Prolificity of Portuguese Serrana Goats between 1987 and 2015
2017-01-06SacotoSandraSimesJo
Sacoto Sandra, Simões João
1CECAV, Department of Animal Science, University of Trás-os-Montes e Alto Douro
2CECAV, Department of Veterinary Science, University of Trás-os-Montes e Alto Douro
Prolificity of Portuguese Serrana Goats between 1987 and 2015
Sacoto Sandra1*, Simões João2
1CECAV, Department of Animal Science, University of Trás-os-Montes e Alto Douro
2CECAV, Department of Veterinary Science, University of Trás-os-Montes e Alto Douro
ARTICLE INFO
Article history:
Received 2016
Received in revised form 2016
Accepted 2016
Available online 2016
Mountain agro-pastoralism
Objective:To evaluate the prolificity of Serrana goats and their four ecotypes (Transmontano, Jarmelista, Ribatejano and Serra) as well its monthly distribution according to parity.Methods:Data (n=316 610 parturitions) were recorded from 1987 to 2015 and obtained from the pedigree book of Serrana goats for all four ecotypes. The period (month and year) of parturition, females’ parity and normal or abnormal parturitions, including abortions, were considered as independent variables. The dependent variable prolificity was calculated by the following formula: number of born kids/parturition.Results:It was observed an average prolificity of 1.47; from parity order 1 to parity order 5 prolificity increased from 1.33 to 1.57 (P<0.001). The Ribatejano (n=45,925), Jarmelista (n=31 723), Transmontano (n=238 106) and Serra (n=856) ecotypes showed prolificity averages of 1.60, 1.54, 1.44, and 1.35 (P<0.001), respectively. Globally, and regarding the monthly kidding distribution for multiparous and nulliparous females of Transmontano, Jarmelista and Ribatejano ecotypes, it was also observed for parturitions from December and forward a decrease on prolificity with the minimum value being achieved in April (P<0.001).Conclusion:These results suggest that females bred in July/August and October could be negatively affected by factors such as feed availability, environmental constraints and management conditions, with impacts on reproductive performance, namely on prolificity, which should be more investigated. Our study allows us to state that the Ribatejano and Jarmelista ecotypes presented higher prolificity than Transmontano ecotype probably due, in part, to genetic differences.
1. Introduction
Goat production is an attractive and profitable activity even when practiced is marginal and poor agricultural areas[1,2] like the majority of goat farms that exist in Mediterranean regions, including the inner areas of Portugal.
Among the reproductive parameters, prolificity is one of the most important in determining the efficiency of the system[3,4] as it is directly related to the economic viability of the flock[5,6]. The number of young born alive per kidding is an important factor in increasing productivity as it contributes more to the total weight weaned per does than the growth rate of kids[6].
The prolificity of meat or milk-meat dual purpose goat breeds, as is the case of the Portuguese Serrana breed, play an important role in the economy of animal production systems. This prolificity is primarily dependent on the genotype, and there are even sheep breeds (with Booroola gene) and goats, such as Jining Grey Chinese breed, considered extremely prolific[7].
The prolificity contributes significantly to the annual genetic gain in flocks in which selection is performed, since a lower prolificity rate implies a lower number of born kids per year, which difficult the replacement of livestock, reduces the selection pressure and increases the interval between generations. In fact, prolificity has high heritability compared with other reproductive parameters,leading to a quick response to selection[8]. Other factors may also contribute to the number of kids born by female, among which could be highlight the nutritional and feeding level, body condition, parity and females’age[3,9] or even sexual stimulation/ follicular heat and stress.
Increasing meat production is a challenge and a goal for most of the producers. This can be achieved by many ways but the analysis of the genetic potential of a specific breed and how it can be used to improve meat production is a tool that must be available to the producers is order to improve their incomes.
This study aimed to determine the prolificity of the different ecotypes of Portuguese Serrana goats and evaluate its monthly distribution from 1987 to 2015.
2. Materials and methods
2.1. Animal records
The pedigree book (https://genpro.ruralbit.com) of Serrana goat’s ecotypes Transmontano, Ribatejano, Jarmelista and Serra, between 1987 and 2015, were used in the present study. Except for Ribatejanoecotype, all remaining ecotypes were located in mountain regions.
The period (month and year) of parturition, females’ parity and normal or abnormal parturitions, including abortions, were considered. These records were obtained by technicians responsible for the pedigree book (ANCRAS-Associação Nacional de Caprinicultores da Raça Serrana) which visited farms periodically to identify the individual animals, including kids, and abnormal reproductive occurrences reported by farmers such as the number of abortionsand stillborns at parturition. Abortionswere detected normally during the second half of pregnancy.
2.2. Statistical analysis
Average prolificity was calculated considering the result of the division of parturitions by the number of fetus in each one, i.e., number of born kids/parturition.
Polynomial regressions degree 3 was used for average prolificity estimates according to each independent variable. The normal distribution of the samples was confirmed before each regression analysis. Still, the Van der Waerden test was also independently used in order to compare all groups and the P value obtained was always<0.001.
The comparison of average of prolificity between the four ecotypes of Serrana goats were performed using the Van der Waerden test.
The JMP® 10 statistical software[10] was used for all statistical analysis.
3. Results
During the period between 1987 and 2015, it was observed an average prolificity of 1.47 foetuses by kidding (n=316,610) in the Serrana goat breed.
The kidding’s of up to 1, 2 and 3 foetuses were represented in quartiles 50%, 90% and 99.5%, respectively. The remaining 0.5% was represented by births with 4 to 9 foetuses. Prolificity varied (P<0.001) according to parity (Figure 1), reaching its maximum around the 5th kidding. When were considered only deliveries without occurrence of stillbirths (and/or abortions), i.e. normal live births (n=292 951), there was an average prolificity of 1.51 for the quartiles above mentioned.
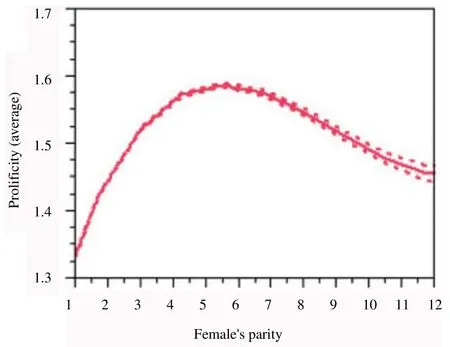
Figure 1. Estimation of average prolificity according to parity for Serrana goats
We‘re also observed different (P<0.001) average prolificity between the four studied ecotypes (Table 1), with the ecotype Ribatejano showing the highest value.
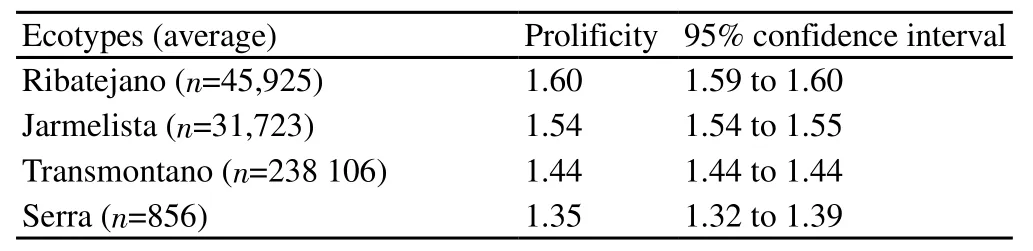
Table 1 Average prolificity of Serrana goats ecotypes (P<0.001).
The estimation of average prolificity along the years and months is reported in Figure 1 and Figure 2, respectively, and estimation average prolificity according to parturition month for Serrana goats ecotypes Transmontano, Ribatejano and Jarmelista between 1987 and 2015 is showed in Figure 3.
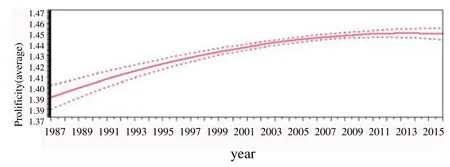
Figure 2. Estimation of average prolificity along the years for Serrana goats Transmontano ecotype between 1987 and 2015
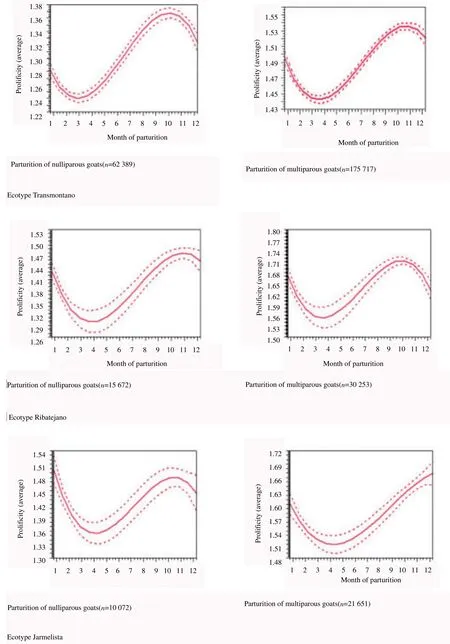
Figure 3. Estimation average prolificity according to parturition month for Serrana goats ecotypes.
4. Discussion
The average prolificity of 1.47 in Serrana goats is similar to the reported in crossbred dairy goats of the Brazilian northeastern semiarid region[5], which score was 1.49 kids by parturition.
Females’ parity had a significant effect on litter size as also observed by Hoque et al. [6] and Pan et al[1]. Prolificity of Serrana goats increased up to parity order 5, maintaining the value of this ratio for parity orders 6 and 7, then decreasing for higher parity orders. These results are in accordance to those observed by Pang et al.[11]. Among the different parity groups the highest value for litter size were observed for goat’s age group 5[6]. The maximum and minimum litter size was found for parity order 5 and parity order 1, respectively.
This variation in prolificity according to parity is in line with the already described for this specie/breed and in accordance with the different rates of ovulation between nulliparous and multiparous Serrana goats observed by in previous study[12].
Pang et al. [11] denoted that high concentrations of progesterone observed in the plateau phase in high prolific goats relative to poor prolific ones were physiologically relevant, since they reflect the pooled output from many small corpora lutea. Also the falling progesterone levels in the plateau and early follicular phases, which are involved with inducing the pre-ovulatory surge of LH, were greater in high than poor prolific goats. This last observation suggested that this decreased might be related to high ovulation rate. These researchers also found that the pre-ovulatory follicles and corpora lutea were smaller but more prevalent in goats of high prolificity, resulting in different progesterone and oestradiol secretion patterns in comparison to poor prolific goats.
Several authors in studies conducted in sheep, agree that prolificity is essentially linked to FecB gene which plays a decisive role in ovulation[13-16]. However, in goats, the mechanism involving the prolificity remains unclear.
In our study, there was a higher average prolificity in Ribatejano and Jarmelista ecotypes in opposition to ecotypes Transmontano and Serra. For the Transmontano ecotype, prolificity remained relatively constant over the years, as suggested by the low variation on the confidence interval between 1999 and 2009, although it can be observed a slight increase of 0.05 foetuses per kidding during the three decades of the study.
Moreover the different ecotypes, that can suggest unequal genetic determination for prolificity, also the management conditions and feed availability at crucial times of the year (namely breeding season) may contribute to the observed differences. Sarmento et al. [5] found that the month of mating, the parityorder and age at first kidding are significant factors that influence the prolificity rate.
When prolificity was evaluated according to parturition month, it was observed a decrease in kidding from November reaching its minimum in March, after which it increased. These parturitions represent fertilizations in June, so less forage/feed availability and eventually the thermal stress may be associated with these results. Note that these observations were made independently of the occurrence of aborted foetuses or stillbirths. Females bred during the period of greater feed availability were more likely to have multiple kidding. However, the probability of multiple kidding are similar among females with 3 or more kidding, once these have reached physiological maturity[5].
In our study, it was observed a similar tendency in prolificity average, depending on the month of kidding, but with significantly higher values for the multiparous females.
Kidding season, and obviously mating season, affected litter size (P<0.001) similarly to the findings observed other authors[6,16].
The trenddenoted in nulliparous females, in our study, seems to be biologically correct, since younger females have not yet reached sexual maturity, anatomically and physiologically, displaying a higher incidence of single kidding.
In multiparous females there is a considerable increase in the probability of multiple kidding with increasing weight at mating and parity order. Lighter females and at parity order 1 have a lower probability of kidding more than one foetus although this could increase according to the weight of the female at mating [5], highlighting the influence of body condition at the time of fertilization.
It should be also highlighted that according to the experience of some of our goat producers, goats delivering from August to October/November (during the natural breeding season) from farms whose goats were not separate from bucks (as occurs mainly withthe Transmontano ecotype) are rapidly fertilized. Thus, there is a kidding interval shorter than usual, which can lead to an (apparent) low prolificity on the next parturition season. However, it is necessary to investigate this potential link between kidding interval and prolificity.
Prolificity is affected by environmental and climatic variations (month of the year), physiological maturity (parity order), body condition (weight) and the age of onset of reproductive life. The improvement on this reproductive parameter can be easily obtained by adjusting the dietary level and age at first breeding.
In conclusion, the data obtained from the studbook database of Serrana goat breed, from the Genpro program, it was concluded that: 1) The average prolificity in Serrana goats, estimated from 316,610 parturition records, was 1.47 with significant variations between ecotypes; 2) Similar to other breeds and species, prolificity increased up to 5th kidding, with a subsequent decrease; 3) there were observed significant monthly changes of average prolificity, probably caused by environmental factors, such as variations in feed availability, among others.
Acknowlegment
The authors would like to thank Manuel Silveira (Ruralbit) and Franscisco Pereira (ANCRAS) for the full access to the data of the pedigree records of Serrana Goats. This work was supported by the Foundation for Science and Technology, project reference UID/ CVT/00772/2013.
Conflict of interest statement
The authors declare that they have no conflict of interest.
[1] Pan S, Biswas CK, Majumdar D, SenguptaD, Patra A, Ghosh S, et al. Influence of age, body weight, parity and morphometric traits on litter size in prolific Black Bengal goats. J Appl Anim Res 2015; 43(1): 104-111.
[2] AnousMR, Rashed MA, MotaoaHR, SadekMH, Saad YM, Osman MA, et al. Identification of fecundity gene in Egyptian goats using genetic markers. I. Biochemical polymorphic markers. Egypt J Genet Cytol 2008; 37: 83-94.
[3] Haldar A, Pal P, Datta M, Paul R, Pal SK, Majumdar D, et al. Prolificacy and its relationship with age, body weight, parity, previous litter size and body linear type traits in meat-type goats. Asian-Australas J Anim Sci 2014; 27: 628-634.
[4] Palai TK, Bisoi PC, Maity A, Behera PC, Sahoo G, Polley S, et al. Prolificacy in Raighar goat is independent of FecB gene. Vet World 2013; 6(8): 479-481.
[5] Sarmento JLR, Pimenta-Filho EC, Abreu UGP, Ribeiro MN, Sousa JER. Litter size of crossbreed dairy goats in the northeastern semi-arid. R Bras Zootec 2010; 39(7): 1471-1476.
[6] Hoque MA, Amin MR, Baik DH. Genetic and non-genetic causes of variation in gestation length, litter size and litter weight in goats. Asian-Australas J Anim Sci 2002; 15(6): 772-776.
[7] Feng T, Geng CX, Lang XZ, Chu MX, Cao GL, Di R, et al. Polymorphisms of caprine GDF9 gene and their association with litter size in Jining Grey goats. Mol Biol Rep 2011; 38: 5189-5197.
[8] Rao S, Notter DR. Genetic analysis of litter size in Targhee, Suffolk and Polypay sheep. J Anim Sci 2000; 78: 2133-2120.
[9] Crepaldi P, Corti M, Cicogna M. Factors affecting milk production and prolificacy of Alpine goats in Lombardy (Italy). Small Rum Res 1999; 32: 83-88.
[10] Cary NC. Basic Analysis and Graphing. 2rd ed. SAS Institute Inc; 2012. JMP® 10.
[11] Pang XS, Wang ZY, Zhu TG, Yin DZ, Zhang YL, Meng L, et al. Concentrations of progesterone and estradiol in peripheral plasma during the estrous cycle and after ovariectomy in Huanghuai goats of high or poor prolificacy. Asian-Aust J Anim Sci 2010; 23(2): 188-196.
[12] Simões J, Baril G, Almeida JC, Azevedo J, Fontes P, Mascarenhas R. Time of ovulation in nulliparous and multiparous goats. Animal 2008; 2(5): 761-768.
[13] McNatty KP, Lun S, Heath DA, Ball K, Smith P, Hudson NL, et al. Differences in ovarian activity between Booroola x Merino ewes which were homozygous, heterozygous and non-carriers of a major gene influencing their ovulation rate. J Reprod Fertil 1986; 77: 193-205.
[14] McNatty KP, Juengel JL, Wilson T, Galloway SM, Davis GH. Genetic mutations influencing ovulation rate in sheep. Reprod Fertil Dev 2001; 13(7-8): 549-555.
[15] Xia Y, O’Shea T, Murison R, McFarlane JR. Concentrations of progesterone, follistatin, and follicle-stimulating hormone in peripheral plasma across the estrous cycle and pregnancy in Merino ewes that are homozygous or noncarriers of the Booroola gene. Biol Reprod 2003; 69: 1079-1084.
[16] Dickson UL, Torres GH, Becerril CP, Gonzalez FC, Osorio MA, Garcia OB. Comparison of Alpine and Nubian goats for some reproductive triats under dry tropical conditions. Small Rumin Res 2000; 36: 91-95.
ment heading
10.1016/j.apjr.2016.10.001
*Corresponding author: Sacoto Sandra, CECAV, Department of Animal Science, University of Trás-os-Montes e Alto Douro.
Tel: +351 259350416
E-mail: ssacoto@utad.pt
Foundation project: This work was supported by the Foundation for Science and Technology (FCT), project reference UID/CVT/00772/2013.
Local goat breeds
Reproductive efficiency
杂志排行

Asian Pacific Journal of Reproduction的其它文章
- Hormonal changes and spermatogenesis of male rat puppies born by mothers consuming soybean extract
- Factors affecting length of gestation in artificially inseminated Marwari Mares of India
- Nicotine effect toward the oocyte level of rats(Rattus novergicus)
- Effect of Thaumatococcus daniellii leaf rat-feed on potassium bromate induced testicular toxicity
- Substitution of egg yolk with different concentrations of soybean lecithin in trisbased extender during bulls' semen preservability
- The effect of freeze-drying media and storage temperature on ultrastructure and DNA of freeze-dried buffalo bull spermatozoa