水旱共作体系中生物耕作对菜田土壤矿化菌数量及矿化强度的影响
2017-01-12郑宪清张翰林李双喜张娟琴陈新兵吕卫光陶晓斌
郑宪清,张翰林,李双喜,宋 科,张娟琴,陈新兵,吕卫光*,陶晓斌
(1上海市农业科学院生态环境保护研究所,2农业部上海农业环境与耕地保育科学观测实验站,上海 201403;3上海市上海农场,上海 224151;4上海瀛千生态农业有限公司,上海 202158)
水旱共作体系中生物耕作对菜田土壤矿化菌数量及矿化强度的影响
郑宪清1,2,张翰林1,2,李双喜1,2,宋 科1,2,张娟琴1,2,陈新兵3,吕卫光1,2*,陶晓斌4
(1上海市农业科学院生态环境保护研究所,2农业部上海农业环境与耕地保育科学观测实验站,上海 201403;3上海市上海农场,上海 224151;4上海瀛千生态农业有限公司,上海 202158)
针对传统蔬菜种植中高复种指数、强机械扰动和大量施肥导致土壤功能微生物菌群数量下降、功能降低和土传病害频发等问题,在水旱共作的立体生态种养模式下,长期定位试验田中添加蚯蚓,探索生物耕作对菜田土壤氨化和硝化菌数量及强度的影响。3年的试验结果表明:同一耕作层内氨化细菌数量随生物耕作时间延长呈现先增加后降低的趋势,最高值达2 332.1万个/g干土;硝化细菌数量随耕作时间延长呈上升趋势,与无蚯蚓的免耕处理相比,增加7.89—12.73倍,最高值达3.872万个/g干土;硝化作用强度在生物耕作的第二年达到最高值99.99%;5—20 cm耕作层硝化菌数量和强度均高于0—5 cm土层;在0—5 cm土层主要是土壤全氮和速效钾起主导作用,影响因子综合解释率达80.2%—99.5%;5—20 cm土层全氮、pH和速效磷共同影响硝化强度,影响因子综合解释率达93.8%—96.9%;生物耕作深度和耕作时间对氨化菌和硝化菌数量均有显著的交互影响。
菜园土壤;土壤微生物;矿化作用;生物耕作
氮素是植物体的主要组成成分之一,也是影响作物生长发育的重要因子之一,在土壤中主要以有机氮形式存在,占土壤全氮量的95%以上。而土壤中有机态氮必须矿化成氨态氮和硝态氮等无机态氮[1]才能被植物吸收和利用,因此,有机氮的矿化率决定土壤中可利用无机氮的含量,土壤氨化菌与硝化菌是土壤中有机氮分解的“引擎”,对有机态氮的矿化起关键作用[2-3]。土壤氮素矿化过程受控于多种因子,如土壤动物、微生物、植物种类、环境因子和人类活动等,这些因子单独或者协同作用控制氨化菌和硝化菌的数量和强度,进而控制氮素的矿化速率。有研究发现常规机械耕作,破坏土壤结构,降低物种丰度和均匀度,也显著降低细菌多样性[4]。钟文辉等[5]认为土壤免耕相比于常规耕作显著提高了土壤细菌的多样性指数,而显著影响土壤氮素循环。一些学者提出了“生物耕作”的方法[6],即利用土壤动物蚯蚓在生活和新陈代谢过程中的取食、排泄和掘穴等生命活动,调节农田微生态环境,改善微生物区系,实现养分良性循环与适时供给,促进土壤物质循环和能量转化[7]。研究表明,蚯蚓活动可以增加土壤微生物总量[8-12],增强微生物活性[13-14],改变土壤微生物群落组成,提升土壤酶活性,从而增大土壤氮矿化速率和硝化作用,然而,目前关于蚯蚓耕作对菜田土壤氮素转化过程的功能微生物影响的研究较少[15]。本研究通过在菜园设置水旱共作的立体种养模式,并引入蚯蚓,设置蚯蚓防逃的生物耕作的定位试验,研究水旱共存的环境下蚯蚓活动对菜园土壤氮转化功能菌的影响,为立体种养模式中的生物耕作环节进一步研究与应用提供基础数据。
1 材料与方法
1.1 试验设计
本研究在立体生态种养模式下开展,具体分为3个部分,1)旱作部分,畦面土壤上种植旱作蔬菜;2)水旱交界部分,土壤中含水量适中处养殖蚯蚓;3)水作部分,垄沟内放水养殖水产生物。3个部分组合在一个系统里按照空间和时间错位协同原理同时进行(图1)。
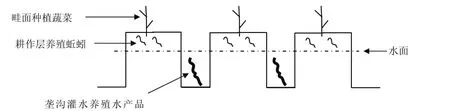
图1 立体生态种养模式示意图Fig.1 Sketch map of ecological planting and breeding pattern
生物耕作是指在土壤中投放威廉环毛蚓(Pheretima guillelmi),利用蚯蚓自身的生命活动对土壤的耕翻扰动代替传统的机械耕作,使土壤疏松程度达到作物种植要求的模式。
本研究的生物耕作试验开始于2009年,试验小区设在崇明县三星镇西新村菜田内,设置以下处理:
(1)免耕(对照,简写NT):不进行任何形式的耕作处理,土壤中原有的蚯蚓采用灌水腌泡的方式将其驱除或杀灭。
(2)生物耕作处理:试验样地进行生物耕作之前,采取常规耕作。驱除原有蚯蚓后,投放试验用蚯蚓25条/m2,试验进行3年,每年试验开始时进行蚯蚓数量测定(多于25条/m2则移除多的部分,少则添加至25条/m2)。试验进行一年称为生物耕作一年(Biological tillage for one year,简写BT1),试验进行2年称为生物耕作2年(Biological tillage for two years,简写BT2),试验进行3年称为生物耕作3年(简写BT3)。每个处理设3个重复,小区面积240 m2,小区周围开设深沟并灌水至水面距离地表20 cm左右,防止蚯蚓逃跑。各处理均实行西瓜-花菜的轮作模式:西瓜以商品有机肥(含有机质413.4 g/kg,N 17.1 g/kg,P2O512.4 g/kg,K2O 12.3 g/kg)18 t/hm2作基肥,复合肥(15-15-15)375.0 kg/hm2(90%做基肥,10%做追肥),均撒施于地表;花菜以商品有机肥(同上)15 t/hm2作基肥,复合肥(15-15-15)750 kg/hm2(60%做基肥,40%做追肥),均撒施于地表。
1.2 土壤样品采集
2011年西瓜收获后以“S”形多点采集0—5 cm和5—20 cm土层土壤样品,鲜样立即测试土壤氨化菌数量、硝化菌数量、氨化作用强度和硝化作用强度等土壤微生物区系指标,留一部分土壤自然风干留存,用于测定土壤基本理化性状。
1.3 测定项目与方法
1.3.1 土壤理化性状指标测定
测试指标包括有机质、全氮、全磷、全钾、速效氮、速效磷、速效钾、pH、含水量,测试方法参照鲁如坤[16]的土壤农业化学分析方法,结果见表1。

表1 生物耕作对土壤理化性状变化的影响Table 1 Effects of biological tillage on soil physical and chemical properties
1.3.2 土壤微生物区系指标测定
土壤氨化菌和硝化菌数量采用最大或然值法,土壤氨化强度和硝化强度采用悬液培养法测定[17]。
1.3.3 土壤中蚯蚓数量测定
采用福尔马林法,测定1 m2范围内20 min蚯蚓爬出量[18]。
1.4 数据处理
试验数据应用Excel和SPSS 13.0统计软件进行统计分析、方差分析和线性回归分析。
2 结果与分析
2.1 水旱共作模式下生物耕作对菜田土壤矿化菌数量及强度的影响
2.1.1 菜田土壤中氨化菌数量的变化
生物耕作时间的不同影响了不同深层土壤氨化细菌的数量。经过3年的生物耕作试验,可以发现随着生物耕作时间的延长,无论是土壤表层还是亚表层,氨化细菌的数量均呈现先升高后降低的变化趋势,在0—5 cm土层,在生物耕作第2年达到最大值,即1 310.3万个/g干土,相对于免耕升高28倍;在5—20 cm土层,在生物耕作一年后达到最大值,即2 332.1万个/g干土,相对于免耕升高26倍,5—20 cm土层氨化菌数量稍高于0—5 cm土层(表2)。
2.1.2 菜田土壤中硝化菌数量的变化
从表2中可以看出,在水旱共作的立体种养模式下,采用蚯蚓进行生物耕作的3年内,随着生物耕作时间的延长,两个耕作层硝化细菌数量均呈上升趋势,即BT3>BT2>BT1>NT,且到耕作第3年较前两年差异达到显著,在0—5 cm土层,硝化细菌数量由免耕处理的0.146万个/g干土上升至生物耕作3年处理的1.152万个/g干土,在5—20 cm土层,同样从免耕到生物耕作3年,硝化菌数量从0.304万个/g干土上升至3.872万个/g干土,分别增加7.89倍和12.73倍,说明生物耕作时间对土壤硝化细菌数量的提升有促进作用。5—20 cm土层硝化菌数量均高于0—5 cm土层,也说明耕作土层深度不同,硝化细菌的数量变化也不一样。

表2 生物耕作后表土(0—5 cm)和亚表层(5—20 cm)中氨化和硝化菌数量和强度的变化Table 2 Variations of ammonifier and nitrifier quantities and ammonifying and nitrifying capacities in 0—5 cm and 5—20 cm soil layers after biological tillage
2.1.3 菜田土壤微生物氨化作用强度的变化
氨化作用是氨化细菌分解有机含氮化合物释放出铵的过程,是土壤氮素转化过程中的第一步,其强度的强弱影响着土壤氮素转化速率。从表2中可以看出,虽然氨化细菌数量在生物耕作的3年里变化幅度很大,但是其作用强度并没有明显的变化,即随着时间的延长,氨化作用强度无显著性差异,基本保持在0.05 mg/100 mL,说明氨化菌数量多寡不是氨化作用强度的唯一影响因子,在某些特定的环境下,氨化作用强度可能除了受氨化菌数量的影响外还受如土壤水分含量和温度等其他因子的影响。
2.1.4 菜田土壤微生物硝化作用强度的变化
硝化强度是指亚硝酸氧化细菌将NH4+-N氧化成NO3--N的作用,土壤环境因子对硝化作用的强度有一定的影响。与土壤中氨化作用强度不同,硝化作用强度随生物耕作时间的延长有增加的趋势,在生物耕作的第二年达到最高值99.99%以上,并在接下来的时间里趋于平稳。硝化作用强度与硝化细菌的数量变化有一致性,5—20 cm土层高于0—5 cm土层(见表2)。
2.2 水旱共作立体种养模式下生物耕作对菜田土壤矿化菌数量及强度的影响分析
2.2.1 水旱共作模式下生物耕作对土壤氨化和硝化的影响
如表3所示,在水旱共作的立体种养模式下,随着生物耕作时间的延长,在0—5 cm(R2=0.874)和5—20 cm(R2=0.925)土层硝化菌数量均极显著提升(P<0.01),同徐晓燕等[19]的研究结果相一致,即接种蚯蚓一定时间处理后,会使土壤中微生物类群的数量增加。生物耕作使得土壤硝化菌数量增加,进而显著地提升土壤硝化强度,在0—5 cm土层达显著,在5—20 cm土层呈极显著,也许是因为经蚯蚓腹腔内酶以及酶对土壤养分转化更有利于氨化菌和硝化菌的生长繁殖[20],蚯蚓对土壤翻动形成的空隙提供的有氧环境有助于蚯蚓活性[21]。同时,刘婷等[22]和刘德辉等[23]分别利用赤子爱胜蚓和秉氏环毛蚓的研究发现添加蚯蚓活动能够使土壤或添加物料的pH趋于中性,而中性的环境较利于氨化菌和硝化菌等功能菌群的生长。本研究中添加的威廉环毛蚓由于体型较大,达到5 g左右,它们的活动能够更大强度的耕翻与扰动土壤,形成较好的团粒结构和土壤孔隙度,为土壤微生物的活动提供了充足的有氧环境;从而增强微生物的活性,最终提升土壤硝化作用强度。而从表3中看出生物耕作时间对氨化菌数量提升作用不显著,也没有提升氨化作用强度。或许因为在生物耕作条件下,生物耕作深度、耕作时间不仅单独对土壤氨化和硝化细菌数量产生影响,它们的交互作用也影响氨化和硝化菌数量。但只有耕作时间对硝化强度产生显著影响,而没有对氨化强度产生影响(表4)。也有研究认为,虽然土壤通气状况影响氨化细菌的优势种群,但通气状况好坏程度对氨化作用强度的影响不大[24]。
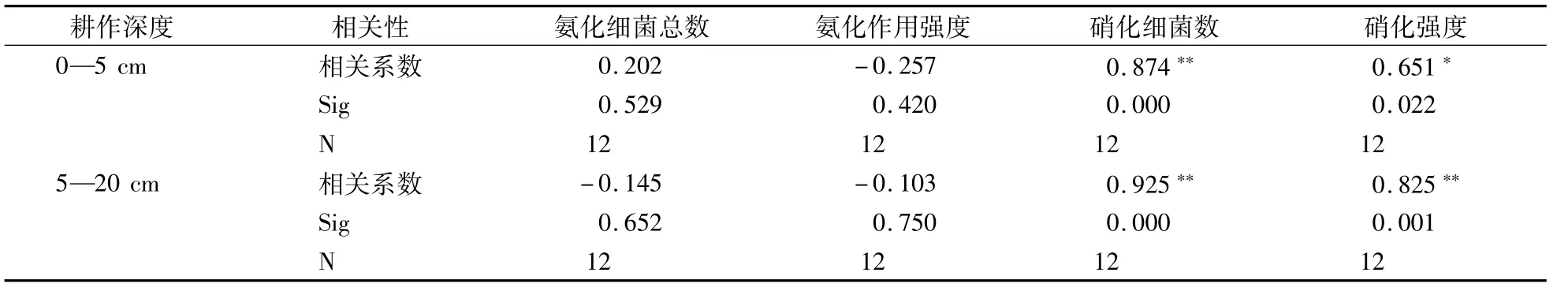
表3 生物耕作与氨化过程和硝化过程的相关性分析Table 3 Correlation between biological tillage and ammonification/nitrification process
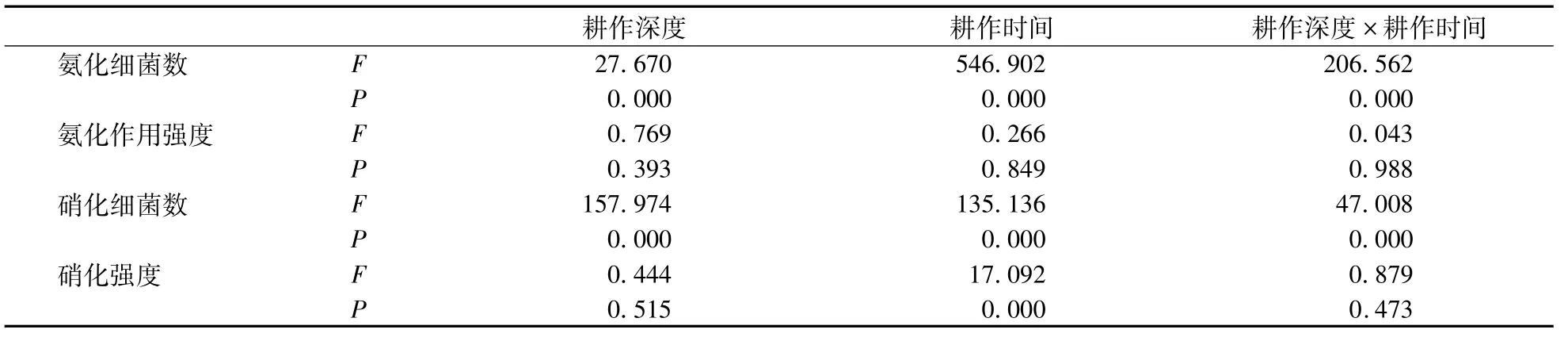
表4 氨化和硝化强度的影响因子的统计分析(R2=0.993)Table 4 Statistic analysis of factors influencing ammonifying and nitrifying capacities
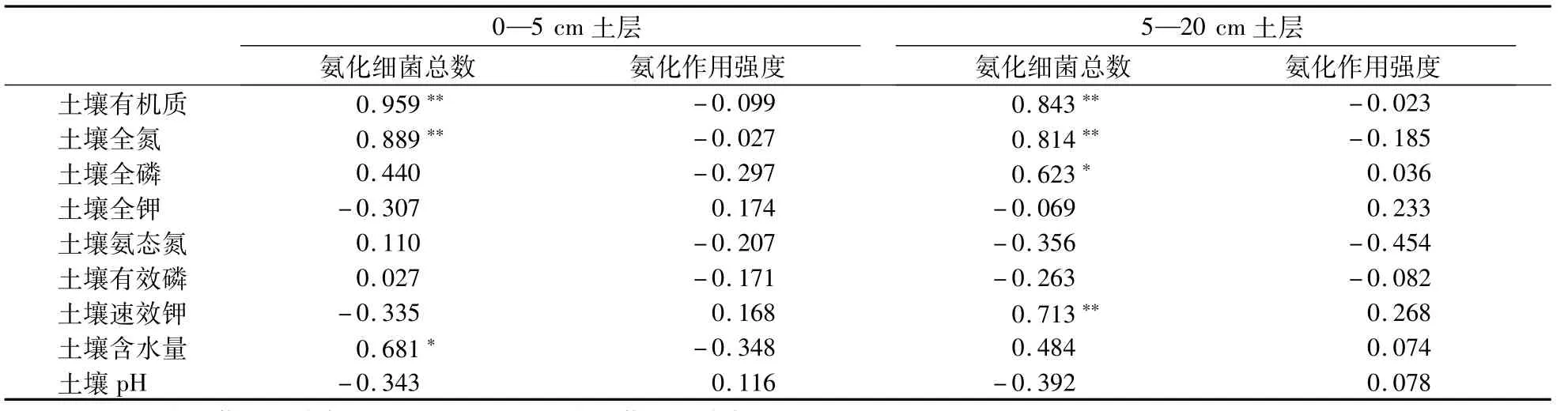
表5 土壤性质与不同土层氨化细菌和氨化强度之间的相关性Table 5 Correlations of soil properties with different soil layers’ammonifiers and ammonifying capacity
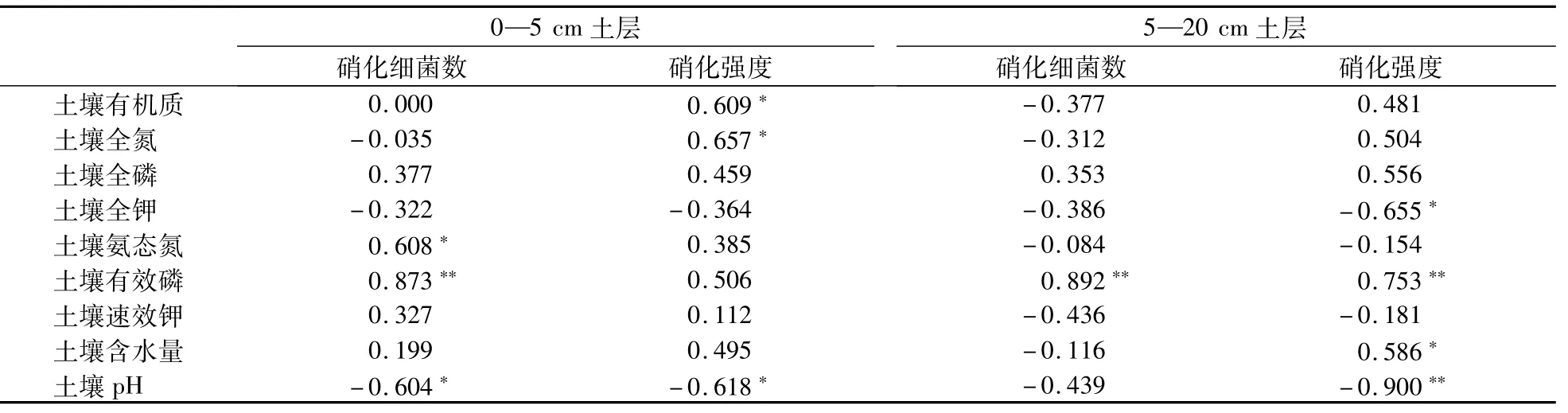
表6 土壤性质与不同土层硝化细菌和硝化强度之间的相关性Table 6 Correlations of soil properties with different soil layers’nitrifiers and nitrifying capacity
2.2.2 生物耕作下土壤性质对土壤氨化和硝化的影响
在相同的水热因子、植被和施肥外界环境下,生物耕作过程中蚯蚓通过自身活动影响土壤的水、肥、气、热状况变化,从而引起土壤氮素转化发生一系列变化[25-26],也影响了土壤氨化和硝化菌的生长、繁殖,其数量出现显著性变化,而且随着耕作时间延长,土壤硝化强度明显加强(P<0.05),而对氨化作用强度影响不明显。显然土壤有机质、氮、磷、钾等养分以及土壤含水量是影响土壤微生物数量和活性的重要因素[27-29],但是在不同土层深度上主导因素不同。如表5所示,在0—5 cm土层,土壤有机质和全氮对氨化细菌数量的增加有极大地促进作用(P<0.01),土壤含水量也有较强的作用(P<0.05),说明土壤有机物质做为氨化和硝化菌生长的营养源,进而显著影响其数量的变化[30],而本研究中,在5—20 cm土层,除了土壤有机质和全氮极大地促进氨化菌数量的增加外,土壤速效钾与氨化菌数量的增加有极显著的相关性(P<0.01),同时土壤全磷也有较强的作用(P<0.05)。可能由于环毛蚓属上食下居型,主要以表层土壤颗粒和有机物的混合物为食,活动于5—15 cm矿质土层,改变了5—20 cm土层有机质含量,调整了此层的土壤碳氮比等养分配比,从而促进土壤氨化菌和硝化菌生长与繁殖[31-34]。也有学者认为氨化细菌不仅受土壤全氮、全磷的影响较大[35],也随着土壤水分的不断增加,氨化细菌数量也随之增长[36],土壤中氨化细菌数量直接反映了氨化作用的强度[37],而本研究中5—20 cm土层各影响因子与氨化强度均无显著相关性,可能是因为本研究中土壤含水量过高有关。前人研究发现除了养分,含水量和孔隙度外,土壤pH也是影响土壤硝化细菌数量和活性的关键因子,从表5和表6可以看出,土壤pH没有显著影响土壤氨化菌数量和强度而显著影响硝化菌数量和强度,因为在土壤pH为6.4—8.3时,硝化作用都会强烈进行,而氨化菌更趋向于中性条件,本试验的土壤pH较高,均在8.3附近,因此不利于氨化过程,但不抑制硝化过程。
为进一步探究生物耕作过程中土壤理化性状对氨化和硝化过程的贡献率,以理化性状值为自变量,以氨化菌数和硝化菌数及强度为因变量,经逐步回归分析,得最优回归方程如下(前3式为0—5 cm土层,后3式为5—20 cm土层;式中X1、X2、X4、X5、X6、X7、X8和X9分别代表有机质、全氮、全钾、速效氮、速效磷、速效钾、含水量和pH):

分析结果显示,在0—5 cm土层中,生物耕作通过影响有机质、全钾和速效氮等含量而影响氨化菌数量;通过影响速效磷和速效钾变化,而影响土壤硝化菌数量变化;通过影响土壤全氮和速效钾的量,而影响土壤强硝化强度,其影响因子综合解释率分别为99.5%、92.9%和80.2%。在5—20 cm土层生物耕作通过影响速效钾、全氮和全磷,而影响氨化菌数量;通过影响速效磷、含水量和速效氮量,而影响土壤硝化菌数量,通过影响pH、速效磷和全氮,而影响硝化强度,其影响因子综合解释率分别达96.9%、93.8%和94.1%。说明生态种养时间的延长有利于土壤生物活性的提升[38],而经逐步回归分析没有发现某些养分元素对氨化强度有主导性影响作用,从而说明此条件下生物耕作对土壤氨化作用影响有限。
3 结论
(1)生物耕作时间对菜园土壤氮转化功能菌数量产生影响,3年的试验结果显示,氨化细菌数量呈现先增加后降低的趋势,硝化细菌数量随耕作时间延长呈上升趋势,均表现出5—20 cm土层数量高于0—5 cm土层。同时,生物耕作时间影响菜园土土壤硝化作用强度,硝化作用强度随生物耕作时间的延长有增加的趋势,在生物耕作的第二年达到最高值99.99%以上,5—20 cm土层高于0—5 cm土层。但对氨化作用强度无显著影响。
(2)经逐步回归分析,生物耕作对硝化作用强度的影响因土层不同而有差异,0—5 cm土层主要是土壤全氮和速效钾起主导作用,影响因子综合解释率达80.2%—99.5%。5—20 cm土层全氮、pH和速效磷共同影响硝化强度,影响因子综合解释率达93.8%—96.9%。在生物耕作的3年里,耕作深度和耕作时间对氨化菌和硝化菌数量有着明显的交互影响作用,达到了极显著程度。而对土壤氨化和硝化强度的交互作用不显著。
[1]李生秀.中国旱地土壤植物氮素[M].北京:科学出版社,2008.
[2]陈蓓,张仁涉.免耕与覆盖对土壤微生物数量及组成的影响[J].甘肃农业大学学报,2004,39(6):634-638.
[3]李忠佩,吴晓晨,陈碧云,等.不同利用方式下土壤有机碳转化及微生物群落功能多样性变化[J].中国农业科学,2007,40(8):1712-1721.
[4]ALVEY S,YANG C H,BUERKERT A.Cereal/legume rotation effects on rhizosphere bacterial community structure in west african soils[J]. Biology and Fertility of Soils,2003,37(2):73-82.
[5]钟文辉,蔡祖聪.土壤管理措施及环境因素对土壤微生物多样性影响研究进展[J].生物多样性,2004,12(4):456-46.
[6]郑宪清,李双喜,吕卫光,等.生物耕作对蔬菜田土壤养分及酶活性的影响[J].植物营养与肥料学报,2012,18(5):1177-1184.
[7]TIAN G,OLIMAH J A,ADEOYE G O,et al.Regeneration of earthworm populations in a degraded soil by natural,planted fallows under humid tropical conditions[J].Soil Science Society of America Journal,2000,64(1):222-228.
[8]BLAIR J M,PARMELEE R W,ALLEN M F,et al.Changes in soil N pools in response to earthworm population manipulations in agroecosystems with different N sources[J].Soil Biology and Biochemistry,1997,29(3/4):361-367.
[9]HENDRIX P F,PETERSON A C,BEARE M H,et al.Long-term effects of earthworms on microbial biomass nitrogen in coarse and fine textured soils[J].Appl Soil Ecol,1998,9(1/2/3):375-380.
[10]WOLTERS V,JOERGENSEN R G.Microbial carbon turnover in beech forest soils worked by Aporrectodea caliginosa(Savigny)(Oligochaeta:Lumbricidae)[J].Soil Biol.Biochem,1992,24(24):171-177.
[11]ARAUJO Y,LUIZ F J,BARROS E.Effect of earthworm addition on soil nitrogen availability,microbial biomass,litter decomposition in mesocosms[J].Biology and Fertility of Soils,2004,39(3):146-152.
[12]PASHANASI B,LAVELLE P,ALEGRE J.Effect of inoculation with the endogeic earthworm Pontoscolex corethrurus on soil chemical characteristics and plant growth in a low input agricultural system of Peruvian Amazonia[J].Soil Biol Biochem,1993,24(92):1655-1659.
[13]ZHANG B G,LI G T,SHEN T S,et al.Changes in microbial biomass C,N,and P and enzyme activities in soil incubated with the earthworms Metaphire guillelmi or Eisenia fetida[J].Soil Biology and Biochemistry,2001,32(14):2055-2062.
[14]SCHEU S,SCHLITT N,TIUNOV A V,et al.Effects of the presence and community composition of earthworms on microbial community functioning[J].Oecologia,2002,133(2):254-260.
[15]BOHLEN PJ,SCHEU S,HALE CM,et al.Non-native invasive earthworms as agents of change in northern temperate forests[J].Front.Ecol. Environ,2004,2(8):427-435.
[16]鲁如坤.土壤农业化学分析方法[M].北京:中国农业科技出版社,2000:146-197.
[17]许光辉.土壤微生物分析方法手册[M].北京:农业出版社,1986:110-240.
[18]李文凤,张晓平,梁爱珍,等.免耕对黑土蚯蚓数量和土壤密度的影响[J].农业系统科学与综合研究,2007,23(4):489-493.
[19]徐晓燕,高晓玲,郑琪,等.不同季节蚯蚓活动对土壤微生物类群数量的影响[J].安徽农业科学,2011,39(11):6478-6479.
[20]曹佳,王冲,皇彦,等.蚯蚓对土壤微生物及生物肥力的影响研究进展[J].应用生态学报,2015,26(5):1579-1586.
[21]ZHANG B G,LI G T,SHEN T S,et al.Changes in microbial biomass C,N and P and enzyme activities in soil incubated with the earthworms Metaphire guillelmi or Eisenia fetida[J].Soil Biology and Biochemistry,2000,32(14):2055-2062.
[22]刘婷,任宗玲,张池,等.蚯蚓堆置处理对农业有机废弃物的化学及生物学影响的主成分分析[J].应用生态学报,2012,23(3):779-784.
[23]刘德辉,胡锋,胡佩.蚯蚓活动对红壤磷素有效性的影响及其活化机理研究.生态学报,2003,23(11):2299-2306.
[24]欧惠平,沈方科,陈秋实,等.免耕和稻草还田对稻田土壤氮素转化强度的影响[J].土壤通报,2012,43(4):877-882.
[25]高亚军,李生秀,李世清,等.农田秸秆覆盖对冬小麦水氮效应的影响[J].应用生态学报,2005,16(8):1450-1454.
[26]刘建国,卞新民,李彦斌,等.长期连作和秸秆还田对棉田土壤生物活性的影响[J].应用生态学报,2008,19(5):1027-1032.
[27]李玲玲,黄高宝,张仁陟,等.免耕秸秆覆盖对旱作农田土壤水分的影响[J].水土保持学报,2005,19(5):95-96.
[28]黄高宝,李玲玲,张仁陟,等.免耕秸秆覆盖对旱作麦田土壤温度的影响[J].干旱地区农业研究,2006,24(5):1-4.
[29]唐劲驰,周波,黎健龙,等.蚯蚓生物有机培肥技术(FBO)对茶园土壤微生物特征及酶活性的影响[J].茶叶科学,2016,36(1):45-51.
[30]逄蕾,黄高宝.不同利用方式对旱地土壤有机碳转化的影响[J].水土保持学报,2006,20(3):110-113.
[31]樊丽琴,南志标,沈禹颖,等.保护性耕作对黄土高原豌豆田土壤微生物量碳的影响[J].草原与草坪,2005(4):51-54.
[32]范丙全,刘巧玲.保护性耕作与秸秆还田对土壤微生物及其溶磷特性的影响[J].生态农业学报,2005,13(3):130-132.
[33]王启兰,王长庭,杜岩功,等.放牧对高寒嵩草草甸土壤微生物量碳的影响及其与土壤环境的关系[J].草业学报,2008,17(2):39-41.
[34]郭彦军,韩建国.农牧交错带退耕还草对土壤酶活性的影响[J].草业学报,2008,17(5):16-23.
[35]王丽宏,杨光立,曾昭海,等.稻田冬种黑麦草对饲草生产和土壤微生物效应的影响[J].草业学报,2008,17(2):157-161.
[36]韩晓彬.黑土有机质功效的研究[J].农业系统科学与综合研究,2001,17(4):256-259.
[37]刁治民.高寒草地的微生物氮素生理群区系研究[J].土壤,1996(1):49-53.
[38]周雪芳,朱晓伟,陈泽恺,等.稻蛙生态种养对土壤微生物及无机磷含量的影响[J].核农学报,2016,30(5):971-977.
(责任编辑:程智强)
Effects of biological tillage on mineralization bacteria and strength of garden soil in furrow aquaculture
ZHENG Xian-qing1,2,ZHANG Han-lin1,2,LI Shuang-xi1,2,SONG Ke1,2,ZHANG Juan-qin1,2,
CHEN Xin-bing3,LÜ Wei-guang1,2*,TAO Xiao-bin4
(1Institute of Eco-Environment and Plant Protection,Shanghai Academy of Agricultural Sciences,2Scientific Observation and Experimental Station for Agricultural Environment and Land Conservation at Shanghai,Ministry of Agriculture,Shanghai 201403,China;3Shanghai Farm,Shanghai 224151,China;4Shanghai Yingqian Eco-Agriculture Company Limited,Shanghai 202158,China)
In order to deal with problems of high-cropping-index conventional vegetable planting such as soil functional microbe quantity decreasing,their function declining and soil-borne disease frequently occurring caused by strong mechanical disturbance and mass fertilizer application,a long-term location experiment was carried out,in which earthworms were added into test plots with furrow aquaculture,so as to investigate the effects of biological tillage on ammonifiers and nitrifiers of vegetable soil.The 3-year experimental results showed that in the same plow layer the ammonifiers firstly increased and then decreased,and the maximum number was 23.321 million per gram dry soil;The nitrifiers presented a rising trend with time and the maximum number reached 38 720 per gram dry soil,increasing by 7.89—12.73 times over that under the treatment of non-tillage without earthworm;The nitrifying capacity reached the highest value of 99.99%in the second year of biological tillage;The nitrifier quantity and nitrifying capacity in 5—20 cm soil layer were greater than those in 0—5 cm soil layer;In 0—5 cm soil layer the total nitrogen and readily available potassium played a leading role and their comprehensive influencing factor interpretation rate reached 80.2%—99.5%;In 5—20 cm soil layer the total nitrogen,pH and readily available phosphorus jointly influenced the nitrifying capacity and their comprehensive influencing factor interpretation rate reached 93.8%—96.9%;Both biological tillage depth and time had a significant interaction on both ammonifier and nitrifier quantities.
Garden soil;Soil microbe;Mineralization;Biological tillage
S154
A
1000-3924(2016)06-075-07
2016-04-21
上海市农委青年人才成长计划[沪农青字(2015)第1-23号];上海市科委科技攻关项目(14391901502);上海市科委成果转化项目(133919N1400);上海市农委重点攻关项目[沪农科攻字(2011)第6-2号]
郑宪清(1980—),男,硕士,助研,主要从事土壤生态学研究。Tel:021-62202486,E-mail:zxqfxf@163.cm
*通信作者:吕卫光(1972—),男,博士,研究员,主要从事植物营养学及土壤生态环境学研究。E-mail:lwei1217@sina.com.cn
