甘蓝型油菜PEPC基因ihpRNA表达载体的构建与遗传转化研究
2017-01-17谭太龙邬贤梦官春云熊兴华
邢 蔓,谭太龙,李 健,邬贤梦,官春云,熊兴华
(1.湖南农业大学 作物基因工程湖南省重点实验室,湖南 长沙 410128;2.国家油料改良中心湖南分中心,湖南 长沙 410128)
甘蓝型油菜PEPC基因ihpRNA表达载体的构建与遗传转化研究
邢 蔓1,2,谭太龙2,李 健2,邬贤梦2,官春云2,熊兴华1,2
(1.湖南农业大学 作物基因工程湖南省重点实验室,湖南 长沙 410128;2.国家油料改良中心湖南分中心,湖南 长沙 410128)
为提高油菜含油量,构建了基于油菜磷酸烯醇式丙酮酸羧化酶基因(PEPC)的ihpRNA植物表达载体。分别从甘蓝型油菜基因组中克隆了种子储藏蛋白napin基因启动子(315 bp)、PEPC基因的外显子片段(246 bp)及PEPC基因的内含子(754 bp),包含了其剪切位点之前4 bp和之后18 bp片段。通过中间载体pBluescript Ⅱ SK(+)再构建到pBI121中,构成种子特异表达的内含子剪接的PEPC基因的RNA 干扰载体pBI121.NP+IP-。利用农杆菌介导法对湘油15号进行遗传转化,获得卡那霉素抗性转化植株9株,PCR检测证实外源片段成功导入其中7株油菜基因组中。对T2种子中含油量检测发现比对照提高10%左右。
甘蓝型油菜;PEPC基因;ihpRNA载体;转基因油菜;含油量
油菜是我国重要的油料作物,年产油400万t,占植物油生产量的1/3,每年还需进口植物油1 000万t。然而,我国多数品种含油量为40%左右,比进口菜籽的含油量低3%~5%[1],提高油菜含油量一直是油菜育种研究的主要目标之一,也是当前国际上油料作物基因工程研究的一个热点[2]。

本试验依据ihpRNA介导的RNAi技术原理,分别克隆了napin启动子、PEPC基因片段及其内含子,构建了对应于油菜PEPC基因的ihpRNA植物表达载体,通过遗传转化以抑制油菜PEPC基因在种子中的表达。转基因油菜含油量提高了10%,为高油油菜分子育种提供研究材料。
1 材料和方法
1.1 植物材料
甘蓝型油菜(BrassicanapusL.)湘油15号,由湖南农业大学国家油料作物改良中心湖南分中心提供。
1.2 质粒、菌种、酶和试剂
pBI121、大肠杆菌菌种DH5α、农杆菌LBA4404、pBluescript Ⅱ SK(+)由湖南农业大学作物基因工程重点实验室提供。TaqDNA聚合酶、pGEM-T Easy、各种限制性内切酶、T4 DNA连接酶、dNTP均购自TaKaRa公司。质粒提取试剂盒和DNA快速纯化/回收试剂盒均购自北京鼎国生物公司,其余生化试剂均为进口或国产分析纯试剂。引物由丽欣生物公司合成,万百生物公司完成测序。其余生化试剂均为进口或国产分析纯试剂。
1.3 PCR引物
根据GenBank中已发表的油菜相应基因序列分别设计PCR引物,并在其5′端引入相应的酶切位点(下划线为引物酶切位点序列;斜体代表PEPC内含子剪切位点之前和之后的序列)。napin上游引物(FWn) AAGCTTTAGACTCTCATCCCCTTTTA,napin下游引物(RVn) GAATTCTGAGTAAAGAGTG AAGCGG;PEPC+上游引物(FWp+)GAATTCGTGGC TGGTCACAAGGA,PEPC+下游引物(RVp+)GGATCC AGTGTCTTCAAGTCCAGGT;PEPC-上游引物(FWp-) GAGCTCGTGGCTGGTCACAAGGA,PEPC-下游引物(RVp-) TCTAGAAGTGTCTTCAAGTCCA GGT;内含子上游引物(FWi)GGATCCATTTGTTACC GTTTCCCTTT,内含子下游引物(RVi)TCTAGACTCGTAGCATTCTTGAACCTT。
1.4 油菜napin启动子、PEPC内含子及正、反基因片段序列克隆与测序油菜DNA PCR扩增:50 μL体系扩增基因,反应体系包括:基因组DNA 1 μL、2.5 mmol/L dNTP 5 μL、10×PCR Buffer 5 μL、Taq酶2 μL、含5 pmol/μL引物各2 μL,加水至50 μL。PCR程序为94 ℃预变性3 min;94 ℃变性1 min,50 ℃退火1 min,72 ℃延伸90 s,35个循环。 PCR产物与pGEM-T Easy连接参照产品说明书进行。大肠杆菌DH5α感受态细胞的制备、转化、重组质粒DNA的提取与酶切均按常规方法进行。经Hind Ⅲ和EcoRⅠ酶切鉴定napin启动子;用EcoR Ⅰ和BamH Ⅰ以及SacⅠ和XbaⅠ进行酶切鉴定246 bp正向与反向基因片段;用BamH Ⅰ+XbaⅠ酶切鉴定PEPC内含子片段。鉴定的阳性克隆送万百生物公司测序。将各基因测序结果分别在GenBank 中进行序列的同源性比较。
1.5 种子特异性表达的ihpRNA植物载体pBI121.NP+IP-的构建将双酶切片段分别回收。首先将回收的napin启动子插入pBluescript Ⅱ SK(+)的Hind Ⅲ和EcoRⅠ位点构建pSK.N;再将用EcoR Ⅰ和BamH Ⅰ双酶切回收的PEPC正向基因片段(PEPC+)插入pSK.N的相同酶切位点上构建pSK.NP+;然后回收pSK.NP+经Hind Ⅲ+BamH Ⅰ酶切产生561 bp 的napin基因启动子和PEPC+片段插入pBI121的同样酶切位点构建pBI121.NP+;将BamH Ⅰ+XbaⅠ酶切产生754 bp 的PEPC内含子片段插入pBI121.NP+的同样酶切位点构建pBI121.NP+Ⅰ;最后经SacⅠ和XbaⅠ酶切产生246 bp 的PEPC基因反向片段(PEPC-)插入pBI121.NP+I的同样酶切位点上构建pBI121.NP+IP-(图1)。

图1 PEPC基因ihpRNA载体示意图Fig.1 The schematic diagram of ihpRNA vector targeting PEPC gene
1.6 甘蓝型油菜的遗传转化
将植物表达载体pBI121.NP+IP-通过冻融法转入农杆菌LBA4404,菌落PCR鉴定。利用农杆菌介导法,以湘油15号的子叶柄为外植体进行遗传转化。
1.7 转基因油菜T0的PCR检测
取非转基因再生油菜苗和转基因抗卡那霉素油菜苗分别提取总DNA,利用napin启动子上游引物和内含子的下游引物,PCR扩增检测ihpRNA-PEPC。
1.8 转基因油菜后代的含油量检测
从转基因后代中选择含油量高的材料进行自交,T2种子进行含油量检测。采取标准索氏抽提法的提取步骤参照中华人民共和国国家标准GB2906-82。测定重复3次,对结果进行统计分析。
2 结果与分析
2.1napin基因启动子、PEPC基因正反义序列及内含子的克隆提取湘油15叶片总DNA,分别利用引物FWn/RVn、FWp+/RVp+、FWp-/RVp-、FWi/RVi PCR克隆napin基因启动子、PEPC基因正、反义链片段和内含子序列。经1%琼脂糖凝胶电泳分析表明:PCR产物大小均与设计的序列大小相符(图2-4)。将扩增产物回收后分别与T-Easy相连,获得重组质粒。分别经Hind Ⅲ/EcoR Ⅰ、EcoR Ⅰ/BamH Ⅰ、SacⅠ/XbaⅠ 和BamH Ⅰ/XbaⅠ双酶切鉴定后进行测序。测序结果表明:napin启动子为315 bp(图2),与GenBank中油菜1.7 Snapin、napB的启动子序列同源性达到100%,与2Snapin启动子同源性达到了99%。PEPC基因正、反义链片段大小为246 bp(图3),与GenBank中PEPC3序列(D13987)的同源性达到95%以上。PEPC内含子以及其剪接位点上游4 bp 和下游18 bp的序列长度为754 bp(图4),与GenBank中的PEPC序列完全一致。

M.100 bp DNA Marker;1~2.napin启动子片段。M.100 bp DNA Marker;1-2.napin promoter fragments.

M.100 bp DNA Marker;1~2.PEPC+正向片段;3~4.PEPC-反向片段。 M.100 bp DNA Marker;1-2.PEPC+ sense fragments;3-4.PEPC- anti-sense fragments.

M.100 bp DNA Marker;1.PEPC内含子片段。M.100 bp DNA Marker;1.PEPC intron fragments.
2.2 种子特异性表达的PEPCihpRNA的植物载体构建
分别将双酶切的4个片段回收。 首先将napin启动子插入pBluescript Ⅱ SK(+)的Hind Ⅲ和EcoR Ⅰ位点构建pSK.N;再将回收的PEPC+正向基因片段插入pSK.N的相同酶切位点上构建pSK.NP+(图5);然后将napin基因启动子和PEPC+正向片段插入pBI121的同样酶切位点构建pBI121.NP+(图6);再依次插入内含子片段和反向片段而构建pBI121.NP+I(图7)和pBI121.NP+IP-(图8)。每个中间载体分别进行酶切鉴定和PCR鉴定。对pBI121.NP+IP-分别进行napin、NP+(napin与PEPC+)、intron、IP-(intron与PEPC-)的 PCR鉴定(图8)。

M.DNA Marker Ⅲ;1~2. Hind Ⅲ和EcoRⅠ酶切pSK.NP+。M.DNA Marker Ⅲ;1-2. Hind Ⅲ and EcoRⅠdigested pSK.NP+.

M.DNA Marker Ⅲ;1. Hind Ⅲ和BamH Ⅰ酶切pBI121.NP+。M.DNA Marker Ⅲ;1. Hind Ⅲ and BamH Ⅰ digested pBI121.NP+.

M.DNA Marker Ⅲ;1. BamH Ⅰ和Xba Ⅰ双酶切pBI121.NP+Ⅰ。M.DNA Marker Ⅲ;1. BamH Ⅰ and Xba Ⅰ digested pBI121.NP+Ⅰ.

M.DNA Marker Ⅲ;1.napin启动子;2.napin和PEPC+片段; 3.PEPC 内含子;4.intron 和PEPC-片段。M.DNA Marker Ⅲ;1.napin;2.NP+;3.PEPC intron;4.IP-.
2.3 农杆菌介导pBI121.NP+IP-载体转化湘油15
在培养基上取发芽5 d左右的湘油15幼苗带生长点的子叶节,预培养1 d。经取OD值为0.5左右的农杆菌LBA4404(含pBI121.NP+IP-表达载体)浸染7 min后,共培养在共培养基上暗培养3 d,再转入放入Kan筛选培养基进行Kan 梯度筛选分化培养(先10 mg/L,后提高到15 mg/L,30 d继代1次,继代2次)。待筛选后的绿芽长到2~3 cm时,切下绿芽转入生根培养基诱导生根。当根系发达成为完整的植株后,经炼苗便可移栽至营养钵中(图9)。
2.4 转基因油菜T0的PCR检测
以napin启动子的FW引物与内含子的RW组成引物对,对9株再生转ihpRNA-PEPC抗性植株进行PCR检测,扩增得到产物为1 300 bp左右。检测结果发现:9株再生植株中能扩增出目的条带的植株有7株(图10),PCR阳性率为77%。
2.5 转基因油菜后代的含油量检测
利用索氏提取法测定对照植株与转基因T2植株种子含油量,结果如图11 所示:5个转基因株系种子含油量都明显高于对照植株,说明通过RNAi技术干扰种子内源PEPC,并能提高转基因植株种子中油分积累。
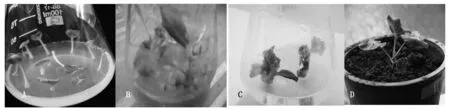
A.子叶节预培养;B.子叶节筛选培养; C.在生根培养基中的完整植株;D.在花盆中生长的转基因植株。A.Cotyledon node preculture;B.Cotyledon node on selection medium; C.Whole plant on rooting medium;D.Transgenic plants in pots.

M.DNA Marker Ⅲ;1.负对照;2.正对照;3~6,8~10.转基因植株;7,11.非转基因植株。M.DNA Marker Ⅲ;1.Negative control;2.Positive control;3-6,8-10.Transgenic plants;7,11.Non-transgenic plant.
3 讨论
熊兴华等[14]曾从甘蓝型油菜中克隆了1 147 bp的napin启动子,谭小力等[15]对此启动子在拟南芥中表达进行了功能验证,此napin启动子控制基因只在和种子有关的部位和种子衍生的部位表达。Stålberg等[16]在转基因烟草中对甘蓝型油菜种子储藏蛋白napA基因的启动子进行了研究,发现napin启动子具有正、负调控元件。删除-1 101~-309 bp可增强GUS的表达,删除-309~-211 bp 降低其表达,从-152~+44 bp是种子特异表达所必需的最小序列,而5′端删除到-126 bp则无启动子功能。

CK.非转基因油菜(湘油15号);T2-1~T2-5.转基因植株。CK.Non-transgenic plant(Xiangyou 15);T2-1-T2-5.Transgenic plants.
本试验依据napA基因的启动子序列克隆了315 bp的napin启动子序列(-310~+5 bp),测序结果完全一致,可以在油菜种子中实现超强表达。
陈锦清等[4]转35S启动子驱动的反义PEPC基因在油菜中表达使其含油量达50%以上。张艳等[17]将高粱PEPC基因转入大豆,T1植株的净光合速率提高了20.7%,气孔导度和蒸腾速率均有不同程度的提高。陈全战等[18]将玉米PEPC基因导入甘蓝型油菜,PEPC活性提高17倍,净光合速率提高15.5%。Wang等[19]将玉米的PEPC基因导入小麦,提高了转化小麦的耐旱性和千粒质量。王永霞等[20]将玉米PEPC基因导入小麦后,促进了小麦原有的C4循环,提高了小麦的光合效率和籽粒产量。
目前,构建ihpRNA表达载体用于特定基因表达调控的研究应用广泛,而如何构建高效载体则是其中关键的一步。首先,本试验选择PEPC基因的编码区作为靶标序列构建载体。笔者根据GenBank中序列,设计引物,直接从甘蓝型油菜湘油15基因组DNA中克隆PEPC基因片段,正向和反向片段在转录后形成的mRNA严格互补且与内源靶序列一致。其次,本试验所构建的RNAi植物表达载体是1个ihpRNA(Intron splicing hpRNA,ihpRNA)结构,研究表明,含内含子剪接的反转重复结构的沉默效果优于hpRNA。构建的pBI121.NP+IP-是先插入正向片段,再插入反向片段;此外,在克隆内含子片段时选择了从其剪接位点前4 bp和剪接位点后面的18 bp克隆,有利于形成更稳定的发夹结构,增强其沉默效果。
为满足人们对食用油消费不断增加和生产生物柴油的需要,增加油料作物种子油的产量成为油料作物研究的主要目标,前人的研究证实通过操纵油合成途径的基因只可适当提高含油量。目前,我们正在进行利用转录因子基因和油合成关键基因来调控油菜含油量的研究。
[1] 傅寿仲,张洁夫,戚存扣,等.甘蓝型油菜高含油量种质选育研究[J].中国油料作物学报,2008,30(3):279-283.
[2] Detlef H,Carsten O,Ludger A,et al.Breading progress towards high oil content in oilseed rape(BrassicanapusL.)essential innovations to meet current and future market needs[C]//Wuhan:Proceedings The 12th International Rapeseed Congress,2007:159-162.
[3] Khaled S,Zeineb O,Habib K,et al.Evolution of phosphoenolpyruvate carboxylase activity and lipid content during seed maturation of two spring rapeseed cultivars(BrassicanapusL.) [J].C R Biologies,2006,329(9):719-725.
[4] 陈锦清,郎春秀.反义PEP基因调空油菜籽粒蛋白质/油脂含量比率的研究[J].农业生物技术学报,1999,7(4):316-320.
[5] 夏 军,郑明刚,王 玲,等.运用CRISPR/Cas系统敲除大肠杆菌磷酸烯醇式丙酮酸羧化酶基因及其对脂肪酸代谢的影响[J].微生物学通报,2016,43(8):1864-1871.
[6] 田琪琳,施定基,贾晓会,等.“细菌型”pepc2 基因反向表达载体构建及在莱茵衣藻中的表达[J].上海海洋大学学报,2013,22(5):665-671.
[7] Mamedovt T G,Chollet R.Discovery of novel Phosphoenolpyruvate Carboxylase ( PEPC ) genes and their active polypeptides in the green microalgaChlamydomonasreinhardtii[J].Biological Sciences,2010,65(5/6):99-105.
[8] 王岳坤,阳江华.橡胶树磷酸烯醇式丙酮酸羧化酶基因HbPPC1 的克隆和表达分析[J].热带作物学报,2015,36(5):895-900.
[9] 冯瑞云,白云凤,李 平,等.籽粒苋C4型磷酸烯醇式丙酮酸羧化酶基因的克隆和表达[J].作物学报,2011,37(10):1801-1808.
[10] 吴 琼,许为刚,李 艳,等.田间条件下转玉米C4 型PEPC基因小麦的光和生理特性[J].作物学报,2011,37(11):2046-2052.
[11] 潘丽娟,刘风珍,万勇善,等.PEPC基因在高油品种吉花4号及其亲本中的表达比较[J].花生学报,2015,44(4):37-41.
[12] 王志慧,童超波,袁午舟.四种油料作物中的细菌型PEPC基因的鉴定及在发育种子中的表达[J].中国油料作物学报,2013,35(1):8-16.
[13] 田保明.基于RNAi技术的BnFAE1基因沉默及对甘蓝型油菜芥酸合成的影响[D].武汉:华中农业大学,2010.
[14] 熊兴华,官春云,李 恂,等.油菜种子特异表达napin基因启动子的克隆及序列分析[J].生物技术,2003,13(3):4-5.
[15] 谭小力,张志燕,伍 娟,等.油菜种子特异表达启动子napin的功能鉴定[J].中国油料作物学报,2005,27(4):85-88.
[16] Stålberg K,Ellerström M,Josefsson L G,et al.Deletion analysis of a 2S seed storage protein promoter ofBrassicanapus in transgenic tobacco[J].Plant Molecular Biology,1993,23(4):671-683.
[17] 张 艳,满为群,南相日.高粱C4型PEPC基因转入大豆可改善大豆光合特性[J].分子植物育种,2015,13(2):294-300.
[18] 陈全战,张边江,吴 梅.转pepc基因油菜及其光合生理表现[J].西北农业学报,2013,22(8):42-46.
[19] Wang F L,Liu R H,Wu G T,et al.Specific downregulation of the Bacterial-TypePEPCgene by artificial MicroRNA improves salt tolerance inArabidopsis[J].Plant Molecular Biology Reporter,2012,30(5):1080-1087.
[20] 王永霞,许为钢,胡 琳,等.外源玉米PEPC基因对小麦C3、C4途径相关基因表达的效应[J].麦类作物学报,2016,36(1):1-8.
Construction of Seed-specific ihpRNA Expression Vector Targeting PEPC Gene and Transformation of Rapeseed
XING Man1,2,TAN Tailong2,LI Jian2,WU Xianmeng2,GUAN Chunyun2,XIONG Xinghua1,2
(1.Crop Gene Engineering Key Laboratory of Hunan Province,Hunan Agricultural University,Changsha 410128, China;2.Hunan Branch of National Oilseed Crops Improvement Center,Changsha 410128,China)
To study the relationship between rapeseed oil content and phosphoenolpyruvate carboxylase activity,we constructed the seed-specific ihpRNA expression vector targetingPEPCgene inBrassicanapusL..A fragment of 315 bpnapinpromoter and 246 bp exon fragment ofPEPCgene were amplified from the genomic DNA of Xiangyou 15.At the same time,a intron(754 bp) ofPEPCgene including 4 bp and 18 bp DNA fragment before and after the splice site respectively was cloned into corresponding enzyme sites.According to the structural principle of expression vector,these fragments were cloned into pBluescript Ⅱ SK(+),respectively.Then we inserted the ihpRNA structure into plant expression vector pBI121,designated pBI121.NP+IP-.Transformed Xiangyou 15 byAgrobacteriumtumefacienscontaining pBI121.NP+IP-,seven transgenic plants from nine anti-kanamycin plants were confirmed by PCR detection.The oil content of transgenic plants was about 10% higher than that of the controls.
BrassicanapusL.;PEPCgene;ihpRNA expression vector;Transgenic rapeseed;Oil content
2016-07-15
国家“973”计划项目(2015CB150200);湖南省科技计划项目(2014FJ1006-3)
邢 蔓(1992-),男,新疆伊犁人,在读硕士,主要从事油菜分子生物学研究。
熊兴华(1967-),男,湖南安乡人,副研究员,博士,主要从事油菜分子生物学及基因工程研究。
Q78;S635.03
A
1000-7091(2016)06-0007-05
10.7668/hbnxb.2016.06.002
