干旱胁迫下3个杨树品种叶片膜透性及部分渗透调节物质的变化
2017-01-12杨淑红任媛媛朱延林
杨淑红,朱 镝,任媛媛,朱延林
(1河南省林业科学研究院,郑州 450008;2福建农林大学林学院,福州 350002)
干旱胁迫下3个杨树品种叶片膜透性及部分渗透调节物质的变化
杨淑红1,朱 镝2,任媛媛1,朱延林1
(1河南省林业科学研究院,郑州 450008;2福建农林大学林学院,福州 350002)
在土壤持续干旱胁迫下,研究了杨树品种‘2025’及其2个芽变品种‘全红杨’、‘中红杨’当年生嫁接苗叶片的质膜相对透性及渗透调节物质可溶性糖、脯氨酸和可溶性蛋白质含量的变化及关系,并进行了抗旱性评价。结果表明:随干旱胁迫的持续,三者叶片质膜相对透性明显增大,‘全红杨’始终高于‘中红杨’和‘2025’。三者叶片可溶性蛋白质和游离脯氨酸总量有减少的趋势,但可溶性糖含量均能保持在较高的水平,与质膜相对透性成显著正相关(1>|r|≥0.7),这有利于植株适应干旱胁迫和抗旱能力的获得;‘2025’叶片脯氨酸的变化与质膜相对透性呈显著正相关(1>|r|≥0.7),膜系统相对稳定。3个品种各生理指标对照组和处理组均有极显著差异(P<0.01),经隶属函数法分析得出:抗旱能力为‘2025’>‘中红杨’>‘全红杨’,但差异不显著(P>0.05);通过灰色关联法分析各指标对干旱胁迫的敏感性可知:游离脯氨酸是反映3个杨树品种抗旱能力的主要指标。
杨树品种;干旱胁迫;渗透调节;抗旱能力
干旱是我国目前面临的重要生态问题,水是树木生长和形成的先决条件,对树种分布、生长、生态生理过程等有重要的影响,因此进行树木新品种干旱胁迫研究尤为必要[1]。树木的耐干旱性是长期自然选择的结果,主要由体内的遗传基因控制,具有遗传的相对稳定性和潜在反应性,树木只有处于干旱环境一段时间,耐水胁迫性才能充分显示出来。大量证据表明,细胞膜是植物体内部与外部物质交换的通道,为植物细胞维持正常的生理代谢活动提供相对稳定的内环境。干旱胁迫首先造成细胞膜系统状态发生改变而触发一系列水分伤害反应,树木叶片质膜相对透性变化可反映出细胞遭受逆境伤害的程度,质膜相对透性增强,细胞及细胞器内稳定的生理生化环境遭到破坏,导致细胞生理功能丧失,大量内溶物外渗,最终细胞衰老死亡[2-4],严重影响树木的正常生长发育。干旱胁迫也可使蛋白质和核酸变性失活,严重时会抑制蛋白质的合成并诱导其降解以及纤维分子间的桥键松驰,从而破坏生物膜的结构与功能。可溶性蛋白质、可溶性糖和游离脯氨酸均是植物体内重要的有机渗透调节物质,其含量有助于细胞和组织的保水,起到抗脱水作用,是判断植物细胞遭受水分胁迫程度的常用生理指标[5-7]。
‘中红杨’(Populus×euramericana‘Zhonghong’)为美洲黑杨‘2025’(P.deltoids cv.‘Lux’(I-69/55)× P.deltoids cv.‘Shan Hai Guan’)的芽变彩叶品种,随后在‘中红杨’基础上选育出芽变新品种‘全红杨’(Populus×euramericana‘Quanhong’)。‘全红杨’姿态秀丽,枝叶色泽较‘中红杨’更加亮丽持久,观赏效果甚佳。‘中红杨’和‘全红杨’均已通过新品种认定,并分别于2006年和2011年获得国家新品种保护权,均为难得的速生丰产性高品质彩叶园林观赏树木。本试验以‘全红杨’、‘中红杨’和‘2025’当年生嫁接苗为试材,盆栽控水使植株处于土壤持续干旱胁迫下,研究叶片膜透性和渗透调节物质可溶性糖、脯氨酸及可溶性蛋白质含量的变化及相互关系,比较三者抗旱性生理差异,并采用隶属函数法和灰色关联分析法对3个杨树品种及生理指标进行抗旱能力综合评价。
1 材料与方法
1.1 试验材料
试验地点为河南省林业科学研究院院内,2011年2月下旬将长势健康一致的‘2025’一年生实生苗移植于普通塑料圆形花盆内(上直径×下直径×高:350 mm×250 mm×350 mm),每盆1株,平茬至20 cm高,盆土为50%普通园土+50%腐质土,装土量一致,正常管理。3月末分别嫁接‘全红杨’、‘中红杨’和‘2025’各50盆。4个月后待嫁接苗稳定成活,长势旺盛时进行控水试验。
1.2 试验处理
2011年7月28日,各品种挑选长势良好均衡的30盆嫁接苗,搬至排水条件良好的硬化地面上,随即浇水至饱和。试验完全随机区组设计,对照(CK)组与土壤干旱胁迫处理(H)组每个品种15盆,按5株/行×3行排列,CK和H组组间距3 m。CK组苗木每隔3—5 d浇1次水至盆土饱和,H组苗木7月29日起停止浇水,使土壤含水量逐渐降低,遇阴雨天采用移动遮雨棚防风遮雨,8月25日解除胁迫浇复活水至饱和,随后正常养护。分别在干旱胁迫0 d(7月28日)、4 d(8月1日)、7 d(8月4日)、14 d(8月11日)、21 d(8月18日)、28 d(8月25日)及复水7 d(9月1日)的8:00采集各组各品种植株枝条顶端往下第3—5片叶,设3个重复,采样后的植株不再重复采样。样叶袋封存置于冰桶中并立即带回实验室进行理化指标测定。
1.3 测定指标及方法
质膜透性测定采用相对电导率法[8];可溶性糖测定采用蒽酮比色法;游离脯氨酸测定采用磺基水杨酸比色法;可溶性蛋白质测定采用考马斯亮蓝法[9]。
1.4 统计分析
采用Excel 2007和SPSS 17.0软件进行数据处理及统计分析。
采用隶属函数法对3个杨树品种抗旱能力进行综合评定[3,10]。为避免不同物种间对照值的差异对试验结果造成影响,对所测得的各项生理指标值进行转换,求得各生理指标在不同时间的变化系数。公式:Ii=处理指标i/对照指标i0。
指标与抗旱性正相关隶属函数公式:R(Xij)=(Xij-Xmin)/(Xmax-Xmin);指标与抗旱性负相关隶属函数公式:R(Xij)=1-(Xij-Xmin)/(Xmax-Xmin)。式中Xij为i品种j指标抗旱系数,Xmax、Xmin为j指标抗旱系数的最大值和最小值,R(Xij)为i品种j指标的抗旱隶属值。求取各抗旱指标隶属函数值的平均值,综合评定值越大说明抗旱能力越强。
采用灰色关联分析法[11]将3个杨树品种的各项生理指标及抗旱隶属函数均值设为数据集合,建立灰色系统。设抗旱隶属函数平均值作为参考数据列(母序列)X0;以各项生理指标的平均抗旱系数作为比较列(子序列)X1、X2、X3、X4,分别表示质膜相对透性、可溶性糖、脯氨酸和可溶性蛋白质。利用计算机DPS处理系统执行灰色关联分析,分辨系数取常规值0.5[3,10],得出3个杨树品种各指标与抗旱性的关联度与关联序。
2 结果与分析
2.1 土壤持续干旱对叶片质膜相对透性的影响
由图1可知,对照组‘全红杨’、‘中红杨’和‘2025’叶片相对电导率平均值分别为29.69%、30.80%和30.78%,差异不显著(P>0.05)。干旱胁迫期间,三者叶片相对电导率持续升高,胁迫21 d时‘全红杨’较对照组升高102.27%;胁迫28 d时,‘全红杨’、‘中红杨’和‘2025’叶片相对电导率极显著高于各自对照组(P<0.01),分别升高84.96%、85.09%和91.83%,且‘全红杨’显著高于‘中红杨’和‘2025’。复水7 d时,‘全红杨’相对电导率较对照组升高43.51%,极显著高于对照组,‘中红杨’和‘2025’显著高于对照组(P<0.05)。
2.2 土壤持续干旱胁迫对叶片可溶性糖含量的影响
可溶性糖是合成其他有机溶质的碳架保护和能量来源,胁迫条件下可增加细胞原生质浓度,降低细胞渗透势,维持细胞内正常的水分代谢,对细胞膜和原生胶体有稳定作用,有利于提高植株的抗逆性[6]。由图2可知,对照组‘全红杨’叶片可溶性糖含量平均值显著低于‘中红杨’和‘2025’。干旱胁迫期间,三者叶片可溶性糖含量均明显增加,处理初期‘全红杨’叶片可溶性糖含量迅速升高,胁迫21 d时达最大值29.78 mg/g FW,较对照组升高48.95%;胁迫28 d时,‘全红杨’、‘中红杨’和‘2025’可溶性糖含量为28.55 mg/g FW、33.49 mg/g FW和35.28 mg/g FW,分别较各自对照组升高39.24%、45.13%和56.17%。复水7 d时,三者叶片可溶性糖含量仍极显著高于各自对照组47.02%、41.43%和43.40%。相关分析表明:干旱胁迫下,‘全红杨’、‘中红杨’和‘2025’叶片可溶性糖的变化均与质膜相对透性呈显著相关(Y=3.1196+0.8917X-0.0076X2,r=0.793**,df=18,F=24.074**;Y=14.3991+0.3312X,r=0.791**,df=19,F=31.7233**;Y=22.0955+0.0039X2,r=0.806**,df=19,F=37.7383**;P=0.01,1>|r|≥0.7为显著相关;0.7>|r|≥0.4为有相关;0.4>|r|≥0为无相关[12],下同)。
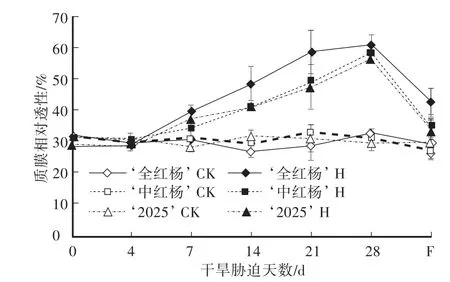
图1 干旱胁迫及复水对叶片质膜相对透性的影响Fig.1 Effects of drought stress and rewatering on the relative permeability of the plasma membrane of leaves

图2 干旱胁迫及复水对叶片可溶性糖含量的影响Fig.2 Effects of drought stress and rewatering on the soluble sugar content in leaves
2.3 土壤持续干旱胁迫对叶片脯氨酸含量的影响
游离脯氨酸作为一种碳水化合物的来源,是酶和细胞结构的保护剂,在含水量很低的细胞内仍能提供足够的自由水,稳定膜系统,参与叶绿素合成,提高植物抗性[7,13]。由图3可知,对照组‘全红杨’叶片游离脯氨酸含量平均值显著低于‘中红杨’和‘2025’。干旱胁迫期间,‘全红杨’、‘中红杨’和‘2025’叶片脯氨酸含量均先升后降,积累峰值分别为31.27μg/g FW(胁迫14 d)、32.71μg/g FW(胁迫21 d)和31.98 μg/g FW(胁迫21 d),较各自对照组升高28.96%、36.24%和26.29%;胁迫28 d时,‘全红杨’、‘中红杨’和‘2025’叶片脯氨酸含量仍极显著高于对照组,分别较各自对照组升高13.62%、17.03%和24.44%,胁迫后期‘全红杨’叶片游离脯氨酸含量极显著低于‘中红杨’和‘2025’。复水7 d,三者叶片脯氨酸含量仍显著高于各自对照组2.57%、4.44%和8.52%。相关分析表明:干旱胁迫下,叶片脯氨酸的变化与质膜相对透性‘全红杨’、‘中红杨’为正相关(r=0.473;r=0.671),‘2025’为显著正相关(Y=18.4872+0.2228X,r=0.748**,df=19,F=24.134**)。
2.4 土壤持续干旱胁迫对叶片可溶性蛋白质含量的影响
植物体内可溶性蛋白质大多是参与各种代谢的酶类,具有较强的亲水胶体性质,其含量可影响细胞的保水力,也是植物对逆境胁迫适应的一种表现。由图4可知,对照组中,‘全红杨’、‘中红杨’和‘2025’叶片可溶性蛋白质含量平均值差异不显著。干旱胁迫期间,三者叶片可溶性蛋白质含量变化各不相同,‘全红杨’叶片可溶性蛋白质含量先升高后降低,胁迫7 d时达到最大值47.17 mg/g FW,较对照组升高15.98%,胁迫28 d时为29.36 mg/g FW,较对照组降低25.12%;‘中红杨’叶片可溶性蛋白质含量呈降-升-降变化曲线,胁迫7 d、21 d和28 d时含量分别为36.08 mg/g FW、47.69 mg/g FW和37.34 mg/g FW,分别较对照组升高-11.69%、13.42%和-11.01%;‘2025’叶片可溶蛋白质含量呈振荡升高后快速下降的趋势,胁迫21 d时达最大值50.01 mg/g FW,较对照组升高19.90%,胁迫28 d时降为40.65 mg/g FW,较对照组降低5.20%。胁迫前期,‘中红杨’叶片可溶性蛋白质含量极显著低于‘全红杨’、‘2025’及对照组;胁迫后期,‘全红杨’叶片可溶性蛋白质含量极显著低于‘中红杨’、‘2025’及对照组。复水7 d时,‘全红杨’叶片可溶性蛋白质含量极显著低于对照组,较对照组降低35.71%,‘中红杨’和‘2025’显著低于各自对照组11.01%和4.04%。相关分析表明:干旱胁迫下,‘全红杨’可溶性蛋白质的变化与质膜相对透性为有相关(r=-0.456),‘中红杨’和‘2025’可溶性蛋白质的变化与质膜相对透性为无相关(r=0.291;r=0.208)。
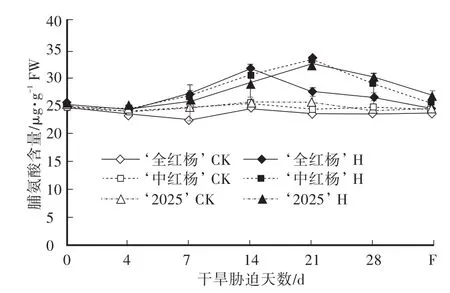
图3 干旱胁迫及复水对叶片脯氨酸含量的影响Fig.3 Effects of drought stress and rewatering on the proline content in leaves
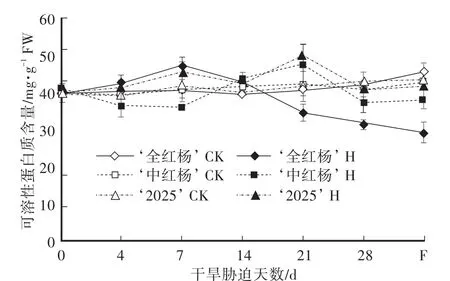
图4 干旱胁迫及复水对叶片可溶性蛋白质含量的影响Fig.4 Effects of drought stress and rewatering on the soluble proline content in leaves
2.5 抗旱性的综合评价
利用隶属函数法,对3个杨树品种在干旱胁迫下质膜相对透性、可溶性糖、游离脯氨酸和可溶性蛋白质所表现的抗旱性状进行综合评价。分析结果表明(表1),抗旱能力为:‘2025’>‘中红杨’>‘全红杨’,差异不显著。

表1 3个杨树品种叶片生理指标的平均隶属函数值及排序Table 1 Average membership function value and ranking of physiological indexes in leaves of 3 poplar varieties
根据灰色系统理论,若某指标与抗旱性的关联度越大,则说明该指标与变化系数的关系越密切,对干旱胁迫的反应越敏感。从表2可以看出,游离脯氨酸与3个杨树品种的变化系数关联度最大为0.746,其次是可溶性蛋白质和可溶性糖,均处于较强关联水平,而质膜相对透性对三者抗旱性的影响较小,此研究结果与黄承玲等[3]、周江等[10]基本相同,而与胡尚连等[14]不同。
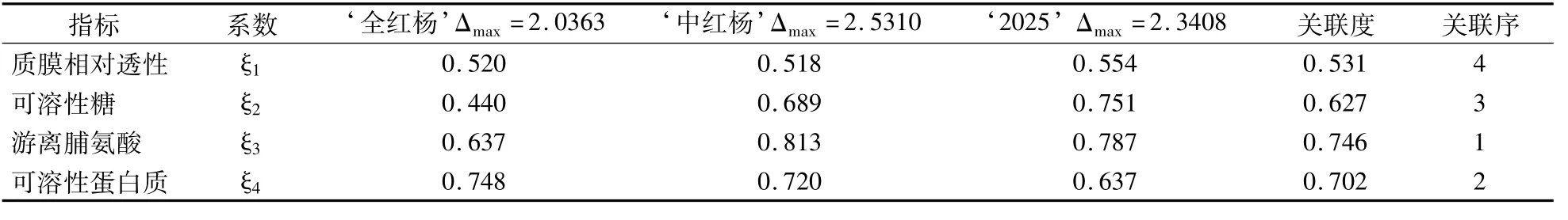
表2 3个杨树品种抗旱性与各生理指标的关联系数、关联度和关联序Table 2 Correlation coefficient,correlation degree and correlation order of drought resistance and physiological indexes of 3 poplar varieties
3 结论与讨论
与许多抗旱性研究结果一样[3,15-16],随着干旱胁迫的持续,3个杨树品种叶片的质膜相对透性均明显增大,‘全红杨’质膜透性始终高于‘中红杨’和‘2025’,细胞内电解质大量外泄,电解质减少,干旱逆境对细胞膜的选择透性破坏严重,使得胁迫后期‘全红杨’细胞膜相对透性升高减缓,这与新西伯利亚银白杨研究结果类似[17]。
蛋白质分解是水分胁迫干扰植物体内氮代谢过程的最突出表现,轻度干旱胁迫可刺激植物体内抗氧化酶合成以增强渗透调节能力,不溶性蛋白质变为可溶性蛋白质,当胁迫超过植物所能忍耐的阈值时,细胞体内肽酶活性提高,大量活性氧会损伤DNA的复制过程,使细胞蛋白质和核酸的合成代谢能力减弱,最终叶肉细胞中可溶性蛋白质合成受阻降解[18],蛋白质降解越慢越少,可减轻产生的氨类物质对植物的毒害,植株抗性越强,蛋白质含量的降低与植物的衰老也密切相关[5,19]。研究中‘全红杨’叶片可溶性蛋白质含量先升高后急速降低,与大白杜鹃[3]、苜蓿[20]等不抗旱树种的研究结果相同;‘中红杨’和‘2025’在胁迫中期可溶性蛋白质含量升高,表现出不同形式的渗透调节能力,与虎榛子、辽东栎[21]、刺槐[5]等的研究相似。3个杨树品种的可溶性蛋白质含量变化不同,可能与各自渗透调节功能有关,适应干旱的主导途径有所差异。可溶性蛋白质含量在不同植物之间变化也较为复杂,有学者发现,白桦实生苗在干旱胁迫下细胞内束缚水的含量并未随可溶性蛋白质含量的降低而减少[22]。另外,植物为了避免胁迫造成伤害,干旱引起蛋白质含量变化的同时,往往也伴随诱导产生一些抗逆蛋白质,称之为“旱胁蛋白”或“逆境蛋白”,这种热稳定性蛋白的生成很可能在抗旱中起着更大的作用[5,23]。
大多研究认为植物脯氨酸积累越多抗逆性能力超强,含量有时会超过正常条件下的十倍或百倍,但也有少数植物在逆境条件下,脯氨酸含量变化不大[24]。其与抗旱性的关系也有不同的观点,张明生等[25]认为游离脯氨酸与品种抗旱性间的相关性不显著,马讳等[26]认为脯氨酸的含量只是反映植株受旱程度,在不抗旱品种中反而积累较多。干旱胁迫下,‘全红杨’、‘中红杨’和‘2025’叶片脯氨酸变化与高山杜鹃[3]、火炬树[27]等的研究结果类似,脯氨酸含量均先升后降,相对于对照组增幅不大,最多增加28.96%、36.24%和26.29%。‘全红杨’叶片中脯氨酸急剧增加出现的时间早于‘中红杨’和‘2025’,曾有人提出以干旱胁迫下游离脯氨酸大量积累的水势阈值作为评价抗旱能力的参考[5]。
干旱胁迫过程中,3个杨树品种叶片可溶性蛋白质和游离脯氨酸总量有减少的趋势,但可溶性糖含量不断的增加或保持在较高的水平,有利于植株应对干旱胁迫和获得抗旱能力[5,15]。一方面是胁迫后期呼吸速率显著上升,多糖的分解速率加快,另外可能三者叶片本身含有较多的多糖,使可溶性糖的产生有充足的原料。胁迫后期‘全红杨’叶片可溶性糖含量稳定在一定高度,复水后变化不明显,说明干旱对其造成了一定的伤害,合成和降解机制受到损伤,其可溶性糖的渗透调节能力相对弱一些。相关分析表明:‘全红杨’、‘中红杨’和‘2025’叶片可溶性糖的变化与质膜相对透性均呈显著正相关,可溶性糖在渗透调节中发挥着重要作用。‘2025’叶片脯氨酸的变化与质膜相对透性呈显著正相关,有利于‘2025’膜系统的稳定,使其具有较强的抗旱能力。‘全红杨’可溶性蛋白质的变化与质膜相对透性呈负相关,干旱胁迫导致苗木明显衰老,虽然‘中红杨’和‘2025’叶片可溶性蛋白质的变化与质膜相对透性为无显著相关,但相关性为正,可能与“旱胁蛋白”或“逆境蛋白”的生成有关。通过生理指标的隶属函数平均值对3个杨树品种进行综合评价,表明抗旱能力为‘2025’>‘中红杨’>‘全红杨’,三者间差异不显著。灰色关联分析表明,渗透调节物质游离脯氨酸与3个杨树品种的抗旱性关系较密切,其次是可溶性蛋白质,而质膜相对透性对三者抗旱性的影响较小。可见,叶片中游离脯氨酸含量的变化及其对膜系统稳定的影响是反映3个杨树品种抗旱能力差异的主要指标。
[1]冯玉龙,张亚杰,朱春全.根系渗透胁迫时杨树光合作用光抑制与活性氧的关系[J].应用生态学报,2003,14(8):1213-1217.
[2]胡化广,张振铭,李芳芳.干旱胁迫对两种结缕草草坪质量和生理特征的影响[J].上海农业学报,2014,30(5):109-113.
[3]黄承玲,陈训,高贵龙.3种高山杜鹃对持续干旱的生理响应及抗旱性评价[J].林业科学,2011,47(6):48-55.
[4]邓丽娟,沈红香,姚允聪.观赏海棠品种对土壤干旱胁迫的响应差异[J].林业科学,2011,47(3):25-32.
[5]张莉,续九如.水分胁迫下刺槐不同无性系生理生化反应的研究[J].林业科学,2003,39(4):162-167.
[6]尤丽佳,郭新波,付雪睛,等.干旱胁迫对转拟南芥Atpsy基因生菜的影响[J].上海农业学报,2014,30(5):38-43.
[7]孔祥生,张妙霞,王学永,等.水分胁迫下2个牡丹品种生理生化差异比较[J].林业科学,2011,47(9):162-167.
[8]李合生.植物生理生物实验理论与技术[M].北京:高等教育出版社,2000:134-200.
[9]高俊风.植物生理学实验指导[M].北京:高等教育出版社,2006.
[10]周江,裴宗平,胡佳佳,等.干旱胁迫下3种岩石边坡生态修复植物的抗旱性[J].干旱区研究,2012,29(3):440-444.
[11]邓聚龙.灰色预测与决策〔M〕.武汉:华中科技大学出版社,1986:103-108.
[12]北京林业大学.统计学原理与林业统计学[M].北京;中国林业出版社,1986.
[13]史玉炜,王燕凌,李文兵,等.水分胁迫对刚毛柽柳可溶性蛋白、可溶性糖和脯氨酸含量变化的影响[J].新疆农业大学学报,2007,30(2):5-8.
[14]胡尚连,曹颖,段宁,等.不同类型竹种抗寒性的灰色关联与聚类分析[J].福建林学院学报,2010,30(4):327-332.
[15]柯世省,杨敏文.水分胁迫对云锦杜鹃抗氧化系统和脂类过氧化的影响[J].园艺学报,2007,34(5):1217-1222.
[16]高建社,孙楠,马宝有,等.白杨派几个无性系抗旱性比较研究[J].西北林学院学报,2007,22(3):64-66.
[17]崔晓涛,杨玲,沈海龙.干旱胁迫对新西伯得亚银白杨抗氧化系统和幼苗生长的影响[J].植物研究,2009,29(6):701-707.
[18]王晶英,赵雨森,王臻,等.干旱胁迫对银中杨生理生化特性的影响[J].水土保持学报,2006,20(1):197-200.
[19]魏良民.几种旱生植物碳水化合物和蛋白质变化的研究[J].干旱区研究,1991,8(4):38-41.
[20]康俊梅,杨青川,樊奋成.干旱对苜蓿叶片可溶性蛋白的影响[J].草地学报,2005,13(3):199-202.
[21]王海珍,梁宗锁,韩蕊莲,等.土壤干旱对黄土高原乡土树种水分代谢与渗透调节物质的影响[J].西北植物学报,2004,24(10):1822-1827.
[22]孙国荣,张睿,姜丽芬,等.干旱胁迫下白桦实生苗叶片的水分代谢与部分渗透调节物质的变化[J].植物研究,2001,21(3):431-415.
[23]胡玉净,邓丽娟,张杰,等.干旱胁迫下外源ABA对观赏海棠叶片可溶性蛋白的脱水素积累的影响[J].林业科学,2012,48(4):35-42.
[24]李晶,阎秀峰,袓元刚.低温胁迫下红松幼苗活性氧的产生及保护酶的变化[J].植物学报,2000,42(2):148-152.
[25]张明生,彭忠华,谢波,等.甘薯离体叶片失水速率及渗透调节物质与品种抗旱性的关系[J].中国农业科学,2004,37(1):152-156.
[26]马讳,王彩云.几种引进冷季型草坪草的生长及抗旱生理指标[J].草地科学,2001,18(2):57-61.
[27]喻晓丽,邸雪颖,宋丽萍.水分胁迫对火炬树幼苗生长和生理特性的影响[J].林业科学,2007,43(11):57-61.
(责任编辑:闫其涛)
Change of leaf membrane permeability and some osmotic regulation substances of 3 poplar varieties under drought stress
YANG Shu-hong1,ZHU Di2,REN Yuan-yuan1,ZHU Yan-lin1
(1Henan Academy of Forestry,Zhengzhou 450008,China;2Forestry College of Fujian Agriculture and Forestry University,Fuzhou 350002,China)
The paper reports the changes of relative plasma membrane permeability and the contents of some osmotic regulation substances such as soluble sugar,soluble protein and proline in leaves and the evaluation of drought resistance of poplar‘2025’and its two bud mutation varieties under continuous drought stress of soil.The results showed that with continuous drought stress,the relative plasma membrane permeability all increased obviously,and the‘Quanghong’poplar were always higher than‘Zhonghong’and‘2025’.The total soluble protein and free proline in leaves of the 3 varieties had a tendency to reduce,but the soluble sugar content kept in a higher level.There was a significant positive correlation between the soluble sugar content and the relative plasma membrane permeability(1>|r|≥0.7),which was good for the plants to adapt to drought stress and obtain drought-resistant ability.There was a significant positive correlation between the change of proline content in leaves of poplar‘2025’and the permeability of plasma membrane(1>|r|≥0.7),and the membrane system was relative stable.There were extremely significant differences between the control group and the treatment group of the 3 varieties(P<0.01).It was concluded that the drought resistant ability was poplar‘2025’>‘Zhonghong’>‘Quanhong’by membership function method,and there were no significant difference(P>0.05)among the 3 varieties.The sensitivity of each index to drought stress was analyzed by the grey correlation method,results showed that the free proline was the main index reflecting the drought resistant ability of 3 poplar varieties.
Poplar variety;Drought stress;Osmotic regulation;Drought resistant ability
S792.11.01
A
1000-3924(2016)06-118-06
2015-09-24
项目来源:河南省基础与前沿技术研究计划项目(132300413223);河南省林科院基本科研业务费项目(2016JB02-001)
杨淑红(1975—),女,本科,高级工程师,研究方向为城市林业与园林绿化。E-mail:yangshh0315@163.com
