冬虫夏草菌丝状菌源增殖培养的研究
2016-12-27贺宗毅张德利毛先兵陈仕江
贺宗毅,李 黎,张德利,李 卿,毛先兵,陈仕江*
(1.重庆市中药研究院,重庆 400065;2.重庆市中药资源学重点实验室,重庆 400065;3.中国中医科学院,中药资源中心重庆分中心,重庆 400065;4.甘孜州康定贡嘎中华虫草产业有限责任公司,四川甘孜 626000;5.西南药业股份有限公司,重庆 400000)

冬虫夏草菌丝状菌源增殖培养的研究
贺宗毅1,2,3,4,李 黎1,2,3,4,张德利1,2,3,4,李 卿1,毛先兵5,陈仕江1,2,3,4*
(1.重庆市中药研究院,重庆 400065;2.重庆市中药资源学重点实验室,重庆 400065;3.中国中医科学院,中药资源中心重庆分中心,重庆 400065;4.甘孜州康定贡嘎中华虫草产业有限责任公司,四川甘孜 626000;5.西南药业股份有限公司,重庆 400000)
冬虫夏草菌是珍稀濒危名贵中药材冬虫夏草的无性型菌种,是侵染蝠蛾幼虫的唯一菌种,其侵染后形成名贵中药材冬虫夏草。冬虫夏草菌在不同营养及条件下生长形态不同,主要有丝状体和菌球体两种生长形态。冬虫夏草菌丝状菌体能够侵染蝠蛾幼虫,有可能作为一种新的接种体被广泛应用;但丝状菌体对营养要求苛刻、生长缓慢、菌丝体得率低的特点,阻碍了冬虫夏草菌丝状菌体作为侵染蝠蛾幼虫的接种体的开发及应用,从而阻碍了冬虫夏草人工培殖产业化的进程。为了提高冬虫夏草菌丝状菌体的生物表达量,本文对营养及培养条件进行了优化研究,得出了最佳条件为:葡萄糖质量浓度40 g/L、酵母粉质量浓度65 g/L、培养温度17℃、MgSO4质量浓度3 g/L、KH2PO4质量浓度1.5 g/L、培养时间12 d,按最优培养条件,冬虫夏草菌丝状菌体干重得率15.67 g/L,为下一步新的接种体的制备提供菌源保障。
冬虫夏草菌;丝状菌体;虫草蝠蛾幼虫;培养
冬虫夏草Ophiocordycepssinensis(Berk.)(Sungetal.,2007)系我国特产珍稀名贵中药材,是冬虫夏草菌O.sinensis侵入蝠蛾属Hepialus幼虫体内后形成的一种菌物药,是冬虫夏草菌丝体生长发育的产物。冬虫夏草的用药历史始载于《本草纲目拾遗》,称其有“补肺益肾、止血化痰”之功效。近代药理和临床研究表明,冬虫夏草与冬虫夏草菌丝体产物药理作用相近,均具有免疫调节(Wuetal.,2006)、抗氧化(Dong&Yao,2008)、调节高脂血症(Zhangetal.,2006)、调节血糖(Kohetal.,2003)、抗肿瘤(Wuetal.,2005)、延缓衰老(Jietal.,2009)等功效。
目前蝠蛾幼虫的接种体来源有冬虫夏草菌的分生孢子和冬虫夏草菌丝体,分生孢子是接种体的主要来源,但在冬虫夏草人工培殖的产业化过程中发现冬虫夏草菌产分生孢子的能力在逐渐下降,甚至有些批次的菌体不产生分生孢子,阻碍了冬虫夏草人工培殖的产业化,在蝠蛾幼虫接种体领域寻求新的突破将会推动冬虫夏草人工培殖产业化的进程。已有研究表明除冬虫夏草菌分生孢子具有侵染力外,冬虫夏草菌丝体亦具有侵染力(俞永信,2004)。长期以来冬虫夏草菌丝状菌体作为侵染体的研究没有得到重视,致使可用于接种体制备的丝状菌体培养的报道较少。
冬虫夏草菌在不同营养条件下生长形态不同,菌球体更易获得,丝状菌体的形成对营养要求更为苛刻,因此是否能够大量培养出丝状菌体对推动蝠蛾幼虫接种体的创制具有重要意义。本文以丝状菌体生物量和腺苷为考察指标,研究丝状菌体增殖培养条件,为下一步蝠蛾幼虫接种体的制备提供菌源保障。
1 材料与方法
1.1 材料与仪器
供试菌株:冬虫夏草Ophiocordycepssinensis菌种,保存于重庆市中药研究院。
仪器设备:净化工作台SW-CJ-1D(苏州净化设备有限公司);生化培养SPX-150B-Z(上海博迅实业有限公司医疗设备厂);分析天平AUY120(岛津国际贸易公司);Aailent 1260(安捷伦科技有限公司)。
1.2 实验方法
1.2.1 斜面种子培养
于母种试管中切取出黄豆粒大小的菌丝块,接种于斜面中部。16℃恒温培养30 d。菌种用斜面培养基保存和培养。斜面培养基为PPDA培养基。
1.2.2 液体种子培养
将培养好的斜面菌种切取蚕豆大小,接于液体培养基中,250 mL三角瓶装液量100 mL,16℃,150 r/min,摇床培养12 d 。
液体种子基础培养基为:葡萄糖15 g,酵母粉5 g,硫酸镁0.5 g,磷酸二氢钾0.5 g,磷酸氢二钾0.5 g,水1000 mL。
1.2.3 灭菌及接种
灭菌:将配制好的液体培养基分装,500 mL三角瓶装液量100 mL,用棉塞和牛皮纸封口后,放入高压锅内,在121℃条件下灭菌25 min。
接种:在洁净工作台上进行接种,选用纯化的试管菌种,挑取黄豆粒大小的菌丝接入三角瓶液体中,每瓶接一支固体菌种,接菌时,室内要严格消毒,防止杂菌进入培养液而影响虫草菌丝生长。
1.2.4 摇瓶发酵培养
按10%的接种量取液体种子进行接种,250 mL三角瓶装液量100 mL,16℃,150 r/min,摇床培养12 d 。
1.2.5 生物量计算
将培养液置于有孔径为0.1 cm的筛板上,反复用蒸馏水冲洗筛板至丝状菌体全部通过筛板,收集滤液,离心(4000 r/min,15 min),用蒸馏水冲洗菌丝滤饼2次,再离心,菌丝60℃电热恒温干燥箱中烘干至恒重,称重得菌丝体干重生物量。
1.2.6 Plackett-Burman筛选实验及正交实验设计
Plackett-Burman筛选实验:为了筛选出影响冬虫夏草菌丝状菌体生长的显著因子,本文将在单因素实验的基础上设计Plackett-Burman筛选实验,每个因子取高(+1),低(-1)2个水平。以菌丝体干重生物量为响应值Y。参数和水平设计见表1,各因素的效应评价及系数估计见表3。P<0.05说明该因素对菌丝体生物量的影响是显著的,可对该因素作进一步的考察。实验结果见表2。
正交实验:将对Plackett-Burman筛选的因素指标进行正交实验考察。各因素选取水平情况、试验结果及分析见表4和表5。
1.2.7 冬虫夏草菌丝体腺苷含量的测定
将烘干至恒重的菌丝体粉碎并过三号药筛,得冬虫夏菌菌丝体粉末,按2015版《中国药典》附录VI D方法测定。
1.2.8 数据处理
实验数据采用SPPS 19.0软件进行方差分析,采用Origin 8.5进行软件作图。
2 结果与分析
2.1 单因素实验
2.1.1 碳源的确定
分别考察葡萄糖、蔗糖、半乳糖、麦芽糖、乳糖和可溶性淀粉等6种碳水化合物对冬虫夏草菌丝状菌体生长的影响,结果见图1。碳源是冬虫夏草菌能量代谢的来源,不同种类的碳源对其冬虫夏草菌丝体得率影响不尽相同。由图1可知冬虫夏草菌利用碳源的情况各不相同,经SPSS统计学分析表明以葡萄糖为碳源时冬虫夏草菌丝体产量显著高于其它碳源(P<0.05),而以蔗糖、麦芽糖、可溶性淀粉为碳源时,菌体产量无显著性差异(P>0.05)。故选葡萄糖作为冬虫夏草菌生长的碳源。

图1 碳源种类对冬虫夏草菌生长的影响Fig.1 The effect of carbon sources on Ophiocordyceps sinensis cell growth 注:0,空白;1,葡萄糖;2,蔗糖;3,半乳糖;4,麦芽糖;5,乳糖;6,可溶性淀粉。Note:0,Control;1,Glucose;2,Sucrose;3,Galactose;4,Malt sugar;5,Lactose;6,Starch.
2.1.2 氮源的确定
分别考察有机氮源蛋白胨、酵母粉、牛肉膏和无机氮源尿素、硫酸铵、磷酸二氢铵对菌体生长的影响。结果如图2。氮源是冬虫夏草菌生长过程中不可缺少的元素,研究了上述氮源,从图2中可知冬虫夏草菌能较好地利用有机氮源,几乎不利用无机氮源,这与冬虫夏草菌侵染到蝠蛾幼虫体内后利用昆虫体为营养进行生长的营养模式一致。有机氮源所产菌丝体量均显著优于无机氮源所产菌丝体的量(P<0.05),以酵母粉为氮源时的菌丝体产量显著优于以蛋白胨、牛肉膏为氮源时的产量(P<0.05),故选择酵母粉作为冬虫夏草菌培养的氮源。

图2 不同氮源对冬虫夏草菌生长的影响Fig.2 The effect of nitrogen sources on Ophiocordyceps sinensis cell growth 注:1,空白;2,蛋白;3,酵母粉;4,牛肉膏;5,尿素;6,硫酸铵;7,磷酸二氢铵。Note:1,Control;2,Peptone;3,Yeast powder;4,Beef extract;5,Urea;6,Ammonium sulfate;7,Ammonium dihydrogen phosphate.
2.1.3 碳源质量浓度对冬虫夏草菌体生长的影响
碳源是冬虫夏草菌能量代谢的来源。在培养基中分别添加不同质量浓度的葡萄糖,其它条件一致,培养完成后研究其对腺苷含量及菌丝体生物量的影响。结果如图3所示。在一定的葡萄糖质量浓度范围内腺苷产量和菌丝体干重均随着葡萄糖质量浓度的增大而增大。当葡萄糖质量浓度分别为40 g/L、50 g/L时,冬虫夏草菌丝体干重分别达到最大值与次大值,但两者之间无显著性差异(P>0.05);当葡萄糖质量浓度分别为40 g/L时菌丝体干重均显著优于其它水平(P<0.05)(葡萄糖质量浓度50 g/L除外);葡萄糖质量浓度为50 g/L时,更有利于腺苷的合成,但与葡萄糖质量浓度为40 g/L时无显著差异(P>0.05),腺苷作为次生代谢产物,可能与腺苷合成途径中需要耗能有关。当葡萄糖质量浓度超过50 g/L时,均不利于腺苷及菌体生物量的积累。综合菌丝体干重得率考虑,选择葡萄糖质量浓度为40 g/L进行下一步试验。

图3 葡萄糖添加量对冬虫夏草菌生长的影响Fig.3 The effect of the amount of glucose on Ophiocordyceps sinensis cell growth
2.1.4 氮源质量浓度对冬虫夏草菌体生长的影响
向培养基中分别添加不同质量浓度的酵母粉,其它条件相同,研究其对腺苷含量及菌丝体生物量的影响。结果如图4所示。随着酵母粉质量浓度的增加,虫草菌体干重及腺苷产量增加。当酵母粉质量为60 g/L或大于60 g/L时,菌丝体干重达到最大值并趋于平稳,菌体产量显著优于其它酵母粉质量浓度(P< 0.05),说明60 g/L的酵母粉用量能够满足冬虫夏草菌菌丝体的生长。而酵母粉质量为75 g/L时更有利于菌丝体内腺苷含量的累积,但与酵母粉质量浓度为60 g/L、90 g/L时无显著差异(P>0.05)。综合经济性考量,选择酵母粉用量为60 g/L。

图4 酵母粉添加量对冬虫夏草菌生长的影响Fig.4 The effect of the amount of yeast powder on Ophiocordyceps sinensis cell growth
2.1.5 培养温度对冬虫夏草菌生长的影响
冬虫夏草菌来源于高原特有物种冬虫夏草,冬虫夏草主产区平均温度长年低于20℃,使得冬虫夏草菌具“嗜低温”的生物学特性且对温度敏感。因此在进行冬虫夏草菌丝体培养时对温度的筛选极其重要。其它条件相同,分别研究了不同温度对腺苷含量及菌丝体生物量的影响。结果如图5所示。冬虫夏草菌丝体生物量及腺苷含量随着温度的升高而升高,当温度超过19℃时均不利于菌丝体及腺苷的积累,当温度达到22℃时,菌丝体几乎不生长,其对温度的敏感性亦显示了其嗜低温的特性。当培养温度为16℃时,菌体产量显著优于其它温度培养下的菌丝体产量(P<0.05),故选择16℃作为冬虫夏草菌的培养温度。

图5 培养温度对冬虫夏草菌生长的影响Fig. 5 The effect of the temperature on Ophiocordyceps sinensis cell growth
2.1.6 无机盐配比对冬虫夏草菌生长的影响

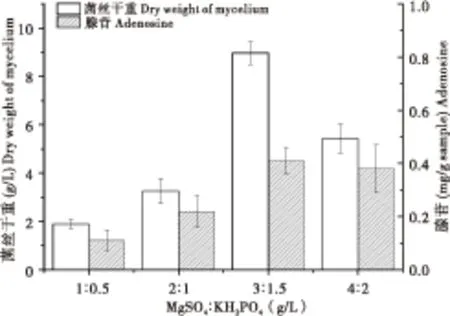
图6 不同无机盐配比对冬虫夏草菌生长的影响Fig.6 The effect of mineral salts ratio on Ophiocordyceps sinensis cell growth
2.1.7 培养时间对冬虫夏草菌生长的影响
在实验过程中选择1-15 d为培养周期,在其它条件不变的情况下,考察其对冬虫夏草菌生长的影响,结果如图7。随着培养时间的推移,冬虫夏草菌丝体及腺苷含量逐渐增加,当到第12天时腺苷含量和菌体干重均达到最大值,此后随着时间的延长腺苷含量及菌体生物量近趋于平稳,因此选择12 d作为培养时间。

图7 培养时间对冬虫夏草菌生长的影响Fig.7 The effect of incubation time on Ophiocordyceps sinensis cell growth
2.2 Plackett-Burman筛选实验
为了筛选出影响冬虫夏草菌体生长的显著因子,根据碳源、氮源浓度、无机盐元素配比、培养温度、培养时间的单因素试验结果,设计n=12的Plackett-Burman筛选实验,每个因子取高(+1),低(-1)2个水平。培养完成后以菌丝体干重生物量为响应值Y。参数和水平设计见表1,实验结果见表2。各因素的效应评价及系数估计见表3。P<0.05说明该因素对菌丝体生物量的影响是显著的,可对该因素作进一步的考察。从表3可得:葡萄糖质量浓度、酵母粉质量浓度、无机盐配比和培养温度对冬虫夏草菌丝体生物量具有显著性影响。
2.3 正交实验
根据Plackett-Burman筛选实验结果,对葡萄糖浓度、酵母粉浓度、培养温度及无机盐配比4个对冬虫夏草菌丝体生长具有显著影响的因素进行正交实验优化。根据单因素中4个因素的试验结果,取各因素的最优值作为中间水平进行L9(34)正交实验。各因素选取水平情况见表,试验结果及分析见表4和表5。
表2 Plackett-Burman设计及响应值
Table 2 Plackett-Burman design and response value

序号Sequencenumber葡萄糖质量浓度(g/L)Theamountofglucose酵母粉质量浓度(g/L)Theamountofyeastpowder培养温度(℃)Temperature培养时间(d)Time无机盐配比(g/L)Mineralsaltsratio菌丝干重(g/L)Dryweightofmycelium1-111-11734±0282-1-1-111914±03131-1-1-11819±03941-111-1761±0445111-11665±036-1111-1762±036711-111821±0338-1-1-1-1-192±03491-11-1-1781±0421011-11-1846±03611-1-1111735±02812-11-1-1-1855±037
注:表中数据为3次重复平均值±标准差。Note:Data in the same column with different capital.

表3 方差分析
注:置信度95%,P<0.05表示显著。Note:The confidence level was 95%,The data was significantly atP<0.05.
表4 正交实验因素水平
Table 4 The factor level of orthogonal experiment

水平Levelsρ(葡萄糖)(g/L)ρ(glucose)Aρ(酵母粉)(g/L)ρ(yeastpowder)B培养温度(℃)TemperatureCρ(MgSO4∶KH2PO4)(g/L)D135551525∶12524060163∶15345651735∶175

表5 正交试验设计及结果分析
注:表中数据为3次重复平均值±标准差。Note:Data in the same column with different capital.
从表4、表5中的结果可表明:葡萄糖是影响冬虫夏草菌丝状菌体生长最显著因素,其余3个因素的显著性影响程度依次是酵母粉、培养温度、无机盐配比。从表5可知4个实验因素的最佳培养组合为:A2B3C3D2,即葡萄糖质量浓度40 g/L,酵母粉质量浓度65 g/L,培养温度17℃,KH2PO4质量浓度1.5 g/L,MgSO4质量浓度3 g/L。按优化后的培养条件,12 d后测得冬虫夏草菌丝状菌体干重平均值为15.67 g/L,腺苷含量平均值为6.52 mg/g样品。比未优化前的菌丝生物量(平均8.54 g/L)提高了83.5%,经统计分析表明效果显著(P<0.05)。
3 结论与讨论
在了解冬虫夏草菌的“嗜低温”、“嗜荤”、“耐缺氧”的生物学特性基础上(柯传奎,2005),确定了培养冬虫夏草菌丝状菌体的最优培养条件:葡萄糖质量浓度40 g/L,酵母粉质量浓度65 g/L,培养温度17℃, KH2PO4质量浓度1.5 g/L,MgSO4质量浓度3 g/L,培养时间12 d,此时丝状菌体干重达15.67 g/L,腺苷含量6.52 mg/g样品。试验结果获得了大量的丝状菌体产物,未见菌丝球形成,可为蝠蛾幼虫接种体的制备提供丝状体菌源。
冬虫夏草菌作为冬虫夏草的唯一无性型菌种现已得到确证(沈南英等,1983;刘锡琎等,1989;李增智等,2000)。但科研工作者为了找寻侵染蝠蛾幼虫的菌种,从上世纪七十年代开始进行了长达二十余年的研究,从冬虫夏草子实体上分离了涉及无性型菌种11属22种真菌,其中包括蝙蝠蛾拟青霉(戴如琴等,1989;戴如琴等,2008)、蝙蝠蛾被孢霉(陈庆涛等,1986)、中国金孢霉(梁宗琦,1991)等真菌,这些菌种存在于冬虫夏草子实体上或伴随着冬虫夏草的成熟(朱佳石等,2007;杨金玲等,2008;高凌等,2011;姚艺桑等,2011;朱佳石等,2012),但以上菌种均不是侵染蝠蛾幼虫最终形成冬虫夏草的无性型菌种。已有研究表明除冬虫夏草菌产生的分生孢子能够侵染蝠蛾幼虫外,冬虫夏草菌丝体亦能侵染蝠蛾幼虫(俞永信,2004),说明采用冬虫夏草菌丝进行侵染成为可能。目前在冬虫夏草菌丝体液态发酵培养方面的报道较多,但部分报道所述菌种非真正的冬虫夏草菌(赵润等,2008;朱朝阳等,2013;王文风等,2016),加之在这些报道中其培养目的是获得具有增强免疫功能的菌粉原料,为了获得最大生物量在培养过程中以高营养、高剪切力、高通氧为主(沈南英等,1998;姚一建等,2006;王晓玲等,2016),致冬虫夏草菌体生长以菌球形态生长为主,按其报道方法培养的冬虫夏草菌菌丝球未见有蝠蛾幼虫被侵染。因此,现有文献报道以生产菌粉原料为目的的培养方法与本文目的不同。最近的研究还显示通过冬虫夏草菌株的筛选,固态发酵培养基的优化,获得较高产分生孢子的菌株,为蝠蛾幼虫的侵染提供大量的分生孢子(张宗耀等,2016),这是目前蝠蛾幼虫被侵染的主要途径,但诱导冬虫夏草菌产分生孢子条件复杂、苛刻,培养时间长,极易污菌,规模化生产难,且冬虫夏草菌多次传代菌种退化严重(吕延华等,2016),分生孢子产量极不稳定。因此,提供一种新的、产量稳定的菌丝原料对接种体的创制具有重要意义。
References)
Chen QT, Wang Q, Liu B. A study on morphology ofMortierellahepialisp. nov.[J].JournalofShanxiUniversity(Natural Science Edition), 1986, 4: 70-74. [陈庆涛, 王琪, 刘波. 蝙蝠蛾被孢霉新种的形态学研究[J]. 山西大学学报 (自然科学版), 1986, 4: 70-74]
Dai RQ, Lan JL, Chen WH,etal. The study on the new species ofPaecilomyceshepialid[J].JournalofChinaAgriculturalUniversity, 1989, 2: 221-224. [戴如琴, 兰江丽, 陈伟华, 等. 蝙蝠蛾拟青霉新种的研究[J]. 北京农业大学学报, 1989, 2: 221-224]
Dai RQ, Li XM, Shao AJ,etal. Nomenclatural validation ofPaecilomyceshepialid[J].Mycosystema, 2008, 5: 641-644.[戴如琴, 李晓明, 邵爱娟, 等. 蝙蝠蛾拟青霉名称的合格化[J]. 菌物学报, 2008, 5: 641-644]
Dong CH, Yao YJ. In vitro evaluation of antioxidant activities of aqueous extracts from natural and cultured mycelia ofCordycepssinensis[J].LWT-FoodScienceandTechnology, 2008, 41 (4): 669-677.
Gao L, Li XH, Zhao JQ,etal.Detection of multipleOphiocordycepssinensismutants in premature stroma ofCordycepssinensisby Massarray SNP MALDI-TOF mass spectrum genotyping[J].JournalofPekingUniversity(Health Sciences), 2011, 2: 259-266. [高凌, 李晓红, 赵建晴, 等. MassARRAY单核苷酸多态性MALDI-TOF质谱基因分型法检测未成熟冬虫夏草子座中多个冬虫夏草菌突变基因型[J]. 北京大学学报(医学版), 2011, 2: 259-266]
Ji DB, Ye J, Li CL,etal. Antiaging effect ofCordycepssinensisextract[J].PhytotherapyResearch, 2009, 23 (1): 116-122.
Ke CK. The Study onOphiocordycepsSinensisin China [M]. Beijing: Science and Technology of China Press, 2005. [柯传奎. 中国冬虫夏草研究[M]. 北京: 中国科学技术出版社, 2005]
Koh JH, Kim JM, Chang UJ,etal. Hypocholesterolemic effect of hot water extract from mycelia ofCordycepssinensis[J].Biological&PharmaceuticalBulletin, 2003, 26: 84-87.
Liang ZQ.A new chrysosporium species isolated fromCordycepssinensis[J].Mycosystema, 1991, 1: 50-56. [梁宗琦. 一个分离自冬虫夏草的金孢霉新种[J]. 真菌学报, 1991, 1: 50-56]
Liu XJ, Guo YL, Yu YX,etal. Isolation and identification ofOphiocordycepssinensisasexual stage[J].Mycosystema, 1989, 1: 35-40. [刘锡琎, 郭英兰, 俞永信, 等. 冬虫夏草菌无性阶段的分离和鉴定[J]. 真菌学报, 1989, 1: 35-40]
Li ZZ, Huang B, Li CR,etal. Molecular evidence for anamorph determination ofCordycepssinensis(Berk. ) SACC I. Relation betweenHirsutellasinensisandC.sinensis[J].Mycosystema, 2000, 1: 60-64. [李增智, 黄勃, 李春如, 等. 确证冬虫夏草无性型的分子生物学证据I. 中国被毛孢与冬虫夏草之间的关系[J]. 菌物系统, 2000, 1: 60-64]
Lv YH, Liang GH, Zhu ZG,etal. The solid fermentation ofOphiocordycepssinensisstrains isolated from different parts of specimen and different generations [J].Mycosystema, 2016, 4: 433-439. [吕延华, 梁关海, 朱志钢, 等. 冬虫夏草不同部位来源的菌株及多次传代固体发酵特性[J]. 菌物学报, 2016, 4: 433-439]
Shen NY, Zeng L, Zhang XC,etal. The separation ofOphiocordycepssinensisfungi[J].EdibleFungi, 1983, 5: 1-5. [沈南英, 曾璐, 张显耻, 等. 冬虫夏草真菌的分离[J]. 食用菌, 1983, 5: 1-5]
Shen NY, Zeng L, Huang R. The fermentation method ofOphiocordycepssinensis[P]. China, 1998, CN1197843,1998-11-04. [沈南英, 曾璐, 黄荣. 中国冬虫夏草真菌的发酵生产方法[P]. 中国, CN1197843,1998-11-04]
Sung GH, Hywel-Jones NL, Sung JM,etal. Phylogenetic classification of Cordyceps and the clavicipitaceous fungi[J].StudiesinMycology, 2007, 57: 5-59.
Wang WF, Xu GH, XU L,etal. A kind of fermentation method onHirsutellasinensispowder[P]. China, 2016, CN1807574, 2016-06-15. [王文风, 徐国华, 徐玲, 等. 一种快速深层液态发酵生产中国被毛孢菌粉的方法 [P]. 中国, 2016, CN105670942A, 2016-06-15]
Wang XL, Zhu CY, Liu GQ,etal. Analysis of key active compounds and fermentation technology ofOphiocordycepssinensismycelium [J].JournalofChineseInstituteofFoodScienceandTechnology, 2016,1: 91-98. [王晓玲, 朱朝阳, 刘高强, 等. 冬虫夏草深层发酵与功效成分分析 [J] . 中国食品学报, 2016, 1: 91-98]
Wu Y, Sun H, Qin F,etal. Effect of various extracts and a polysaccharide from the edible mycelia ofCordycepssinensison cellular and humoral immune response against ovalbumin in mice[J].PhytotherapyResearch, 2006, 20 (8): 646-652.
Wu YL, Ishurd O, Sun CR,etal. Structure analysis and antitumor activity of (1-3)-beta-d-glucans(cordyglucans) from the mycelia ofCordycepssinensis[J].PlantaMedica, 2005, 71 (4): 381-384.
Yang JL, Xiao W, He HX,etal. Molecular phylogenetic analysis ofPaecilomyceshepialiandCordycepssinensis[J].ActaPharmaceuticaSinica, 2008, 4: 421-426. [杨金玲, 肖薇, 何惠霞, 等. 蝙蝠蛾拟青霉与冬虫夏草关系的分子系统学研究[J]. 药学学报, 2008, 4: 421-426]
Yao YJ, Dong CH, Wang B,etal. A kind of cultural method and special medium onOphiocordycepssinensis[P]. China, 2006, CN1807574, 2006-07-26. [姚一建, 董彩虹, 王波, 等. 一种冬虫夏草的培养方法及其专用培养基 [P]. 中国, 2016, CN1807574, 2006-07-26]
Yao YS, Zhou YJ, Gao L,etal. Dynamic alteration of the differential fungal expressions ofOphiocordycepssinensisand its mutant genotypes in stroma and caterpillar during maturation of naturalCordycepssinensis[J].JouralofFungalResearch, 2011, 1: 37-49, 53. [姚艺桑, 周妍娇, 高凌, 等. 冬虫夏草成熟过程中冬虫夏草菌及其突变基因型在子座和僵虫体中的差异表达[J]. 菌物研究, 2011, 1: 37-49, 53]
Yu YX. Studies on artificial culture ofCordycepssinensis[J].JouralofFungalResearch, 2004, 2: 42-46. [俞永信. 人工培养冬虫夏草研究[J] . 菌物研究, 2004, 2: 42-46]
Zhang GQ, Huang YD, Bian Y,etal. Hypoglycemic activity of the fungiCordycepsmilitaris,Cordycepssinensis,TricholomamongolicumandOmphalialapidescensin streptozotocin-induced diabetic rats[J].AppliedMicrobiologyandBiotechnology, 2006, 72 (6): 1152-1156.
Zhang ZY, Liang GH, Liang L,etal. Effects of medium and environmental conditions on the sporulation ofOphiocordycepssinensisin solid fermentation [J].Mycosystema, 2016, 4: 440-449. [张宗耀, 梁关海, 梁蕾, 等. 培养基及培养条件对冬虫夏草固体发酵产分生孢子的影响[J]. 菌物学报, 2016, 4: 440-449]
Zhao R, Guo CJ, Liu GQ. Optimizing on liquid culture media ofCordycepssinensismycelia [J].JournalofTianjinNormalUniversity(Natural Science Edition), 2008, 1: 8-11. [赵润, 郭成金, 刘高强. 冬虫夏草菌丝体液体培养基的优化[J]. 天津师范大学学报(自然科学版), 2008, 1: 8-11]
Zhu CY, Liu GQ, Kang SM,etal. Optimization of fermentation conditions of exo-polysaccharides byOphiocordycepssinensisin liquid fermentation [J].FoodScienceandTechnology, 2013, 5: 18-21. [朱朝阳, 刘高强, 旷思敏, 等. 冬虫夏草真菌产胞外多糖发酵条件的优化[J] . 食品科技, 2013, 5: 18-21]
Zhu JS, Guo YL, Yao YS,etal. Coexistence ofHirsutellasinensisandPaecilomyceshepialiDNA during maturation ofCordycepssinensisand changes in fungal competitive proliferation capability and chemical profiles [J].JournalofFungalResearch, 2007, 4: 214-224. [朱佳石, 郭英兰, 姚艺桑, 等. 冬虫夏草成熟过程中冬虫夏草和蝙蝠蛾拟青霉DNA共存及竞争增殖力、化学成分变化[J]. 菌物研究, 2007, 4: 214-224]
Zhu JS, Zhao JG, Gao L,etal. Dynamically altered expressions of at 6Ophiocordycepssinensismutants in the stroma ofCordycepssinensis[J].JournalofFungalResearch, 2012, 2: 100-112. [朱佳石, 赵家刚, 高凌, 等. 至少6个突变基因型冬虫夏草菌在冬虫夏草子座中表达的动态变化[J]. 菌物研究, 2012, 2: 100-112]
The research on the proliferative culture ofOphiocordycepssinensisfilamentous mycelium
HE Zong-Yi1,2,3,4,LI Li1,2,3,4,ZHANG De-Li1,2,3,4,LI Qing1,MAO Xian-Bing5,CHEN Shi-Jiang1,2,3,4*
(1.Chongqing Academy of Chinese Materia Medica,Chongqing 400065,China;2.Chongqing Key Laboratory of Chinese Medicine Resources,Chongqing 400065,China;3.Chongqing Sub-center of the National Resource Center for Chinese Materia Medica,China Academy of Chinese Medical Sciences,Chongqing 400065,China;4.Ganzi Tibetan Autonomous Prefecture Kangding GonggaOphiocordycepssinensisIndustry Co. LTD.,Ganzi 626000,Sichuan Province,China;5.Southwest Pharmaceutical Co. Ltd.,Chongqing 400000,China)
The herb ofOphiocordycepssinensisis that genusHepialuscaterpillars were infected by parasitic fungusO.sinensisto form a combination of insect and fungi,which it is precious and specific natural resources in China.The asexual stage of the herb ofO.sinensiswasO.sinensiswhich has has two kinds of morphology—the filamentous and pellet mycelium under different nutritional conditions. Moreover,only the filamentous mycelium can infect theHepialuscaterpillars,so it may be applied widely as a new inoculant.However,the defect of slow growth and low yield from the filamentous mycelium ofO.sinensishas hindered its application in the inoculant of theHepialuscaterpillars,and the modern industrial cultivation.In order to improve the yield of theO.sinensismycelium,the nutritional conditions were optimized by orthogonal experiment design. The results showed that the yield of filamentous mycelium reached 15.67 g/L under the conditions of glucose 40 g/L,yeast powder 65 g/L,temperature 17℃,KH2PO41.5 g/L,MgSO43 g/L and incubation time 12 days.The optimized yield of filamentous mycelium in this study will provide a large of raw materials for the preparedHepialusmoths larva inoculant.
Ophiocordycepssinensis;filamentous mycelium;Hepialusmoths larva;culture
重庆市科技平台项目(cstc2014ptyjd10001);重庆市科技惠民计划项目(2013gs500102-D2014-3);重庆市社会民生科技创新专项(cstc2015shmszx120032);重庆市科技攻关项目(cstc2013yykfB110002)
贺宗毅,男,1986年生,硕士,助理研究员,研究方向为冬虫夏草人工培殖,E-mail: cqhcmthzy2004@163.com
* 通讯作者 Author for correspondence, E-mail: shijiangchen@163.com
Received:2016-08-18;接受日期 Accepted:2016-11-21
Q965;S567.3+5
A
1674-0858(2016)06-1205-09
