羽毛还是鳞片
2017-01-06王旭日季燕南黑须球子
季 强, 王旭日, 季燕南, 黑须球子
(1.中国地质科学院地质研究所,北京100037; 2.中国地质环境监测院,北京100081; 3.中国地质大学(北京),北京100083)
羽毛还是鳞片
季 强1, 王旭日1, 季燕南2, 黑须球子3
(1.中国地质科学院地质研究所,北京100037; 2.中国地质环境监测院,北京100081; 3.中国地质大学(北京),北京100083)
简要介绍了羽毛的结构、类型、发育过程、起源及早期演化,讨论了“单根管状羽毛”的性质,指出这种管状结构不是羽毛,而是高度变形的特化鳞片。尽管羽毛与爬行动物的鳞片同源,而且羽毛是由恐龙身上的鳞片演变而来,但关键在于小囊领的分异和羽枝脊的出现与否。也就是说,在形态结构上,如果表皮的角质化衍生物出现了羽根和羽小枝,就应该将其称为羽毛,未发生这种分异的结构应当归于高度变形的特化鳞片。同时还讨论了德国法兰克福圣肯堡自然历史博物馆收藏的带纤维状结构的鹦鹉嘴龙和俄罗斯西伯利亚发现的植食性恐龙(Kulindadromeuszabaikalicus),认为二者发育的纤维状结构或羽毛状结构不是羽毛,而是高度变形的特化鳞片,并不存在鳞片与羽毛共生的现象。就目前的材料而言,羽毛仅限发育于兽脚类恐龙中,并不是所有的恐龙均发育羽毛。
鳞片;羽毛;起源;早期演化;发育过程
0 引 言
长期以来,一直认为羽毛是鸟类特有的结构(皮肤衍生物),在动物世界中只有鸟类才发育羽毛。不过,自1996年中国辽西发现了世界上第一只长羽毛的恐龙——中华龙鸟(Sinosauropteryxsinensis)以后,才开始认识到羽毛并非鸟类专有,恐龙也同样可以发育羽毛,而且羽毛与爬行动物的鳞片同源,是由恐龙的鳞片逐渐演变而来的。
简要介绍了羽毛的结构、类型、发育过程、起源及早期演化,并尝试讨论了羽毛的定义及其性质(周忠和等,1992;侯连海等,1993;季强等,1996,1997,2002a,2002b,2004; Ji et al., 1998, 1999, 2001, 2003, 2005, 2009; Xu et al., 1999, 2000, 2003; Zhou et al., 1999, 2002; Norell et al., 2002; Zhang et al., 2006)。
1 羽毛的结构
羽毛的结构或简或繁,变化较大。雏绒羽的结构比较简单,仅由羽根、羽枝,或由羽根、羽枝和羽小枝组成。正羽的结构较为复杂,一般由羽根、羽干和羽片组成。羽片又由羽枝、羽小枝、羽小钩、羽小槽等组成(图1)。

图1 羽毛结构示意图(据季强,2016)Fig.1 Sketch showing the structure of feathers(after Ji, 2016)
2 羽毛的发育过程
研究表明,羽毛的发育经历了一个十分复杂的过程。羽毛一开始产生于基板,基板向上凸起形成圆锥形羽芽;羽芽周边的细胞不断增生并向皮内凹陷,形成小囊;在小囊的底部,小囊领的角蛋白细胞不断增生,将老细胞向外挤出形成空心管状。表皮层的最外层形成羽鞘,表皮层的内层被隔开形成羽枝脊,然后发育为羽枝。随着羽毛的发育,羽毛从羽鞘中发散出来,正羽的羽枝脊围绕小囊领呈螺旋式生长,联合形成羽干脊,其上再发育羽枝脊。绒羽的羽枝脊生长不呈螺旋式,只在羽毛的底部形成短而简单的羽干。当羽毛发育到成熟阶段,小囊领就变成羽根。
羽毛的发育过程见图2。

图2 羽毛的发育过程(据季强,2016)Fig.2 Developmental process of feathers(after Ji, 2016)
3 羽毛的类型
羽毛的形态结构多种多样,差异很大,功能各不相同,大致可分为正羽(pluma/contour feather)、雏绒羽(natal down/nestling down)、绒羽(plumule/down feather)、毛羽(filoplume/hair-like feather)、须羽(bristle)、粉(powder feather)、半绒羽(semiplume)等。
3.1 雏绒羽
雏绒羽为幼体出壳后覆盖体表的丝状羽毛,一般仅由羽根、羽枝和羽小枝构成,羽小枝完全不具有羽小钩,如鸡、雁、鸮等鸟类的雏绒羽特别发育(图3)。

图3 家鸡的雏绒羽(据郑光美,1995)Fig.3 Natal down of domestic chickens(after Zheng, 1995)
3.2 绒羽
绒羽是指成体正羽下面密集生长的丝状羽毛,一般由羽根、羽枝和羽小枝组成,可发育正羽羽小枝和副羽羽小枝,不具羽小钩或很稀少,羽干短小或缺失,羽枝成簇地从羽根顶部发散开,构成有效的隔热层(图4)。

图4 家鸡的绒羽(据郑光美,1995)Fig.4 Plumule of domestic chickens(after Zheng, 1995)
3.3 正羽
正羽是最常见、最复杂的羽毛,一般覆盖于体表,形成严密的保护层。飞羽和尾羽是特化的正羽,主要与飞行功能有关。正羽由羽根(calamus)、羽干(rachis)和两侧羽片(vane)构成,羽片又由羽枝(barb)、羽小枝(barbule)、羽小钩(hooklet)、羽小槽(groove)等构成。羽根和羽干一起构成羽轴(shaft)(图5)。
3.4 半绒羽
半绒羽一般发育于正羽之下,沿腹侧、颈、背中部及飞羽和尾羽基部分布,其结构介于正羽与绒羽之间,由羽根、羽干、羽枝和羽小枝组成,但没有羽小钩和凸缘,因而不能形成羽片。
3.5 毛羽
毛羽散布于正羽与绒羽之间,羽干细长,顶端发育少许羽枝和羽小枝。毛羽羽根的滤泡附近发育丰富的触觉神经末梢,能感知正羽的姿态,从而控制羽毛的运动(图6)。

图5 正羽(飞行羽毛)(据季强,2016)Fig.5 Pluma (flight feather)(after Ji, 2016)

图6 毛羽结构示意图(据郑光美,1995)Fig6 Sketch showing the structure of hair-like feathers(after Zheng, 1995)
3.6 须羽
须羽是一种特化的正羽,一般发育于口裂两侧、鼻孔周围等头部特定区域,羽轴硬而纤长,形如须毛,基部发育少许羽枝或完全没有羽枝。须羽的根部周围皮肤内发育丰富的感觉小囊,故具有触觉功能(图7)。

图7 须羽结构示意图(据郑光美,1995)Fig.7 Sketch showing the structure of bristles(after Zheng, 1995)
4 羽毛的起源与早期演化
羽毛与爬行动物的鳞片同源,是由恐龙身上的鳞片演变而来的,其早期演化大致可分为5个阶段(图8)。 第一阶段:小囊领发生阶段,开始出现拉长的中空管状结构,一般多见于异齿龙类、角鼻龙类、异特龙类等。
第二阶段:小囊领发生分异的阶段,形成隔开的羽枝脊,从羽根上发育出未分叉的羽枝簇。季强等(1996)将这种羽毛称为原始羽毛(protofeather/profeather),多见于美颌龙类恐龙。
第三阶段:羽小枝板发生阶段,存在2个不同的发展方向:一个方向是羽枝脊围绕小囊领呈螺旋式生长,联合形成羽轴脊,羽干拉长,羽干两侧先发育未分叉的羽枝,然后再发展成具羽小枝的羽枝,形成具开放羽片的羽毛(半绒羽);另一个方向是原始羽毛先发展成羽干短小,端部发育出具羽枝和羽小枝的羽毛(绒羽),然后再发展为具开放羽片的羽毛(半绒羽),这一阶段的羽毛多见于似鸟龙类、镰刀龙类、暴龙类等恐龙。
第四阶段:羽小枝板的分异阶段,羽小枝发育了羽小钩和羽小槽,二者紧密相连,形成具有封闭羽片的羽毛(正羽),多见于窃蛋龙类、伤齿龙类、驰龙类等恐龙。
第五阶段:羽枝脊不对称发育阶段,形成具有不对称封闭羽片的正羽(飞行羽毛),多见于窃蛋龙类、伤齿龙类、驰龙类等恐龙和各种鸟类。
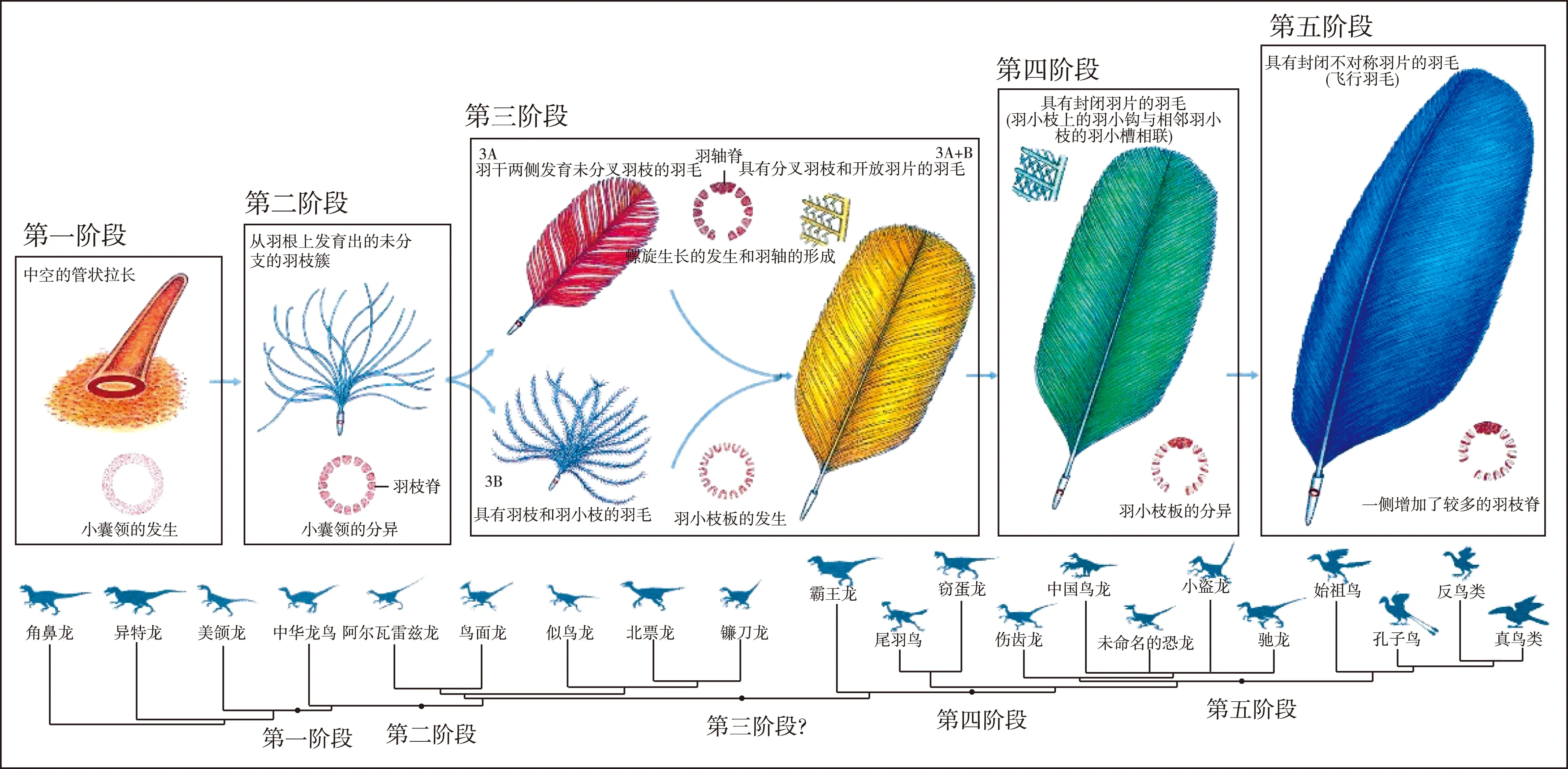
图8 羽毛起源与早期演化示意图(据季强,2016)Fig.8 Sketch showing the origin and early evolution of feathers(after Ji, 2016)
5 讨 论
对于第一阶段的拉长的管状结构,有些学者认为这是最简单的羽毛,另有一些学者则认为是高度变形的特化鳞片。这一问题涉及到羽毛的定义,也就是说,羽毛与鳞片的本质区别是什么。
在过去的几十年中,人们对于羽毛的定义、起源和早期演化问题有着广泛的讨论。季强等(1996)首次提出中华龙鸟身上发育了纤维状的原始羽毛(protofeather/profeather)(图9)。但当时多数学者均不认为这种纤维状的皮肤衍生物是原始羽毛,而将其称为丝状结构(filamentous structure);有些学者甚至认为这种结构可与一些具有滑翔能力的爬行动物身上发育的拉长的管状结构相比较。

图9 中华龙鸟发育的纤维状原始羽毛(据季强,2016)Fig.9 Fibrous protofeathers covered on the body of Sinosauropteryx(after Ji, 2016)
Daedalosaurus是一种小型的具有滑翔能力的爬行动物,产于马达加斯加晚二叠世地层中。这种蜥蜴形的动物具有1条长约40 cm的尾巴,身体两侧各发育21条很长的肋条状结构,支撑皮膜进行滑翔,两“翼”展开可达33 cm。Daedalosaurus发育的这种结构使人联想到现今生活在东南亚热带雨林中的一种飞龙——Draco,其也具有相似的结构,可以滑翔近60 m(图10)。

图10 几种会滑翔的爬行动物晚二叠世的Daedalosaurus,Coelurosauravus;晚三叠世的Icarosaurus,Kuehneosaurus (据Cranfield, 2001)Fig.10 Some gliding reptiles: Daedalosaurus and Coelurosauravus of Late Permian; Icarosaurus and Kuehneosaurus of Late Triassic(after Cranfield, 2001)
Sharovipteryx也是一种细长的具有滑翔能力的爬行动物,产于吉尔吉斯斯坦晚三叠世地层中,身长大约25 cm,前肢短小,后肢极长,三角形的皮膜从后肢伸展到尾巴的前部(腿翼),具有一定的滑翔能力,但滑翔距离不会太远(图11)。

图11 会滑翔的爬行动物Sharovipteryx形态结构示意图(据Cranfield, 2001)Fig.11 Sketch showing the morphology and structure of a gliding reptile Sharovipteryx(after Cranfield, 2001)
Longisquama原指一种具有“长鳞片”的小型爬行动物,产于吉尔吉斯斯坦晚三叠世地层中,身长约10~12 cm,背部两侧各发育一列极长的附属结构,超过了身体的长度,明显具有较强的滑翔能力,但不能利用其背部的附属结构进行煽拍(图12)。

图12 会滑翔的爬行动物Longisquama形态结构示意图(据Cranfield, 2001)Fig.12 Sketch showing the morphology and structure of a gliding reptile Longisquama(after Cranfield, 2001)
从以上介绍可以看出,这些会滑翔的爬行动物发育的翼状结构与中华龙鸟发育的原始羽毛完全是两种概念。当时认定中华龙鸟发育的纤维状皮肤衍生物不是羽毛,其原因看来是多方面的,但最主要的原因是那些学者坚持鸟类“非恐龙起源”的学术思想,认为恐龙根本不可能发育羽毛。
此后,又有一些报道在中国辽西早白垩世义县组中发现1枚长“羽毛”的鹦鹉嘴龙化石(Psittacosaurussp.),在俄罗斯西伯利亚侏罗纪地层中发现1只具有鳞片和丝状结构(皮肤衍生物)的鸟臀类恐龙——Kulindadromeuszabaikalicus(Godefroit et al., 2014)。
中国辽西发现的长“羽毛”的鹦鹉嘴龙标本现收藏在德国法兰克福圣肯堡自然历史博物馆,迄今仍没有见到有关这件化石的正式研究论文。从该化石的图片来看,标本完整精美,呈腹-背压覆状态(腹面朝上),身体内部和四周保存了黑色有机残留物;所谓的“羽毛”似乎保存在尾部的下方(背面),呈长长的纤维状(或为管状),尾部的腹面没有见到这种结构的任何痕迹(图13)。

图13 保存纤维状结构的鹦鹉嘴龙化石(据季强,2016)Fig.13 A specimen of Psittacosaurus with filamentous structures(after Ji, 2016)
发现于俄罗斯西伯利亚侏罗纪的草食性鸟臀类恐龙(Kulindadromeuszabaikalicus)体长约1.5 m,前肢较短,后肢粗长,腰带短小,尾巴很长;较小的鳞片分布在后肢的远端,较大的鳞片分布在尾部,单根的纤维状结构分布在头部和胸部,较为复杂的纤维状结构分布在肱骨、股骨和胫骨周边(图14)。Godefroit等(2014)认为,较为复杂的纤维状(羽毛状)结构与鳞片共生,这种现象在整个恐龙家族中普遍存在,并且进一步断言,所有的恐龙,包括最早期的恐龙,都会发育羽毛。

图14 Kulindadromeus zabaikalicus的生态复原图(据Godefroit et al., 2014)Fig.14 Ecological restoration image of Kulindadromeus zabaikalicus(after Godefroit et al., 2014)
对于中国辽西发现的具纤维状结构的鹦鹉嘴龙而言,首先要确定这种纤维状结构与鹦鹉嘴龙的关系,也就是说,它们是否真的是鹦鹉嘴龙身上发育出来的结构。如果二者没有任何关系,那么也就没有必要讨论鹦鹉嘴龙是否长有纤维状皮肤衍生物或“羽毛”的问题;如果这种纤维状结构确实是鹦鹉嘴龙发育的,那么就需要仔细研究它们的形态结构,然后再讨论它们是否是羽毛的问题。2009年,笔者曾在德国法兰克福圣肯堡自然历史博物馆观察了该馆收藏的那件鹦鹉嘴龙标本,认为:(1) 难以确定这些纤维状结构是否属于鹦鹉嘴龙自身发育的结构;(2) 这些结构为拉长的空心管状,未发现任何与羽毛相关的结构;(3) 即使这些结构是皮肤衍生物,它们应该是拉长的变形的鳞片,而不是羽毛(图15)。

图15 鹦鹉嘴龙尾部的纤维状结构(据季强,2016)Fig.15 Filamentous structure at the tail of Psittacosaurus(after Ji, 2016)
就俄罗斯西伯利亚发现的Kulindadromeuszabaikalicus而言,其身上发育有鳞片是确凿无疑的,但单根纤维状结构和较为复杂的纤维状结构性质值得商榷。首先,单根纤维状结构与复杂纤维状结构最明显的区别在于是否发育了基板(basal plates),二者抑或是同一种结构,只不过单根羽毛没有保存基板;其次,所谓的基板究竟是一种什么性质的结构?是羽根?是鳞片?还是其他结构?Godefroit等(2014)仅仅描述了形态特征,并未就其性质作出说明。从他们提供的图片来看,与其说这些结构是羽毛,还不如说它们是一种高度变形的特化鳞片。Godefroit等(2014)在文献中均使用纤维状结构(filamentous structures)或羽毛状结构(feather-like structures),但在标题和结论中使用了羽毛(feathers)一词,并断言最早期的恐龙都会发育羽毛(图16、图17)。这十分令人费解,不知该结论的依据是什么。

图16 Kulindadromeus zabaikalicus的表皮鳞片和羽毛状结构(据Godefroit et al., 2014)(a) 胫骨和踝骨周围的鳞片;(b) 尾部近端发育的双列鳞片;(c) 尾部鳞片的局部放大;(d) 部分头骨;(e) 保存单根纤维状结构的部位;(f) 单根纤维状结构的局部放大;(g) 左胸部;(h) 保存单根纤维状结构的部位;(i) 单根纤维状结构的局部放大Fig.16 Photos showing epidermal scales and featherlike structure of Kulindadromeus zabaikalicus(after Godefroit et al., 2014)(a) scales around the tibia and ankle; (b) diallel scales at the proximal tail ; (c) local amplification of tail scales; (d) partial skull; (e) position with preserved single filamentous structure; (f) local amplification of single filamentous structure; (g) left chest; (h) position with preserved single filamentous structure; (i) local amplification of single filamentous structure.

图17 Kulindadromeus zabaikalicus的羽毛状结构(据Godefroit et al., 2014)(a) 肱骨、桡骨和尺骨;(b) 复杂纤维状结构;(c) 复杂纤维状结构示意图;(d) 肱骨周围保存的复杂纤维状结构;(e) 复杂纤维状结构的局部放大;(f) 复杂纤维状结构示意图;(g) 胫骨近端周围保存的条带状结构;(h) 保存条带状结构的部位;(i) 条带状结构示意图;(j) 条带状结构的局部放大Fig.17 Featherlike structure of Kulindadromeus zabaikalicus(after Godefroit et al., 2014)(a) humerus, radius and ulna; (b) complex filamentous structure; (c) sketch showing complex filamentous structure; (d) complex filamentous structure preserved around humerus; (e) local amplification of complex filamentous structure; (f) sketch showing complex filamentous structure; (g) strip structure preserved around the proximal tibia; (h) position with preserved strip structure; (i) sketch showing strip structure; (j) local amplification of strip structure
如果这些纤维状结构是高度变形的特化鳞片的话,那么正常的鳞片与特化的鳞片共存就是一个十分平常的现象,并不能将其作为依据,并断言所有的恐龙都长有羽毛。
Xu等(2010)讨论了早期羽毛的发育过程,将早期羽毛分为9种类型(图18),并且列出了不同种类的恐龙发育的羽毛类型。异齿龙类和鹦鹉嘴龙类主要发育类型1和类型2的“羽毛”;暴龙类、美颌龙类和镰刀龙类主要发育类型2—5的羽毛,是否发育类型1的“羽毛”尚不能确定;窃蛋龙类主要发育3、5、7、8类型的羽毛;伤齿龙类主要发育3、5、8类型的羽毛;驰龙类主要发育3—5、8—9类型的羽毛;擅攀鸟龙类主要发育6—7类型的羽毛;始祖鸟主要发育3、5、8、9类型的羽毛;孔子鸟类主要发育3—5、7—9类型的羽毛;反鸟类主要发育3—5、7—9类型的羽毛(图19)。

图18 早期羽毛的类型(据Xu et al., 2010)Fig.18 Types of early feathers(after Xu et al., 2010)

图19 不同种类的恐龙发育不同类型的羽毛(据Xu et al., 2010)Fig.19 Different types of feathers of different types of dinosaurs(after Xu et al., 2010)
6 结 论
(1) Xu等(2010)的研究成果对于人们了解羽毛的早期演化十分重要,但类型1和类型2是否可称为羽毛值得商榷,因为这些拉长的空心管状结构并没有显示任何羽毛的特征。
(2) 尽管羽毛与爬行动物的鳞片同源,而且羽毛是由恐龙身上的鳞片演变而来的,但关键在于小囊领的分异和羽枝脊的出现与否。也就是说,在形态结构上,如果表皮的角质化衍生物出现了羽根和羽小枝,就应该将其称为羽毛,未发生这种分异的结构应当归于高度变形的特化鳞片。
(3) 笔者愚见,就目前的研究材料和研究程度而言,与其将类型1和2归于羽毛,不如将其归于高度变形的特化鳞片,因此羽毛的发育仍仅限于兽脚类恐龙,而不是所有恐龙均发育羽毛(Ostrom, 1974, 1976, 1979; Padian, 1982, 1985, 2003; Padian et al., 2001, 2005; Mayr et al., 2005; Xu et al., 2010; Ji et al., 2015)。
侯连海,张江永,1993. 辽宁早白垩世早期一鸟化石[J]. 古脊椎动物学报,31(3): 217-224.
季强,姬书安,1996. 中国最早鸟类化石的发现及鸟类的起源[J]. 中国地质,23(10):30-33.季强,姬书安.1997. 原始祖鸟(Protarchaeopteryxgen. nov.):中国的始祖鸟类化石[J]. 中国地质,24(3):38-41.季强,姬书安,尤海鲁,等,2002a. 中国首次发现真正会飞的“恐龙”:中华神州鸟(新属、新种)[J]. 地质通报,21(7):363-369.
季强,姬书安,张鸿斌,等,2002b. 辽宁北票首次发现初鸟类化石:东方吉祥鸟(Jixiangornisorentalisgen. et sp. nov.)[J]. 南京大学学报(自然科学版),38(6):723-736.
季强等, 2004. 中国辽西中生代热河生物群[M]. 北京,地质出版社.
季强, 2016. 腾飞之龙[M]. 北京,地质出版社.
周忠和,金帆,张红永, 1992. 辽宁中生代早期鸟类化石的初步研究[J]. 科学通报,37(5):435-437.
郑光美, 1995. 鸟类学[M]. 北京:北京师范大学出版社.FOTH C, TISCHLINGER H, RAUHUT O W M, 2014. New specimen ofArchaeopteryxprovides insights into the evolution of pennaceous feathers [J]. Nature, 511(7507): 79-82.GODEFROIT P, SINITSA S M, DHOUAILLY D, et al., 2014. A Jurassic ornithischian dinosaur from Siberia with both feathers and scales [J]. Science, 345(6195): 451-455.
JI Q, CURRIE P J, NORELL M A, et al., 1998. Two feathered dinosaurs from northeastern China [J]. Nature, 393(6687): 753-761.
JI Q, CHIAPPE L M, JI S A, 1999. A new Late Mesozoic confuciusornithid bird from China [J]. Jour. Vert. Palaeont., 19 (1): 1-7.
JI Q, NORELL M A, GAO K Q, et al., 2001. The distribution of integumentary structures in a feathered dinosaur [J]. Nature, 410(6832): 1084-1088.
JI Q, JI S A, YOU H L, et al., 2003. Early Cretaceous Avilian bird, Shenzhouraptor sinensis from western Liaoning, China [J]. Acta Geological Sinica, 77 (1): 21-27.
JI Q, JI S A, LÜ J C, et al., 2005. First Avilian bird from China (Jinfengopteryxelegansgen. et sp. nov.) [J]. Geological Bulletin of China, 24 (3): 197-210.
JI Q, JI S A, 2009. First large tyranosauroid theropod from the Early Cretaceous Jehol Biota of Northeastern China [J]. Geological Bulletin of China, 28 (10): 1369-1374.JI Y N, WANG X R, KUROSU M, et al., 2015. A debate between the ″arboreal″ and ″cursorial″ hypotheses of the origin of bird flight [J]. Journal of Geology, 39 (2): 201-206.MAYR G, POHL B, PETERS D, 2005. A well-preservedArchaeopteryxspecimen with theropod features [J]. Science, 310(5753): 1483-1486.NORELL M A, JI Q, GAO K Q, et al., 2002. ″Modern″ feathers on a non-avian dinosaur [J]. Nature, 416(6876): 36-37. OSTROM J, 1974.Archaeopteryxand the origin of flight [J]. Quart. Rev. Biol., 49(1): 27-47.
OSTROM J,1976.Archaeopteryxand the origin of birds [J]. Biol. Jour. Linnean Soc., 8(2): 91-182.
OSTROM J, 1979. Bird flight: how did it begin? [J]. American Scientist, 67(1): 46-56.
PADIAN K, 1982. Running, leaping, lifting off: fight evoloved from the ground up, not the trees down [J]. The Sciences, 22: 10-15.
PADIAN K, 1985.The origin and aerodynamics of flight in extinct vertebrates [J]. Palaeontology, 28 (3): 413-433.
PADIAN K, JI Q, JI S, 2001.Feathered dinosaurs and the origin of flight [M]//CARPENTER K,CURRIE P J, TANKE D H. Mesozoic Vertebrate Life: New Research inspired by the Palaeontology of Philip J. Currie. Bloomington and Indianapolis: Indiana Univ. Press, 117-135.
PADIAN K, 2003. Four winged dinosaurs, bird precursors, or neither? [J]. Bio. Science, 53 (5): 450-452.
PADIAN K,DIAL K P, 2005. Origin of flight: could four-winged dinosaurs fly? [J]. Nature, 438(7066): 3-4.
XU X, TANG Z L, WANG X L, 1999. A therizinosaurid dinosaur with integumentary structures from China [J]. Nature, 399(6734): 350-354.XU X, ZHOU Z H, WANG X L, 2000. The smallest known non-avian theropod dinosaur [J]. Nature, 408(6813): 705-708.XU X, ZHOU Z H, WANG X L, et al., 2003. Four-winged dinosaurs from China [J]. Nature, 421(6921): 335-340.
XU X, ZHENG X T, YOU H L, 2010. Exceptional dinosaur fossils show ontogenetic development of early feathers [J]. Nature, 464 (29): 1338-1341.
ZHOU Z H,MARTIN L D, 1999. Feathered dinosaur or bird? :a new look at the hand ofArchaeopteryx[J]. Smithsonian Contributions to Paleobiology, 89: 289-293.
ZHOU Z H, ZHANG F C, 2002. A long-tailed, seed-eating bird from the Early Cretaceous of China [J].Nature,418(6896): 405-409.
ZHANG F C, ZHOU Z H, DYKE D, 2006. Feathers and ′feather-like′ integumentary structures in Liaoning birds and dinosaurs [J]. Geol. Jour., 41(3/4): 395-404.
Feathers or highly modified scales?
JI Qiang1, WANG Xuri1, JI Yannan2, KUROSU Mariko3
(1. Institute of Geology, Chinese Academy of Geological Sciences, Beijing 100037, China; 2. China Geological Environmental Monitoring Institute, Beijing 100081, China; 3. China University of Geosciences (Beijing), Beijing 100083, China)
This paper briefly introduces the structures, morphotypes, developmental processes, origin and early evolution of feathers, and discusses the characters of "single elongate filamentous feathers". It is considered that those elongate filamentous structures are highly modified scales, rather than feathers. As we know, feathers are homologous with scales of reptiles, and they are derived from the scales of dinosaurs. However, feathers can be distinguished from scales biologically by the differentiation of follicle collar and the development of barb ridges, and morphologically by the development of calamus and barbs. In addition, this paper also discusses a psittacosaurid dinosaur with elongate filamentous structures housed by Senckenbergische Naturforschende Gesellschaft, Frankfurt of Germany and a basal neornithischian dinosaur (Kulindadromeuszabaikalicus) from the Jurassic of Siberia with small scales and feather-like structures. It is inferred that both of them do develop highly modified scales, rather than feathers. Maybe feather-like structures can coexist with scales and potentially be widespread among the entire dinosaur clade, but they are not feathers. It is an unwarranted imagination that feathers may have been present in the earliest dinosaurs.
scale; feather; origin; early evolution; development process
10.3969/j.issn.1674-3636.2016.04.535
2016-04-22;
2016-05-05;编辑:詹庚申
国家重点基础研究发展计划(“973”计划)项目(2012CB822004-2),中国地质调查局项目(12120114026801)
季强(1951— ),男,研究员,博士,博士生导师,长期从事理论古生物学和地层学研究,E-mail: jirod@cags.ac.cn
Q915.2+3
A
1674-3636(2016)04-0535-10
