少年进行分组回归分析(见图1),进一步的分析发现, 在携带A2A2基因型的男青少年中, 同伴身体侵害(b=0.06,t=3.48,p< 0.001)和关系侵害(b=0.05,t=2.69,p< 0.01)可以显著正向预测青少年早期抑郁, 而在携带 A1等位基因的男青少年中, 同伴身体侵害(b=?0.002,t=?0.14,p> 0.05)和关系侵害(b=?0.01,t=?1.04,p> 0.05)对青少年早期抑郁无显著预测作用。
3.4 补充分析
为了进一步检验结果的稳定性, 本研究首先将同伴侵害重新编码为 2个类别(受侵害组和无侵害组), 进行 2(同伴侵害:受侵害组 vs.无侵害组) ×2(基因型:A1等位基因vs.A2A2基因型)的方差分析。结果显示, 在男生中, 同伴身体侵害(F(1, 489)=4.72,p< 0.05, η2=0.01)、关系侵害(F(1, 488)=6.90,p< 0.01, η2=0.01)与基因型的交互效应均显著。进一步的简单效应分析显示, 在A2A2基因型携带者中, 遭受过同伴身体或关系侵害的男生比未遭受过身体或关系侵害的男生抑郁水平高(身体侵害:F(1,494)=11.43,p< 0.01, 关系侵害:F(1, 494)=13.90,p< 0.001)。而在A1等位基因携带者中, 受侵害组与无侵害组的抑郁水平无显著差异(身体侵害:F(1,493)=0.62,p> 0.05; 关系侵害:F(1, 494)=0.26,p>0.05)。在女生中, 同伴身体侵害(F(1, 514)=0.92,p>0.05, η2=0.00)、关系侵害(F(1, 514)=3.48,p> 0.05,η2=0.01)与基因型的交互效应均不显著。这一结果与前述分层回归的结果一致。
4 讨论
本研究通过对 1063名青少年的调查, 发现DRD2基因TaqIA多态性与同伴身体侵害、关系侵害对抑郁存在交互效应并表现出性别差异。具体表现为, 在 A2A2等位基因携带者中, 同伴身体侵害和关系侵害可以显著正向预测男青少年抑郁水平,而在 A1等位基因携带者中, 同伴身体和关系侵害对青少年抑郁无显著的预测作用。但是, TaqIA多态性与身体侵害或关系侵害的交互作用与女青少年的抑郁水平无关。
与前人研究相一致(Crick & Bigbee, 1998;Leadbeater et al., 2014), 本研究相关结果显示身体侵害和关系侵害均与青少年抑郁存在显著正向关联, 即同伴侵害对青少年的情绪适应来说是一种风险性因素, 在一定程度上促进或诱发抑郁情绪的发生。此外, 身体侵害和关系侵害具有中等程度的相关, 这说明身体侵害与关系侵害具有共发性, 遭受身体侵害的个体也更可能遭受关系侵害。此外, 基因对青少年早期抑郁的主效应并未达到统计上的显著性水平, 一方面正如 Molenaar, Middeldorp, van Beijsterveldt和 Boomsma (2015)所指出, 候选基因位点的效应量很小。另一方面可能是因为本研究中选取的均为正常健康青少年, 测量的为抑郁情绪,并不像重性抑郁患者那样具有严重的抑郁或抑郁综合症, 因此其受到遗传影响的比重相对较小。此外, 人类的抑郁情绪可能更多地受到基因与环境间更复杂的交互作用机制的影响, 而非仅仅是单基因的直接效应。
尽管已经有较多研究考察了 DRD2基因与不利环境(如压力性生活事件、父母教养行为等)对个体抑郁的影响(e.g., Elovainio et al., 2007; Hayden et al.,2010), 但是同伴环境与 DRD2基因的交互效应却较少受到关注。本研究采用回归分析和方差分析的结果一致发现无论是身体侵害还是关系侵害, 其与DRD2基因的交互作用仅能够预测男青少年的抑郁水平, 而与女青少年的抑郁症状无关。进一步的分析显示, 只有在携带 A2A2基因型的男青少年中,同伴身体侵害和关系侵害可以显著正向预测其抑郁水平, 而在携带 A1等位基因男青少年中, 同伴侵害对青少年抑郁无显著的预测作用。这提示我们,在这一年龄阶段, 携带A2A2基因型的男青少年可能对同伴侵害更加敏感, 因而表现出较高的抑郁水平。
目前尚未有资料阐明TaqIA多态性A2A2基因型对环境影响更为敏感的内部作用机制, 但是fMRI研究显示, 与A1等位基因携带者相比, 那些携带 A2A2基因型的个体在接受消极反馈刺激时,在后内侧额叶皮层(posterior medial frontal cortex,pMFC)、喙部扣带区域(rostral cingulate zone)表现出较高水平的激活(Klein et al., 2007)。这提示在外界刺激作用下, TaqIA基因可能通过激活与边缘系统相关的脑区, 进而影响抑郁的发生, 由于 A2A2基因型携带者的相关脑区的激活水平更高, 即对外界刺激具有更高的反应性而表现出更高的抑郁风险。此外, 有关情绪控制过程的遗传研究显示, 在处理情绪刺激信息时, DRD2基因rs1076560多态性的GG基因型携带者的前额叶激活水平低于杏仁核(Blasi et al., 2009), 使其在处理情绪刺激时, 前额叶不能对杏仁核的激活进行有效的调控, 导致个体对环境刺激的敏感性增加。虽然目前尚未有研究对TaqIA多态性进行直接考察, 但是从基因表达功能方面来看, 这两种多态性功能具有一定的相似性。rs1076560多态性 GG基因型携带者在前额叶和纹状体均有较多的 DRD2 mRNA表达(Zhang et al.,2007), TaqIA多态性A2A2基因型在纹状体亦具有较高的DRD2受体数量(Ritchie & Noble, 2003)。基于此, 我们推测A2A2基因型携带者中也存在相类似的过程, 外界刺激导致个体的杏仁核激活水平较高, 而前额叶对情绪的调控能力相对滞后导致个体对环境的敏感性增加。
本研究发现A2A2基因型携带者对消极环境更加敏感, 这与前述Elovainio等人(2007)、van Roekel等人(2011)的研究结果相一致。然而, 另一些研究却发现, 在儿童和青少年个体中, A1等位基因携带者对压力性环境更加敏感, 表现出较高水平的抑郁症状(Vaske et al., 2009)或者抑郁相关的认知缺陷(Berman & Noble, 1997)。最近, Zhang等人(2015)对青少年的追踪研究发现, DRD2基因与母亲消极教养的交互效应能够显著预测 11、12岁青少年的抑郁水平, 而在13岁时该交互效应不显著。这一研究结果提示, DRD2基因与环境因素的交互作用可能存在发展动态性。Zhang等人(2015)的研究指出青少年早期向中期的过渡阶段是基因×环境交互作用发生变化的关键期, 在不同的年龄阶段基因对抑郁的效应可能截然相反。由此, 被试年龄差异可能是造成本研究结果与已有研究分歧的原因, 未来研究应该进一步采用追踪设计比较不同发展阶段的DRD2基因与同伴环境的交互效应是否存在发展动态性。
此外, 本研究发现基因×环境交互作用与男青少年抑郁的关联更加密切。一方面, 这种基因×环境效应的性别差异可能源于雌激素对 DRD2基因表达的影响。动物研究发现, 雌二醇水平的上升能够增加DRD2基因mRNA的表达和D2受体的合成(Levavi-Sivan, Biran, & Fireman, 2006)。同时, 生化和行为研究发现雌激素能够调节纹状体多巴胺神经信号的传递, 表现出对多巴胺系统的激活功能(Pohjalainen, Rinne, Någren, Syvälahti, & Hietala,1998)。诸多研究一致表明过低的多巴胺系统活性(如较少的多巴胺受体数量和多巴胺功能降低)与重性抑郁密切相关(Dunlop & Nemeroff, 2007)。由此,雌二醇能够通过调节多巴胺系统活性从而对女性起到保护性作用。相比女性, 男性缺少这种保护性机制而表现出较高的抑郁易感性。另一方面, 这一结果也可能是因为在本研究中女青少年的身体侵害和关系侵害得分和变异均低于男青少年。因此,相比男青少年, 在女生群体中较低的同伴侵害水平和变异可能导致 DRD2基因功能得以表达的机会降低。此外, 也可能本研究测评的同伴侵害并不是女生真正敏感的环境风险因素。如有综述研究指出其他同伴风险环境, 如失去亲密朋友, 对女性抑郁的影响大于男性(Maciejewski, Prigerson, & Mazure,2001)。由此, 未来应该进一步筛选女性敏感的环境变量或者更具压力性的风险环境, 考察其与 DRD2基因的交互效应能否预测女性抑郁。
最后需要指出的是, 本研究仍存在一些局限性。首先, 我们仅选取了两个时间点考察了 DRD2基因与早期同伴侵害对青少年抑郁的纵向影响。有研究显示在青少年早期向中期的过渡期可能是基因功能发展变化的关键期(Zhang et al., 2015)。虽然本研究采用纵向设计, 但并未考察基因功能的变化,且发展阶段仍局限于青少年早期, 该研究结果能否扩展到青少年后期及以后的发展阶段仍需要采用时间跨度更长的追踪研究进行验证。其次, 本研究仅发现了基因×环境对男青少年抑郁发展变化的预测作用, 然而, 这并不意味着女生抑郁的发展变化不受遗传因素的影响。如一些研究显示 MAOA与贫困的交互效应能够预测女性抑郁的发展轨迹, 并且与男性具有相似的基因×环境模式(Marmorstein& Hart, 2011)。另一些研究则发现影响男女抑郁发展的敏感基因型不同(Priess-Groben & Hyde,2013)。这些研究提示不同基因与不同的环境因素对抑郁交互作用的性别差异并不具有一致性。由此,未来研究应该同时采用男性易感基因和女性易感基因, 进一步考察特定遗传基因对两性抑郁的不同影响机制, 探索基因×环境在预测抑郁的强度和方向上是否存在性别差异。第三, 虽然在本研究中DRD2基因TaqIA多态性与两种类型的同伴侵害都不相关, 但是有研究显示同伴侵害具有重要的遗传基础(Beaver et al., 2007), 即同伴侵害是一种受遗传影响的环境因素, 因此本研究结果中可能包含遗传—环境相关带来的混淆, 由此应该更加谨慎的看待本研究发现。此外, 由于本研究测量的是常态青少年群体的抑郁症状, 其结论是否能够推广到临床抑郁症患者中有待于进一步验证。
5 结论
本研究考察了DRD2基因TaqIA多态性与同伴侵害对青少年早期抑郁的交互作用, 结果发现, 在A2A2等位基因携带者中, 同伴身体侵害和关系侵害可以显著正向预测男青少年抑郁水平, 而在 A1等位基因携带者中, 同伴侵害对青少年抑郁无显著的预测作用。此外, DRD2基因与同伴身体、关系侵害对抑郁的交互作用存在性别差异, 该交互效应仅存在于男生中, 而与女青少年抑郁无关。
Avenevoli, S., Swendsen, J., He, J.P., Burstein, M., &Merikangas, K.R.(2015).Major depression in the national comorbidity survey–adolescent supplement: Prevalence,correlates, and treatment.Journal of the American Academy of Child & Adolescent Psychiatry, 54(1), 37–44.e2.
Banny, A.M., Cicchetti, D., Rogosch, F.A., Oshri, A., &Crick, N.R.(2013).Vulnerability to depression: A moderated mediation model of the roles of child maltreatment, peer victimization, and serotonin transporter linked polymorphic region genetic variation among children from low socioeconomic status backgrounds.Development and Psychopathology, 25(3), 599–614.
Beaver, K.M., Wright, J.P., DeLisi, M., Daigle, L.E., Swatt,M.L., & Gibson, C.L.(2007).Evidence of a gene X environment interaction in the creation of victimization:Results from a longitudinal sample of adolescents.International Journal of Offender Therapy and Comparative Criminology, 51(6), 620–645.
Belmaker, R.H., & Agam, G.(2008).Major depressive disorder.New England Journal of Medicine, 358(1), 55–68.
Benjamini, Y., & Hochberg, Y.(1995).Controlling the false discovery rate: A practical and powerful approach to multiple testing.Journal of the Royal Statistical Society.Series B (Methodological), 57(1), 289–300.
Benjet, C., Thompson, R.J., & Gotlib, I.H.(2010).5-HTTLPR moderates the effect of relational peer victimization on depressive symptoms in adolescent girls.Journal of Child Psychology and Psychiatry, 51(2), 173–179.
Benoit, A., Lacourse, E., & Claes, M.(2013).Pubertal timing and depressive symptoms in late adolescence: The moderating role of individual, peer, and parental factors.Development and Psychopathology, 25, 455–471.
Berman, S.M., & Noble, E.P.(1997).The D2dopamine receptor (DRD2) gene and family stress: Interactive effects on cognitive functions in children.Behavior Genetics,27(1), 33–43.
Berman, S.M., Ozkaragoz, T., Noble, E.P., Antolin, T., Sheen,C., Siddarth, P.,...Ritchie, T.(2003).Differential associations of sex and D2dopamine receptor (DRD2) genotype with negative affect and other substance abuse risk markers in children of alcoholics.Alcohol, 30(3), 201–210.
Blasi, G., Lo Bianco, L., Taurisano, P., Gelao, B., Romano, R.,Fazio, L.,...Bertolino, A.(2009).Functional variation of the dopamine D2receptor gene is associated with emotional control as well as brain activity and connectivity during emotion processing in humans.The Journal of Neuroscience, 29(47), 14812–14819.
Blum, K., Chen, A.L.C., Oscar-Berman, M., Chen, T.J.H.,Lubar, J., White, N., … Bailey, J.A.(2011).Generational association studies of dopaminergic genes in reward deficiency syndrome (RDS) subjects: Selecting appropriate phenotypes for reward dependence behaviors.International Journal of Environmental Research and Public Health, 8,4425–4459.
Brummett, B.H., Boyle, S.H., Siegler, I.C., Kuhn, C.M.,Ashley-Koch, A., Jonassaint, C.R.,...Williams, R.B.(2008).Effects of environmental stress and gender on associations among symptoms of depression and the serotonin transporter gene linked polymorphic region(5-HTTLPR).Behavior Genetics, 38(1), 34–43.
Cao, C., Wang, M.P., Ji, L.Q., Wei, X., Cao, Y.M., & Zhang,W.X.(2016).The MAOA rs6323 polymorphism interacts with maternal supportive parenting in predicting adolescent depression: Testing the diathesis-stress and differential susceptibility hypotheses.Acta Psychologica Sinica, 48(1),22–35.
[曹丛, 王美萍, 纪林芹, 魏星, 曹衍淼, 张文新.(2016).MAOA基因rs6323多态性与母亲支持性教养对青少年抑郁的交互作用: 素质–压力假说与不同易感性假说的检验.心理学报, 48(1), 22–35.]
Caspi, A., Sugden, K., Moffitt, T.E., Taylor, A., Craig, I.W.,Harrington, H.,...Poulton, R.(2003).Influence of life stress on depression: Moderation by a polymorphism in the 5-HTT gene.Science, 301(5631), 386–389.
Chen, J., Li, X., & McGue, M.(2012).Interacting effect of BDNF Val66Met polymorphism and stressful life events on adolescent depression.Gene, Brain and Behaviors, 11,958–965.
Chen, J., Li, X.Y., & McGue, M.(2013).The interacting effect of the BDNF Val66Met polymorphism and stressful life events on adolescent depression is not an artifact of gene-environment correlation: Evidence from a longitudinal twin study.Journal of Child Psychology and Psychiatry,54(10), 1066–1073.
Chen, J., Li, X.Y., Natsuaki, M.N., Leve, L.D., & Harold, G.T.(2014).Genetic and environmental influences on depressive symptoms in Chinese adolescents.Behavior Genetics, 44(1), 36–44.
Cole, D.A., Dukewich, T.L., Roeder, K., Sinclair, K.R.,McMillan, J., Will, E.,...Felton, J.W.(2014).Linking peer victimization to the development of depressive self-schemas in children and adolescents.Journal of Abnormal Child Psychology, 42(1), 149–160.
Crick, N.R., & Bigbee, M.A.(1998).Relational and overt forms of peer victimization: A multiinformant approach.Journal of Consulting and Clinical Psychology, 66(2),337–347.
DiLalla, L.F., Bersted, K., & John, S.G.(2015).Peer victimization and DRD4 genotype influence problem behaviors in young children.Journal of Youth and Adolescence, 44(8), 1478–1493.
Dunlop, B.W., & Nemeroff, C.B.(2007).The role of dopamine in the pathophysiology of depression.Archives of General Psychiatry, 64(3), 327–337.
Dunn, E.C., Uddin, M., Subramanian, S.V., Smoller, J.W.,Galea, S., & Koenen, K.C.(2011).Research review: Gene–environment interaction research in youth depression–a systematic review with recommendations for future research.Journal of Child Psychology and Psychiatry,52(12), 1223–1238.
Elovainio, M., Jokela, M., Kivimäki, M., Pulkki-Råback, L.,Lehtimäki, T., Airla, N., & Keltikangas-Järvinen, L.(2007).Genetic variants in the DRD2gene moderate the relationship between stressful life events and depressive symptoms in adults: Cardiovascular risk in young Finns study.Psychosomatic Medicine, 69, 391–395.
Fan, X.H., Fang, X.Y., Liu, Y., Lin, X.Y., & Yuan, X.J.(2012).The effect of social support and social identity on the relationship between perceived discrimination and socio-cultural adjustment among Chinese migrant children.Acta Psychologica Sinica, 44(5), 647–663.
[范兴华, 方晓义, 刘杨, 蔺秀云, 袁晓娇.(2012).流动儿童歧视知觉与社会文化适应: 社会支持和社会认同的作用.心理学报, 44(5), 647–663.]
Goodnight, J.A., Bates, J.E., Newman, J.P., Dodge, K.A., &Pettit, G.S.(2006).The interactive influences of friend deviance and reward dominance on the development of externalizing behavior during middle adolescence.Journal of Abnormal Child Psychology, 34, 573–583.
Hankin, B.L.(2015).Depression from childhood through adolescence: Risk mechanisms across multiple systems and levels of analysis.Current Opinion in Psychology, 4, 13–20.Hawker, D.S.J., & Boulton, M.J.(2000).Twenty years'research on peer victimization and psychosocial maladjustment: A meta-analytic review of cross-sectional studies.Journal of Child Psychology and Psychiatry, 41(4), 441–455.
Hayden, E.P., Klein, D.N., Dougherty, L.R., Olino, T.M.,Laptook, R.S., Dyson, M.W.,...Singh, S.M.(2010).The dopamine D2receptor gene and depressive and anxious symptoms in childhood: Associations and evidence for gene–environment correlation and gene–environment interaction.Psychiatric Genetics, 20(6), 304–310.
Iyer, P.A., Dougall, A.L., & Jensen-Campbell, L.A.(2013).Are some adolescents differentially susceptible to the influence of bullying on depression?Journal of Research in Personality, 47(4), 272–281.
Klein, T.A., Neumann, J., Reuter, M., Hennig, J., von Cramon,D.Y., & Ullsperger, M.(2007).Genetically determined differences in learning from errors.Science, 318(5856),1642–1645.
Kovacs, M.(1992).Children’s depression inventory (CDI)manual.Toronto, Canada: Multi-Health Systems Inc.
Leadbeater, B.J., Thompson, K., & Sukhawathanakul, P.(2014).It gets better or does it? Peer victimization and internalizing problems in the transition to young adulthood.Development and Psychopathology, 26(3), 675–688.
Lépine, J.P., & Briley, M.(2011).The increasing burden of depression.Neuropsychiatric Disease and Treatment,7(Suppl 1), 3–7.
Levavi-Sivan, B., Biran, J., & Fireman, E.(2006).Sex steroids are involved in the regulation of gonadotropin-releasing hormone and dopamine D2receptors in female tilapia pituitary.Biology of Reproduction, 75(4), 642–650.
Liberg, B., Adler, M., Jonsson, T., Landén, M., Rahm, C.,Wahlund, L.O.,...Wahlund, B.(2013).The neural correlates of self-paced finger tapping in bipolar depression with motor retardation.Acta Neuropsychiatrica, 25(1), 43–51.
Maciejewski, P.K., Prigerson, H.G., & Mazure, C.M.(2001).Sex differences in event-related risk for major depression.Psychological Medicine, 31(4), 593–604.
Marmorstein, N.R., & Hart, D.(2011).Interactions between MAOA genotype and receipt of public assistance:Predicting change in depressive symptoms and body mass index.Journal of Research on Adolescence, 21(3),619–630.
McLeod, G.F.H., Horwood, L.J., & Fergusson, D.M.(2016).Adolescent depression, adult mental health and psychosocial outcomes at 30 and 35 years.Psychological Medicine, 46(7), 1401–1412.
Mihara, K., Kondo, T., Suzuki, A., Yasui-Furukori, N., Ono, S.,Sano, A.,...Kaneko, S.(2003).Relationship between functional dopamine D2and D3receptors gene polymorphisms and neuroleptic malignant syndrome.American Journal of Medical Genetics Part B: Neuropsychiatric Genetics,117(1), 57–60.
Molenaar, D., Middeldorp, C., van Beijsterveldt, T., & Boomsma,D.I.(2015).Analysis of behavioral and emotional problems in children highlights the role of genotype ×environment interaction.Child Development, 86(6), 1999–2016.
Mynard, H., & Joseph, S.(2000).Development of the multidimensional peer-victimization scale.Aggressive Behavior, 26(2), 169–178.
Natsuaki, M.N., Biehl, M.C., & Ge, X.J.(2009).Trajectories of depressed mood from early adolescence to young adulthood: The effects of pubertal timing and adolescent dating.Journal of Research on Adolescence, 19(1), 47–74.
Ozkaragoz, T., & Noble, E.P.(2000).Extraversion: Interaction between D2dopamine receptor polymorphisms and parental alcoholism.Alcohol, 22(3), 139–146.
Perren, S., Ettekal, I., & Ladd, G.(2013).The impact of peer victimization on later maladjustment: Mediating and moderating effects of hostile and self-blaming attributions.Journal of Child Psychology and Psychiatry, 54(1), 46–55.
Pohjalainen, T., Rinne, J.O., Någren, K., Syvälahti, E., &Hietala, J.(1998).Sex differences in the striatal dopamine D2receptor binding characteristics in vivo.American Journal of Psychiatry, 155(6), 768–773.
Priess-Groben, H.A., & Hyde, J.S.(2013).5-HTTLPR X stress in adolescent depression: Moderation by MAOA and gender.Journal of Abnormal Child Psychology, 41(2),281–294.
Reijntjes, A., Kamphuis, J.H., Prinzie, P., & Telch, M.J.(2010).Peer victimization and internalizing problems in children: A meta-analysis of longitudinal studies.Child Abuse & Neglect, 34(4), 244–252.
Ritchie, T., & Noble, E.P.(2003).Association of seven polymorphisms of the D2dopamine receptor gene with brain receptor-binding characteristics.Neurochemical Research, 28(1), 73–82.
Schwartz, D., Lansford, J.E., Dodge, K.A., Pettit, G.S., &Bates, J.E.(2015).Peer victimization during middle childhood as a lead indicator of internalizing problems and diagnostic outcomes in late adolescence.Journal of Clinical Child & Adolescent Psychology, 44(3), 393–404.
Steinberg, L.(2014).Adolescence(10th ed.).New York:McGraw-Hill.
Sugden, K., Arseneault, L., Harrington, H., Moffitt, T.E., Williams,B., & Caspi, A.(2010).Serotonin transporter gene moderates the development of emotional problems among children following bullying victimization.Journal of the American Academy of Child & Adolescent Psychiatry,49(8), 830–840.
Sugimura, N., & Rudolph, K.D.(2012).Temperamental differences in children's reactions to peer victimization.Journal of Clinical Child & Adolescent Psychology, 41(3),314–328.
Troop-Gordon, W., & Ladd, G.W.(2005).Trajectories of peer victimization and perceptions of the self and schoolmates:Precursors to internalizing and externalizing problems.Child Development, 76(5), 1072–1091.
Twenge, J.M., & Nolen-Hoeksema, S.(2002).Age, gender,race, socioeconomic status, and birth cohort difference on the children's depression inventory: A meta-analysis.Journal of Abnormal Psychology, 111(4), 578–588.
van Roekel, E., Goossens, L., Scholte, R.H.J., Engels, R.C.M.E., & Verhagen, M.(2011).The dopamine D2receptor gene, perceived parental support, and adolescent loneliness:Longitudinal evidence for gene–environment interactions.Journal of Child Psychology and Psychiatry, 52(10),1044–1051.
Vaske, J., Makarios, M., Boisvert, D., Beaver, K.M., & Wright,J.P.(2009).The interaction ofDRD2and violent victimization on depression: An analysis by gender and race.Journal of Affective Disorders, 112(1-3), 120–125.
Verboom, C.E., Sijtsema, J.J., Verhulst, F.C., Penninx, B.W.J.H., & Ormel, J.(2014).Longitudinal associations between depressive problems, academic performance, and social functioning in adolescent boys and girls.Developmental Psychology, 50(1), 247–257.
Watson, D., Stasik, S.M., Ellickson-Larew, S., & Stanton, K.(2015).Extraversion and psychopathology: A facet-level analysis.Journal of Abnormal Psychology, 124(2),432–446.
Zhang, L.L., Li, Z., Chen, J., Li, X.Y., Zhang, J.X., & Belsky, J.(2016).The BDNF Val66Met polymorphism interacts with maternal parenting influencing adolescent depressive symptoms: Evidence of differential susceptibility model.Journal of Youth and Adolescence,45(3), 471–483.
Zhang, W.X., Cao, Y.M., Wang, M.P., Ji, L.Q., Chen, L., &Deater-Deckard, K.(2015).The dopamine D2receptor Polymorphism (DRD2 TaqIA) interacts with maternal parenting in predicting early adolescent depressive symptoms: Evidence of differential susceptibility and age differences.Journal of Youth and Adolescence, 44(7),1428–1440.
Zhang, Y., Bertolino, A., Fazio, L., Blasi, G., Rampino, A.,Romano, R.,...Sadée, W.(2007).Polymorphisms in human dopamine D2receptor gene affect gene expression,splicing, and neuronal activity during working memory.Proceedings of the National Academy of Sciences of the United States of America, 104(51), 20552–20557.
Zhou, H., & Long, L.R.(2004).Statistical remedies for common method biases.Advances in Psychological Science,12(6), 942–950.
[周浩, 龙立荣.(2004).共同方法偏差的统计检验与控制方法.心理科学进展, 12(6), 942–950.]
