蛋白质代谢通路对鸡雄性生殖细胞分化的调控
2017-01-13汤贝贝王颖洁纪艳芹路镇宇张亚妮李碧春
李 东,汤贝贝,王颖洁,纪艳芹,王 飞,路镇宇,王 曼,张亚妮,李碧春
(扬州大学动物科学与技术学院/江苏省动物繁育与分子设计重点实验室,江苏扬州225009)
蛋白质代谢通路对鸡雄性生殖细胞分化的调控
李 东,汤贝贝,王颖洁,纪艳芹,王 飞,路镇宇,王 曼,张亚妮,李碧春
(扬州大学动物科学与技术学院/江苏省动物繁育与分子设计重点实验室,江苏扬州225009)
【目的】探究蛋白质代谢在鸡雄性生殖细胞分化过程中的作用机制,为完善鸡胚胎干细胞(embryonic stem cell,ESC)体外向雄性生殖细胞诱导分化体系研究提供依据。【方法】采用流式分选的方法获取高纯度的ESC、原始生殖细胞(primordial germ cells,PGCs)和精原干细胞(spermatogonia stem cell,SSCs),分别提取细胞的总 RNA,采用 RNA-Seq方法对 3种细胞的转录本进行深度测序,然后进行 WEGO(web gene ontology)和KEGG(kyoto encyclopedia of genes and genomes)通路富集分析,筛选出鸡雄性生殖细胞分化过程中参与蛋白质代谢的关键通路及其关键基因,RT-qPCR (Real time Quantitative PCR) 检测部分关键差异基因的表达变化,并与RNA-Seq(RNA sequencing)结果进行比较分析,同时分别从体内和体外水平对关键基因NOS2进行抑制,观察各分组不同天数的细胞形态变化及检测NOS2和C-kit、Cvh、Stra8、Dazl、integrin α6和integrin β1等生殖标记基因表达变化情况。【结果】在雄性生殖细胞分化的整个阶段,有697个差异基因参与生物代谢,显著富集于精氨酸-脯氨酸代谢通路、酪氨酸代谢通路以及色氨酸代谢通路,在这3条通路上筛选出NOS2、FAH和IDO等关键性基因,这些基因的在ESCs向SSCs分化过程中表达变化趋势与其在RNA-Seq中的结果一致。体内抑制NOS2基因后,NOS2及C-kit、Cvh、Stra8、Dazl、integrin α6和integrin β1等生殖标记基因在空白组和对照组之间无显著性的差异,而在抑制剂组中,NOS2及C-kit、Cvh、Stra8、Dazl、integrin α6和integrin β1等生殖标记基因的mRNA表达量均出现了降低;而体外抑制NOS2基因后,对照组中的ESCs在2、4、6、8和10d内细胞不断增殖,但是未出现类胚体;RA诱导组中,2d出现小的类胚体,4d类胚体增大,且数量增多,6d类胚体边缘开始出现破裂,8d类胚体解体,10d出现类精原样细胞;抑制剂组中,ESCs在 2、4、6、8和 10d内无类胚体出现,且相较于对照组细胞增殖缓慢;RA+抑制剂组中,2和4d内无类胚体出现,细胞增殖缓慢,6d出现小的类胚体,8d类胚体数量少量增多,且体积稍显增大,10d类胚体开始裂解。并且经过抑制剂的抑制后,NOS2及C-kit、Cvh、Stra8、Dazl、integrin α6和integrin β 1等生殖标记基因的表达量在RA诱导组、抑制剂组和RA+抑制剂组中相对于对照组均呈显著性或极显著性的下调趋势。【结论】基于RNA-Seq技术及生物信息学方法筛选出ESCs向雄性生殖细胞分化过程中精氨酸-脯氨酸代谢通路及关键基因NOS2的基础上,通过对NOS2基因在鸡体内和体外的抑制,发现NOS2在被抑制后,ESCs向雄性生殖细胞分化的过程受到抑制。说明了精氨酸-脯氨酸代谢通路及关键基因NOS2对ESCs向雄性生殖细胞分化过程中起到重要的调节作用。
RNA-Seq;鸡胚胎干细胞;原始生殖细胞;精原干细胞;雄性生殖细胞;一氧化氮合成酶2;抑制剂;分化
0 引言
【研究意义】生殖细胞在生命循环和传承中有着不可替代的作用,因为其可以将遗传信息传递至下一代,这一特性使其迅速成为生命科学研究的重要课题之一。多年来,科研工作者一直致力于将ESC体外诱导分化为生殖细胞,以解决不孕不育的问题,并通过生殖细胞的研究来探索细胞增殖、分化和发育的调控机制[1]。【前人研究进展】细胞增殖、分化和发育与细胞内代谢水平息息相关,蛋白质代谢在细胞分化过程中具有重要的作用。韩明权[2]等在研究药物对肿瘤细胞的抑制效应时发现,肿瘤细胞的蛋白质代谢失常,S+G2/M 期细胞比例降低,从而影响了肿瘤细胞的分化和增殖情况;Fei等[3-6]分别发现Smad 蛋白家族是细胞内重要的信号转导蛋白,在多种细胞分化和转分化过程具有重要的调控作用。TAN等[7]在研究精氨酸对新生仔猪小肠细胞的影响时发现,外源添加精氨酸能够调节新生仔猪体内精氨酸代谢,从而影响小肠细胞的增殖、分化和功能。这些研究都充分说明蛋白质代谢在细胞的增殖、分化和发育过程中扮演非常重要的作用。【本研究切入点】因此,本研究基于前期RNA-Seq深度测序的基础上,通过WEGO分析(http://wego.genomics.org.cn/cgi-bin/wego/index.pl)和KEGG(http://www.funnet.ws/)通路显著富集分析筛选与蛋白质代谢相关的通路和关键性差异基因,并对关键基因进行体内和体外抑制,旨在初步探究蛋白质代谢及其关键基因对雄性生殖细胞分化的调节机制,为建立固定、规范、有效的体外细胞诱导体系打下良好的基础。【拟解决的关键问题】本试验初步探究蛋白质代谢通路对雄性生殖细胞分化的影响,还需要通过 RNA干扰、基因敲除或敲入、免疫共沉淀等一系列的方法进一步探究其具体的作用机制。
1 材料与方法
1.1 实验材料和试剂
试验于2012年8月24日分离细胞起至2014年12月20日完成稿件,于扬州大学推广楼320课题组完成。试验受精蛋来自中国农业科学院家禽研究所实验禽场如皋黄鸡群,共使用18 340枚,分3个重复组,在37.5°C,相对湿度为60%的条件下孵化。孵化分期采用文献[12]和[13]建立的方法。
DMEM培养液(Gibco)、小牛血清(Gibco)、丝裂霉素-C(Roche)、β-琉基乙醇(BBI)、鸡血清、L-谷氨酰胺、丙酮酸钠、胰蛋白酶、胶原酶I、碱性成纤维生长因子(bFGF)、人胰岛素样生长因子(hIGF)、人干细胞因子(hSCF)、小鼠白血病抑制因子(mUF)均源自Sigma公司、NOS2抑制剂S-Methylisothiourea sulfate(Santa Cruz)、维甲酸(RA)(Sigma)。
1.2 试验方法
1.2.1 鸡ESCs的分离、培养、传代及分选 ESCs的分离、培养及传代方法参见文献[13, 14]的方法。采用PCR方法对其进行雌雄鉴定,以排除雌性样本的干扰。
采用流式细胞分选技术,对运用抗体标记的ESC、PGC和SSC进行分选,确保获得高纯度的细胞。其中选择SSEA-1、SOX2干细胞表面特异抗原及分子标记物标记ESC,酪氨酸激酶受体C-kit及SSEA-1标记PGC,精原干细胞重要表面标志 integrin α6及integrinβ1双重标记SSC。
1.2.2 RNA-Seq 参照Illumina公司mRNA-Seq步骤进行 RNA建库,在上海欧易生物医学科技有限公司公司使用Illumina公司HisSeq2000进行测序,上样量为50ng。测序结果同数据库进行比对注释后,进行后续试验分析。RNA标准为RIN≥7,28S/18S>0.7。
1.2.3 QRT-PCR检测 利用RNeasy kit试剂盒提取细胞总 RNA,并反转录成 cDNA,以此为模版进行RT-qPCR。按照荧光定量PCR试剂盒说明,使用SYBR荧光试剂和7900 System荧光定量仪器进行Real-time PCR试验,最后在Microsoft Excel软件内用2-ΔΔCt相对定量法分析试验数据。引物具体信息见表1。 Table 1 Primers of related genes in RT-qPCR

表1 RT-QPCR相关基因的引物
1.2.4 NOS2体内、外抑制试验
1.2.4.1 NOS2体内抑制试验 取刚受精的鸡胚,注射方法参考文献[15],分组情况为:①空白组:不经任何处理,直接孵化;②对照组:中端注射100 μL ddH2O;③ 抑制组:中端注射100 μL 1μmol·L-1的NOS2抑制剂。分别取各组第0天的鸡胚、第5.5天的生殖脊、第18天的睾丸组织,提取RNA,反转录为cDNA,RT-qPCR检测NOS2、C-kit、Cvh、Stra8、Dazl、integrinα6和integrin β1等基因的mRNA表达情况。
1.2.4.2 NOS2体外抑制试验 取培养至 2代鸡ESCs,分组情况为:①对照组:添加普通培养基;②RA诱导组:采用RA诱导;③抑制剂组:对照组的基础上添加1μmol·L-1的:④RA+抑制剂组:在RA诱导的基础上添加工作液浓度1μmol·L-1的抑制剂。观察各组第2、4、6、8和10天的细胞的形态变化,并分别取各组第2、4、6、8和10天的细胞,提取RNA,反转录为cDNA,RT-qPCR检测NOS2、C-kit、Cvh、Stra8、Dazl、integrin α6和integrin β1等基因的mRNA表达情况。
1.2.5 数据处理 所有数据以均数±标准误(X± SEM)表示,应用GraphPad Prism5 统计学软件,采用两样本均数间t检验的方法,P<0.05表示差异显著;P<0.01表示差异极显著。
2 结果
2.1 RNA-Seq分析
在ESC vs PGC和PGC vs SSC两组中分别存在7 967和6 496个差异基因,通过VENNY(http:// bioinfogp. cnb.csic.es/tools/venny/)分析发现,有3 580个差异基因同参与了雄性生殖细胞分化的整个过程,并通过DAVID(http://david.abcc.ncifcrf.gov/)数据库对其进行GO功能注释分类,发现有697个差异基因参与到生物代谢过程。
针对这697个差异基因,再通过KEGG富集通路分析发现,参与雄性生殖细胞整个分化过程的代谢差异基因主要富集于精氨酸-脯氨酸代谢通路、酪氨酸代谢通路和色氨酸代谢通路。3条代谢通路中,在雄性生殖细胞分化的全过程中均差异显著的基因分别有 5个(LOC777369、LOC776984、P4Hα3、P5CSL和NOS2)、2个(HGD和FAH)和1个(IDO)(表2)。

表2 雄性生殖细胞分化过程相关调控差异基因Table 2 Related regulatory genes in differentiation process of male germ cells
2.2 RT-qPCR验证通路基因
针对部分关键基因进行 RT-qPCR验证,以其在ESC中表达量为对照,检测出其在PGC和SSC中的相对表达量(图1)。
根据RT-qPCR结果显示,在ESC分化为SSC的整个阶段中,LOC776984、P4Hα3、P5CSL、HGD、FAH和IDO均先上调后下调,LOC777369和NOS2为先下调后上调。这些基因表达趋势与其在RNA-Seq中的表达趋势基本一致。

图1 相关差异基因相对表达量Fig. 1 Relative expression of related differentiation genes
2.3 NOS2体内、外抑制
2.3.1 NOS2体内抑制 体内抑制后,针对NOS2及C-kit、Cvh、Stra8、Dazl、integrin α6和integrin β1等生殖标记基因进行RT-qPCR检测,以其在第0天鸡胚的表达量为对照,检测其第5.5天生殖脊、第18天睾丸组织的mRNA表达量,结果显示,NOS2及C-kit、 Cvh、Stra8、Dazl、integrin α6和integrin β1等生殖标记基因在空白组和对照组之间无显著性的差异,而在抑制剂组中,NOS2及C-kit、Cvh、Stra8、Dazl、integrin α6和integrin β1等生殖标记基因的mRNA表达量均出现了不同程度的降低,说明NOS2在被抑制后,ESCs向雄性生殖细胞分化过程也受到了抑制(图2)。
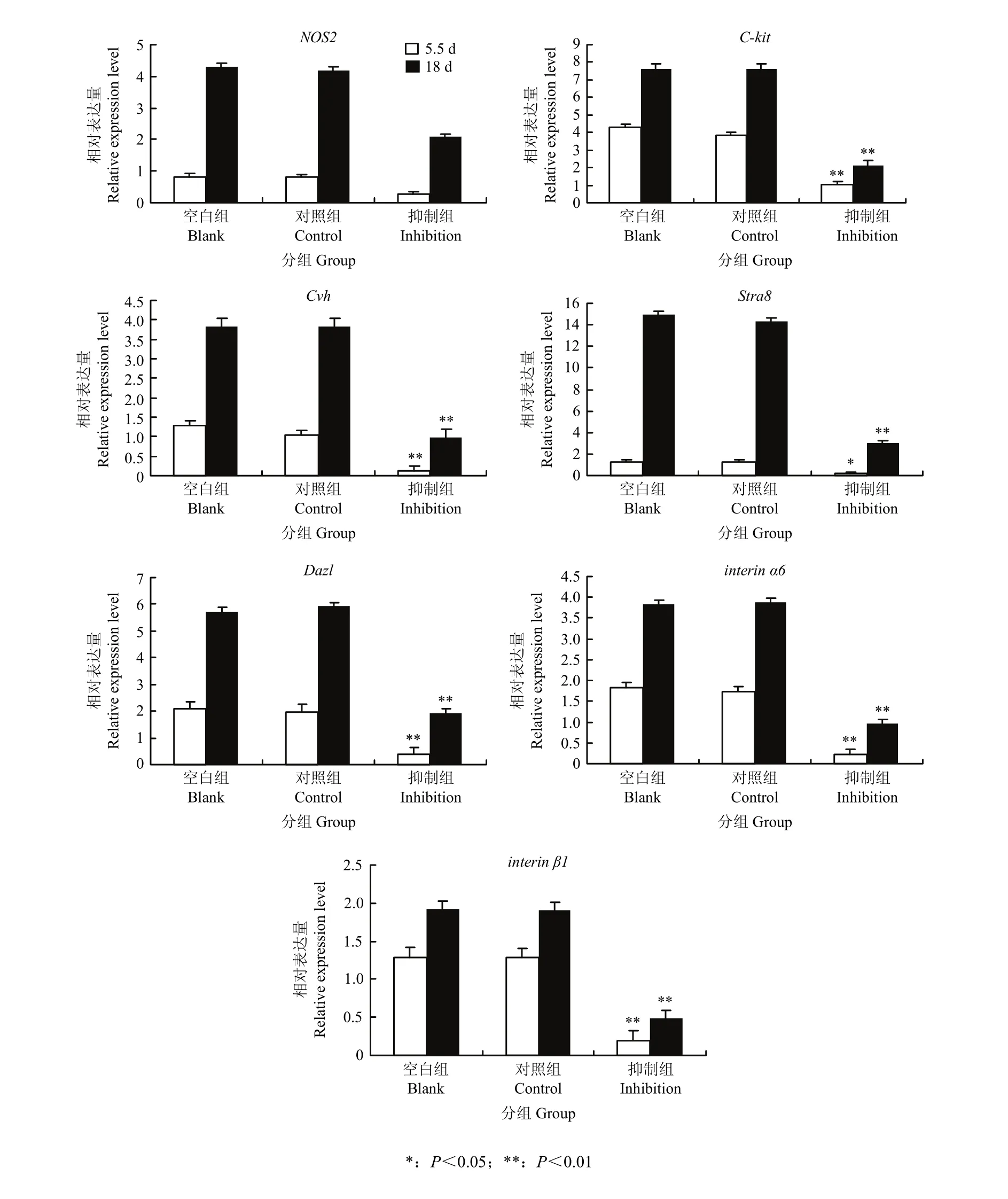
图2 体内抑制中NOS2及相关生殖标记基因相对表达变化Fig. 2 Relative expression of NOS2 and related reproductive marker genes in inhibitory experitment in vivo
2.3.2 NOS2体外抑制 取传至2代的ESCs进行不同分组处理,每隔2 d进行观察细胞形态变化发现,对照组中,ESCs在2、4、6、8和10 d内细胞不断增殖,但是未出现类胚体;RA诱导组中,2 d出现小的类胚体,4 d类胚体增大,且数量增多,6 d类胚体边缘开始出现破裂,8 d类胚体解体,10 d出现类精原样细胞;抑制剂组中,ESCs在2、4、6、8和10d内无类胚体出现,且相较于对照组细胞增殖缓慢;RA+抑制剂组中,2和4 d内无类胚体出现,细胞增殖缓慢,6d出现小的类胚体,8d类胚体数量少量增多,且体积稍显增大,10d类胚体开始裂解。说明NOS2在被抑制后,ESCs向雄性生殖细胞分化过程受到阻碍(图3)。

图3 不同处理组不同天数的ESCs形态变化Fig. 3 Morphological observation of chicken ESCs on different days among different groups (400×)
每隔2 d分别取以上不同分组的细胞,提取细胞RNA,对NOS2及C-kit、Cvh、Stra8、Dazl、integrin α6和integrin β1等生殖标记基因进行RT-qPCR检测,结果发现,经过抑制剂的抑制后,NOS2及C-kit、Cvh、Stra8、Dazl、integrin α6和integrin β1等生殖标记基因的表达量在 RA诱导组、抑制剂组和 RA+抑制剂组中相对于对照组均呈显著性或极显著性的下调趋势,这说明 NOS2在受到抑制剂抑制后,ESCs向雄性生殖细胞的分化受到抑制(图4)。
3 讨论

图4 体外抑制中NOS2及相关生殖标记基因相对表达变化Fig. 4 Relative expression of NOS2 and related reproductive marker genes in inhibitory experitment in vitro
鸡雄性生殖细胞的增殖、分化和发育与其各类代谢过程密切相关,目前已有报道称 BMP4/Smad蛋白通路[16-17]、RA代谢[18]等通路对雄性生殖细胞的增殖、分化、迁移、凋亡具有重要的作用。在这些代谢过程中,蛋白质是生物细胞赖以生存的各种代谢和调控途径的主要执行者,因此蛋白质的代谢对于雄性生殖细胞的增殖、分化和发育至关重要。一氧化氮合成酶2(nitric oxide synthase 2,NOS2)与PGCs的发育和正常生殖细胞的分化密切相关。作为Nanos的同系物,NOS2是在秀丽隐杆线虫(C. Elegans)中首次发现,其结构与Nanos结构类似,都包含一个锌指结构域[19-20]。早期的研究发现,NOS2能够在生物体内催化生成NO,低浓度的NO对生殖细胞的的功能具有保护作用,而高浓度的NO反而具有损坏效应,这说明NOS2通过催化生成NO,在生殖细胞的各种活动中可能起重要的作用[21-24]。YUMIKO[25]等发现NOS2能够启动小鼠雄性生殖细胞的分化,而抑制雌性生殖细胞分化;该研究还发现NOS2表达于SSCs中,是精子形成时维持干细胞数量的一个内因;BARRIOS等[26]发现敲除NOS2的雄性小鼠表现不育,原因是由于在胚胎期缺乏生殖细胞,说明NOS2具有维持生殖细胞数量的作用,然而RA可以下调NOS2的表达,进而促进有丝分裂;CHILDS[27]等发现 Cyp26b1基因敲除,小鼠胚胎生殖细胞中Stra8的表达急剧上调,从而诱导雄性生殖细胞启动减数分裂,这与TSUDA[28]等在NOS2基因敲除小鼠模型中发生的现象一致;NAKAMURA等[29]在以成年青鳉鱼为动物模型,获得NOS2-eGFP标记的卵源干细胞后发现卵源干细胞能够连续产生具备受精能力的干细胞,说明NOS2对生殖细胞的增殖、分化起到重要作用;KÖPRUNNER等[30]以斑马鱼为试验对象,发现缺少NOS1和NOS2时导致PGCs无法维持其功能,并大量死亡;KÖPRUNNER还发现NOS2在PGCs与性腺有效结合过程中也起到重要作用。这些都说明NOS2在生殖细胞自我增殖及其向SSCs分化中的重要作用。
本研究探索出在ESCs向SSCs分化的整个过程中,蛋白质代谢的主要信号通路是精氨酸-脯氨酸代谢通路、酪氨酸代谢通路以及色氨酸代谢通路,其中有大量的研究表明精氨酸-脯氨酸代谢通路上的关键基因NOS2参与了生殖细胞的各种活动,在此理论基础上,以NOS2特异性抑制剂分别在体内和体外对NOS2进行抑制,结果表明NOS2在被抑制后,引起C-kit、Cvh、Stra8、Dazl、integrin α6和integrin β1等生殖标记基因表达量的显著下降,使得ESCs向SSCs方向的分化明显受到抑制。然而,NOS2对ESCs向生殖方向分化的具体调节机制、以及精氨酸-脯氨酸代谢通路与其他的信号通路是否有互作等问题还需要进一步的探究。
另外,亦有多项研究[31-34]表明,FAH和IDO等基因也参与细胞的增殖、分化及其发育,但尚未有其关于影响雄性生殖细胞生成、分化的报道,因此,关于这些基因的具体调节机理还需进一步探索。
4 结论
本研究基于前期探究出NOS2及其所在的精氨酸-脯氨酸代谢通路在鸡雄性生殖细胞分化过程中起到重要调节作用的基础上,以抑制剂对NOS2进行体内、外的抑制,发现NOS2在被抑制后,ESCs向雄性生殖细胞分化的过程受到抑制。说明了精氨酸-脯氨酸代谢通路及关键基因NOS2对ESCs向雄性生殖细胞分化过程中起到重要的调节作用。
[1] 孙敏, 施青青, 李碧春. 胚胎干细胞诱导分化为雄性生殖细胞的研究进展. 生命科学,2012, 24(1):37-42.
SUN M, SHI Q Q, LI B C. Research progress on differentiation of embryonic stem cells into male germ cells. Chinese Bulletin of Life Sciences, 2012, 24(1): 37-42. (in Chinese)
[2] 韩明权, 刘嘉湘, 高虹, 陈善香, 朱晏伟, 许玲. 益肺抗瘤饮对实验性肺癌细胞周期及核酸和蛋白质合成的影响. 中西医结合学报,2003, 1(3):205-208.
HAN M Q, LIU J X, GAO H, CHEN S X, ZHU Y W, XU L. Effects of Yifei Kangliu Qral Liquid on cell cycle and protein-nucleic acid synthesis of experimental lung cancer. Journal China of Integrative Medicine, 2003, 1(3):205-208. (in Chinese)
[3] FEI T, XIA K, LI Z W, ZHOU B, ZHU S S, CHEN H, ZHANG J P, CHEN Z, XIAO H S, HAN J D J, CHEN Y G. Genome-wide mapping of SMAD target genes reveals the role of BMP signaling in embryonic stemcell fate determination. Genome Research, 2010, 20( 1) : 36-44.
[4] CHEN W, JIA W W, WANG K, ZHOU Q, LENG Y, DUAN T, KANG J H. Retinoic acid regulates germ cell differentiation in mouse embryonic stem cells through a Smad-dependent pathway. Biochemical and Biophysical Research Communications, 2012, 418(3): 571-577.
[5] NISHIMURA Y, KURISAKI A, NAKANISHI M, OHNUMA K, NINOMIYA N, KOMAZAKI S, ISHIURA S, ASASHIMA M. Inhibitory Smad proteins promote the differentiation of mouse embryonic stem cells into ependymal-like ciliated cells. Biochemical and Biophysical Research Communications, 2010, 401(1): 1-6.
[6] HELLINGMAN C A, DAVIDSON E N, KOEVOET W, VITTERS E L, VAN DEN BERG W B, VAN OSCH G J V M, VAN DER KRAAN P M. Smad signaling determines chondrogenic differentiation of bonemarrow-derived mesenchymal stem cells: inhibition of Smad1/5/8Pprevents terminal differentiation and calcification. Tissue Engineering: Part A, 2011, 17(7-8): 1157-1167.
[7] TAN B, YIN Y L, KONG X F, LI P, LI X L, GAO H J, LI X G, HUANG R L, WU G Y. L-Arginine stimulates proliferation and prevents endotoxin-induced death of intestinal cells. Amino Acids, 2009, 38(4): 1227-1235.
[8] SULTAN M, SCHULZ M H, RICHARD H, MAGEN A, KLINGENHOFF A, SCHERF M, SEIFERT M, BORODINA T, SOLDATOV A, PARKHOMCHUK D, SCHMIDT D, O'KEEFFE S, HAAS S, VINGRON M, LEHRACH H, YASPO M L. A global view of gene activity and alternative splicing by deep sequencing of the human transcriptome. Science, 2008, 321(5891): 956-960.
[9] TANG F C, BARBACIORU C, BAO S Q, LEE C, NORDMAN E, WANG X H, LAO K Q, SURANI M A. Tracing the derivation of embryonic stem cells from the inner cell mass by single-cell RNA-Seq analysis. Cell Stem Cell, 2010, 5(6): 397-398.
[10] TRAPNELL C, WILLIAMS B A, PERTEA G, MORTAZAVI A, KWAN G, VAN BAREN M J, SAIZBERG S L,WOLD B J, PACHTER L. Transcript assembly and quantification by RNA-Seq reveals unannotated transcripts and isoform switching during cell differentiation. Nature Biotechnology, 2010, 28: 511-515.
[11] TANG F C, BARBACIORU C, NORDMAN E, LI B, XU N L, BASHKIROV V I, LAO K Q, SURANI M A. RNA-Seq analysis to capture the transcriptome landscape of a single cell. Nature Protocols, 2010, 5: 516-535.
[12] HAMBURGER V, HAMILTON H L. A series of normal stages in the development of the chicken. Morphology, 1951, 88(1): 49-92.
[13] 李碧春, 陈国宏, 赵东伟, 王克华, 钱菊芬. 鸡胚 PGCs迁移与性腺发育关系的研究. 扬州大学学报, 2002, 23(1): 18-26.
LI B C, CHEN G H, ZHAO D W, WANG K H, QIAN J F. Relationship between PGCs migration and gonad development in the earlu chicken embryo. Journal of Yangzhou University (Agricultural and Life Science Edition), 2002, 23(1): 18-26. (in Chinese)
[14] 孙敏, 施青青, 傅德智, 阴彦辉, 张亚妮, 李碧春. 鸡胚 ESCs 和
SSCs 特定基因表达差异的研究. 生物技术, 2011, 21(3): 16-19. SUN M, SHI Q Q, FU D Z, YIN Y H, ZHANG Y N, LI B C. Study of the expression of gene cell differentiation associated gene on chicken ESC and SSCs. Biotechnology, 2011, 21(3): 16-19. (in Chinese)
[15] 汪怡临, 靳锴, 蒋舒颖, 赵瑞丰, 左其生, 李东, 王颖洁, 张蕾, 张亚妮, 李碧春. 不同注射部位和剂量对鸡胚发育的影响. 中国家禽, 2014, 36(17): 7-10.
WANG Y L, JIN K, JIANG S Y, ZHAO R F, ZUO Q S, LI D, WANG Y J, ZHANG L, ZHANG Y N, LI B C. Effects of different injection location and dose on chicken embryos. China Poultry, 2014, 36(17): 7-10. (in Chinese)
[16] 陈兵, 易斌, 鲁开智. Smad 蛋白家族调控细胞分化的研究进展.医学研究生学报, 2013, 26 (5): 544-547.
CHEN B, YI B, LU K Z. Advances in researches on Smad proteins in cell differentiation. Journal of Medical Postgraduates, 2013, 26 (5): 544-547. (in Chinese)
[17] ANDREA V C, DIANA J L. Wnt and Bmp fit germ cells to a T. Developmental Cell. 2013, 27(5): 485-487.
[18] SUGIMOTO R, NABESHIMAB Y, YOSHIDA S. Retinoic acid metabolism links the periodical differentiation of germ cells with the cycle of Sertoli cells in mouse seminiferous epithelium. Mechanisms of Development, 2012, 128(11-12): 610-624.
[19] SUBRAMANIAM K, SEYDOUX G. nos-1 and nos-2, two genes related to Drosophilananos, regulate primordial germ cell development and survival in Caenorhabditis elegans. Development, 1999, 126: 4861-4871.
[20] GALLO C M, WANG J T, MOTEGI F, SEYDOUX G. Cytoplasmic partitioning of P granule components is not required to specify the germline in C. elegans. Science, 2010, 330(6011): 1685-1689.
[21] ZINI A, DE LAMIRANDE E. Low levels of nitric oxide promote human sperm capacitation in vitro. Journal of Andrology, 1995, 16 (5): 424-431.
[22] ZINI A, O’BRYAN M K, SCHLEGEL P N. Nitric oxide synthase activity in human seminal plasma. Urology, 2001, 58 (1) :85-89.
[23] HERRERO M B, DE LAMIRANDE E, GAGNON C. Nitric oxide regulates human sperm capacitation and protein-tyrosine phosphorylation in vitro. Biology of Reproduction, 1999, 61(3): 575-581.
[24] MITROPOULOS D, DELICONSTANTINOS G. Nitric oxide synthase and xanthine oxidase activities in the spermatic vein of patients with varicocele: apotential role for nitric oxide and peroxynitrite in sperm dysfunction. The Journal of Urology, 1996, 156(6): 1952-1958.
[25] YUMIKO S. Function of Nanos2 in the male germ cell lineage in mice. Cellular and Molecular Life Sciences, 2010, 67(22): 3815-3822.
[26] BARRIOS F, FILIPPONI D, PELLEGRINI M, PARONETTO M P, SIEN S D, GEREMIA R, ROSSI P, DE FELICI M, JANNINI E A, DOLCI S. Opposing effects of retinoic acid and FGF9 on Nanos2 expression and meiotic entry of mouse germ cells. Journal of Cell Science, 2010, 123: 871-880.
[27] CHILDS A J, COWAN G, KINNELL H L, ANDERSON R A, SAUNDERS P T K. Retinoic acid signalling and the control of meiotic entry in the human fetal gonad. PLoS One, 2011, 6(6): e20249.
[28] TSUDA M, SASAOKA Y, KISO M, ABE K, HARAGUCHI S, KOBAYASHI S, SAGA Y. Conserved role of nanos proteins in germ cell development. Science, 2003, 301(5637): 1239-1241.
[29] NAKAMURA S, KOBAYASHI K, NISHIMURA T, HIGASHIJIMA S, TANAKA M. Identification of germline stem cells in the ovary of the teleost medaka. Science, 2010, 328: 1561-1563.
[30] KÖPRUNNER M, THISSE C, THISSE B, RAZ E. A zebrafish nanos-related gene is essential for the development of primordial germ cells. Genes & Development, 2001, 15: 2877-2885.
[31] 刘秀红, 李宁. 肝病与肝癌的新疗法:干细胞研究的最新进展. 北京医学, 2013, 35(9): 813-814.
LIU X H, LI N. New therapy of Liver disease and liver cancer: the latest progress in the study of stem cells. Beijing Medical Journal, 2013, 35(9): 813-814. (in Chinese)
[32] JANG Y Y, Collector M I, Baylin S B, Deihl A M, Sharkis S J. Hematopoietic stem cells convert into liver cells within days without fusion. Nature Cell Biology, 2004; 6: 532-539.
[33] TAKIKAWA O, KUROIWA T, YAMAZAKI F, KIDO R. Mechanism of interferon-gamma action. Characterization of indoleamine 2,3-dioxygenase in cultured human cells induced by interferon-gamma and evaluation of the enzyme-mediated tryptophan degradation in its anticellular activity. Journal of Biological Chemistry, 1988, 263(4): 2041-2048.
[34] Viola A, Bronte V. Metabolic mechanisms of cancer inducedinhibition of immune responses. Seminars in Cancer Biology, 2007, 17(4): 309-316.
(责任编辑 林鉴非)
Regulatory Study of Protein Metabolism During the Differentiation Process of Chicken Male Germ Cells
LI Dong, TANG Bei-bei, WANG Ying-jie, JI Yan-qin, WANG Fei, LU Zhen-yu, WANG Man, ZHANG Ya-ni, LI Bi-chun
(Key Laboratory of Animal Breeding Reproduction and Molecular Design for Jiangsu Province, College of Animal Science and Technology, Yangzhou University, Yangzhou 225009, Jiangsu)
【Objective】 The aim of this study was to explore the regulatory mechanism of protein metabolism during the differentiation process of chicken male germ cells and provide a basis for improving the induction system of chicken embryonic stem cells (ESCs) differentiation to male germ cells in vitro. 【Method】RNA sequencing was performed using FACS-sorted cells from ESCs, PGCs(primordial germ cells) and SSCs(spermatogonial stem cells), and enrichment analysis, WEGO (Web Gene ontology) and KEGG (Kyoto Encyclopedia of Genes and Genomes), were carried out to find out the relevant pathways and the key genes, the expression level of which was analyzed by qRT-PCR. Moreover, NOS2 both in vitro and in vivo with NOS2 inhibitor was inhibited, and the morphologic changes of ESCs were observed and the mRNA expressions of NOS2 and other germ genes, C-kit, Cvh, Stra8, Dazl, integrin α6 and integrin β1 were detected in different groups and in different days with RT-qPCR. 【Result】 Final results showed that 697 differentially expressed genes were involved in biological metabolism and significantly enriched in arginine-proline metabolic pathway, tyrosine metabolic pathway and tryptophan metabolic pathway and screened some key genes, like NOS2, FAH and IDO. It was found that the expression trends of NOS2, FAH and IDO were the same as that of RNA-Seq. In inhibitory experiment in vivo, the mRNA expression of NOS2, C-kit, Cvh, Stra8, Dazl, integrin α6 and integrin β1 between blank group and control group showed no significant difference. However, in inhibited group, NOS2, C-kit, Cvh, Stra8, Dazl, integrin α6 and integrin β1 expressions were down-regulated inordinately. Moreover, in inhibitory experiment in vitro, ESCs always proliferated on the 2, 4, 6, 8 and 10d, but disappeared the embryonic bodies in the control group. In induced group, small embryonic bodies appeared on the 2d and became bigger and increased on the 4d. Embryonic bodies started to burst in edges on the 6d, break up on the 8d and appeared spermatogonia-like cells. In inhibited group, no embryonic body appeared in the whole process and ESCs proliferated more slow than the control group. In induced-inhibited group, no embryonic body appeared on the 2d and 4d and ESCs proliferated slowly. Small embryonic bodies appeared on the 6d and the number and volume increased slightly on the 8d. On the 10d, the embryonic bodies started to break up. In vitro, NOS2, C-kit, Cvh, Stra8, Dazl, integrin α6 and integrin β1 expressions in induced group, inhibited group and induced-inhibited group were significantly down-regulated compared with the control group. 【Conclusion】In this study, based on the screening of arginine-proline metabolic pathway and NOS2 with RNA-Seq and Bioinformatics, it was found that the process of ESCs differentiation to male germ cell was inhibited after the inhibition of NOS2,which suggested that arginine-proline metabolic pathway and NOS2 has an important regulatory effect on differentiation of ESCs to male germ cells.
RNA-Seq; ESCs; PGCs; SSCs; male germ cells; NOS2; inhibitor; differentiation
2015-08-03;接受日期:2016-11-02
国家自然科学基金(31272429)、高等学校博士学科点专项科研基金资助课题(20103250110006)、江苏省“六大人才高峰”、江苏省优势学科
联系方式:李东,E-mail:lidongyzu@hotmail.com。通信作者李碧春,E-mail:yubcli@yzu.edu.cn。通信作者张亚妮,E-mail:ynzhang@yzu.edu.cn
