花期提前对雌雄桑树花部形态及其生物量的影响
2016-12-24吴秋平何纯博王爱玲李大东董廷发
吴秋平,刘 刚,何纯博,王爱玲,周 莎,曾 贞,李大东,胥 晓,董廷发
(1.西华师范大学a.生命科学学院;b.西南野生动植物资源保护教育部重点实验室,四川 南充 637009;2.四川省农业科学院 蚕业研究所,四川 南充 637000)
花期提前对雌雄桑树花部形态及其生物量的影响
吴秋平1,刘 刚2,何纯博1,王爱玲1,周 莎1,曾 贞1,李大东1,胥 晓1,董廷发1
(1.西华师范大学a.生命科学学院;b.西南野生动植物资源保护教育部重点实验室,四川 南充 637009;2.四川省农业科学院 蚕业研究所,四川 南充 637000)
桑树(MorusalbaL.)是我国重要的经济和生态恢复造林树种,近年来已观测到其开花时期提前的现象。作者通过比较桑树在1月下旬(提前开花期)和3月中旬(正常开花期)间雌雄植株的花序特征、单花形态以及生物量积累的差异,分析花期提前对桑树的繁殖器官带来的潜在影响。结果发现:1)在花序特征方面,与正常花相比,提前开花显著降低了雌雄桑树花序长,而对花序柄长及单花总数无显著影响;2)在单花形态方面,提前开花显著降低了雌花子房大小、花被片大小、柱头宽,但显著增加了雄花花粉数量;3)在生物量积累方面,提前开花显著降低了雌雄单花干重、花序干重;4)此外,还发现雌花序干重与子房大小、花被片大小和单花干重均呈显著正相关,花序长与子房大小、花被片大小和单花干重呈显著正相关;雄花序干重分别与单花干重和花被片大小呈显著正相关。结果表明,提前开花会减小桑树雌株的部分花部性状特征及雌雄株的花部生物量积累,增加雄花花粉数量,并且雌株的变化率大于雄株,这种雌、雄株之间的不对称响应是桑树为保证生殖成功而形成的适应性机制。
雌雄异株;桑树;花期;花部特征
花是被子植物的繁殖器官,其通过传粉、受精以及形成种子以进行有性繁殖。植物的花部特征包括花的颜色、大小、形状、花粉数量以及某一特定时刻的花朵开放数量、花朵寿命、花在花序上的排序方式等,这些花部特征会直接影响传粉的效率[1-5]。通常,对于虫媒花而言,花朵较大、花粉数量较多的个体比那些花朵小、花粉数量少的个体更易吸引传粉者的注意[6]。因此,花部特征的变化对植物的成功繁殖具有很大的影响。
导致植物花部特征变化的原因有很多[7-9],花期提前也是很重要的一个因素。但国内外关于花期提前的研究多集中于探究其变化的原因[10-12]。比如高温胁迫导致某些木本植物春季物候提前[13,14],而一些植物的花期也会随CO2浓度升高而提前[15]。目前,鲜有雌雄异株植物花部特征对其花期变化响应的相关报道。由于雌雄植株分别承担着不同的繁育功能,因此,花期提前对其花部形态的影响可能会在性别间呈现显著差异。
桑树(MorusalbaL.)属桑科(Moraceae)桑属(Morus),为雌雄异株植物,因其具有重要的经济价值和生态功能[3,16-17],在我国广泛种植。桑树正常花期为每年的3—4月[18]。近年来,四川省南充地区栽培的桑树的花期出现了提前的现象,其开花期提前到了1月下旬。由于开花期提前可能会潜在地导致植株对繁育器官生物量分配的策略发生变化,从而对花部形态及其生物量的积累造成影响。因此,本文通过比较不同开花期的花部形态及生物量的变化,旨在验证以下科学问题:1)提前开放的雌(雄)花与正常开放的雌(雄)花形态和生物量性状是否有差异?2)不同花期间雌、雄花间的变化率是否一致?研究结果可评估花期改变对该树种繁育能力和桑果产量的潜在影响,以及为品种选育提供理论参考。
1 材料和方法
1.1 研究地概况
研究地位于四川省南充市西华师范大学生命科学学院实验基地(东经106°03′;北纬30°48′),海拔276 m,属亚热带湿润季风气候,年平均气温15.6—17.4℃,年降雨量980—1 150 mm,相对湿度76 %—86 %,年日照时数 1 215—1 530 h[19]。
1.2 实验材料与设计
实验材料来源于四川省农业科学研究院蚕业研究所,品种为沙2×伦9。2015年1月下旬和3月下旬,在实验地内针对同一苗龄(3年生)的雌雄桑苗选取提前开花的植株各4株,植株选取时注意保证花期和植株大小一致。每株随机选取大、中、小3个花序,雌雄花序共48个分别置于FAA固定液中固定24 h,再转移至70%酒精中保存用于测量[20]。
1.3 性状测定及方法
花柄长度和花序长度用游标卡尺(精确到0.001 mm)测量。雌花子房大小(子房长×子房宽)、柱头长宽及花被片大小(花被片长×花被片宽),雄花花药大小(花药长×花药宽)及花被片大小均用体视显微镜(SMZ-168-TL,Motic (厦门)电子有限公司)测量。雄花花粉粒数量为单室花粉粒数量,在光学显微镜(DM500,LEICA)下计数。之后将各实验材料于80℃烘干至恒重,用万分之一的电子天平称量雌雄花的单花干重及单花序干重。
1.4 数据分析和处理
花序及花部的形态特征和生物量在不同花期间的差异均采用独立样本t检验进行比较;花部特征间的相关性均采用Pearson法检验。显著性水平设为α=0.05。统计分析用SPSS 19.0 (IBM公司,美国)。
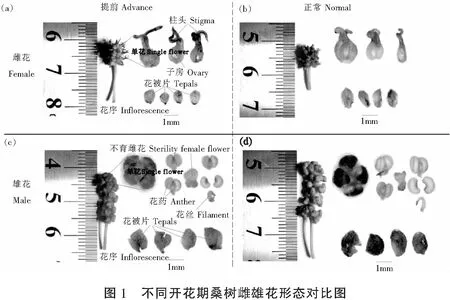
2 结果与分析
2.1 不同花期雌雄花序形态特征
由图1和表1可见,花期提前显著降低了桑树雌雄花序的花序长度(P分别为0.014和0.018),而对花序柄长和单花数量影响不显著。在两种花期中,雌花序的花序长均显著低于雄花序(P<0.001)。
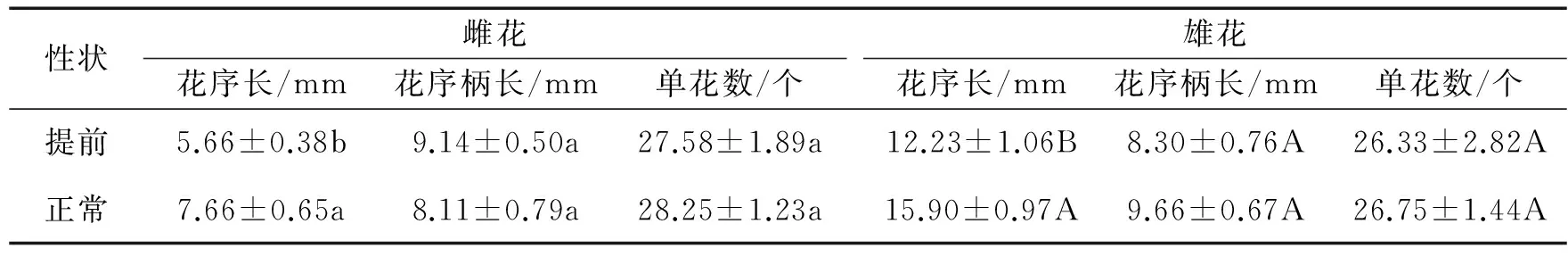
表1 不同花期雌雄桑树的花序形态变化
注:数据为平均值±标准误(n=12),大小写字母分别表示雄花和雌花不同花期间的比较,具有不同字母则表示不同花期间差异显著(P≤0.05)。
2.2 不同花期雌雄单花的形态特征
花期提前对雌雄单花的部分形态特征具有显著影响(图1,表2)。与正常花期的单花相比,提前开花显著缩小了雌花的子房大小、花被片大小和柱头宽度(P=0.003,0.002和0.042),而显著增加了雄花的花粉数量(P=0.008),但对雌花的柱头长度、雄花的花药大小和花被片大小无显著影响。
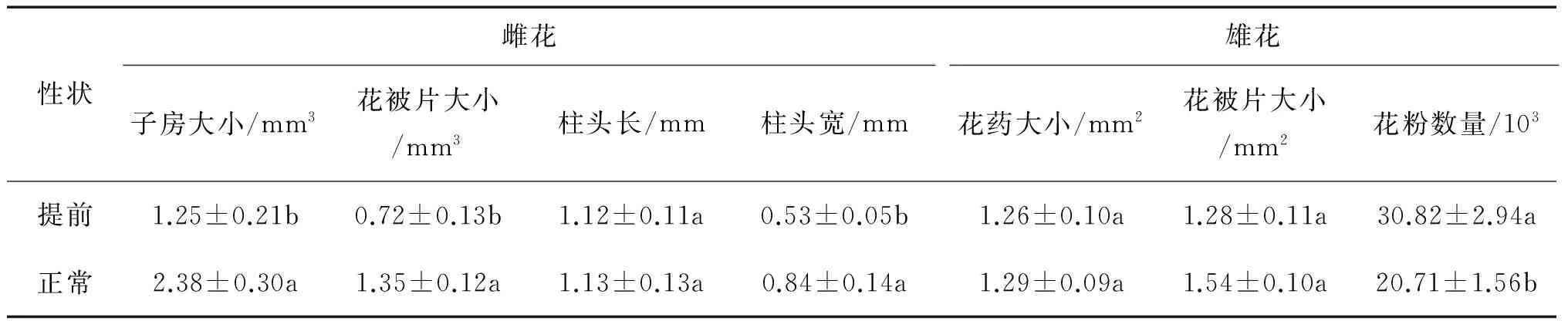
表2 不同花期桑树雌雄花的单花形态变化
注:数据为平均值±标准误(n=12),具有不同字母则表示不同花期间差异显著(P≤0.05)。
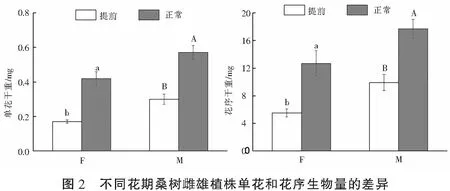
2.3 不同花期雌雄花的生物量
花期提前对桑树花的生物量有显著影响(图2)。与正常花期的花相比,提前开花显著降低了雌花的单花干重和花序干重(P=0.001和0.003)以及雄花的单花干重和花序干重(P=0.013和0.041)。
2.4 不同花期雌雄花部特征的变化率
花期提前对雌花部分性状的影响大于雄花。雌花的花被片大小在不同花期间的变化率显著大于雄花(P=0.02),而雌花的花序柄长、花序长、单花数、单花干重和花序干重在不同花期间的变化率与雄花无显著差异(表3)。
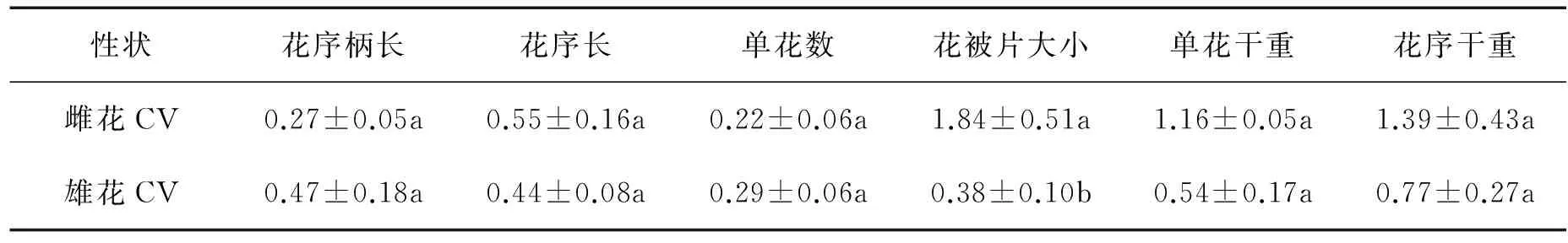
表3 不同花期间雌雄花变化率的比较
注:CV=│提前-正常│/提前,数据为平均值±标准误(n=12)。
2.5 雌雄花花序与单花形态性状的相关性
桑树雌花花序干重与子房大小、花被片大小和单花干重均呈显著正相关,花序长与子房大小、花被片大小和单花干重均呈显著正相关(表4)。花序水平上,花序干重与花序长和单花数均呈显著正相关,花序长与单花数显著正相关。单花水平上,单花干重与子房大小和花被片大小均呈显著正相关,柱头宽与子房大小和花被片大小均呈显著正相关,子房大小与花被片大小显著正相关(表4)。
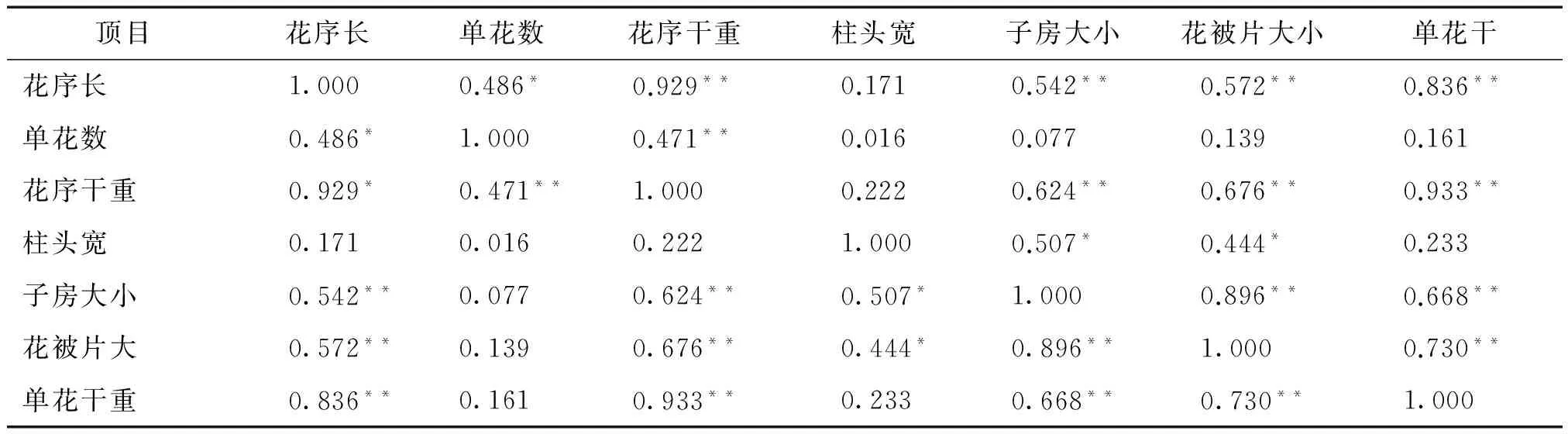
表4 雌花生物量与花形态性状的相关性
注:*:P≤0.05;**:P≤0.01。
由表5可见,桑树雄花花序干重分别与单花干重和花被片大小均呈显著正相关。此外,在花序特征中,花序干重与单花数间呈显著正相关,花序长与花序柄长呈显著正相关。在单花特征中,花药大小与花被片大小和花粉数量均呈显著正相关。

表5 雄花生物量与花形态性状的相关性
注:*:P≤0.05;**:P≤0.01。
3 讨 论
自然界中植物花的特征受到遗传因素和环境因素(传粉者和逆境胁迫等)的共同调节[21]。近年来,由于气候变暖和CO2浓度升高所引起的全球变化对生态系统的影响引起了生态学家的广泛关注。已有研究发现,花对气候的变化非常敏感,这表现在植物的花期、花器官形态及生物量等变化上[22,23]。多种植物的花期会随温度升高、CO2浓度升高等因素而提前,而部分植物花期却延迟[13,24-26]。Fitter和Fitter[27]对英国385种植物进行调查发现,过去十年中这些植物花期平均提前了4.5d,其中16 %提前了15d。而王桔红和陈文[28]对四种菊科植物比较发现,三叶鬼针草(Bidenspilosa)、宽叶鼠曲草(Gnaphaliumadnatum)和夜香牛(Vernoniacinerea)在干旱的环境中,花序生物量分配均有减小的趋势。根据本实验地生长室近三年的气候数据来看,年均温和CO2浓度均逐渐升高,而年均湿度逐渐降低,因此南充地区桑树花期提前可能与气候变化有关,至于是哪种气候变化因子改变了花的物候和花部特征,有待进一步通过控制性实验进行确定。
本实验发现,与正常花期相比,花期提前显著降低了桑树雌株的部分花部性状(如花序长、子房大小、花被片大小、柱头宽、单花干重、单花序干重)以及雄株的单花干重和单花序干重,但显著提高了雄花的花粉数量。并且,雌花的花被片大小在不同花期间变化率显著大于雄花,单花干重和单花序干重的变化率有大于雄花的趋势。这表明桑树雌雄植株花部特征对花期提前存在不对称的响应机制。研究发现,在桑树雌花序方面,其花序干重、花序长、子房大小和花被片大小呈现显著的正相关变化,柱头宽、子房大小和花被片大小也呈现显著的正相关变化。在雄花序方面,花序干重分别与单花干重、单花数和花被片大小均呈显著正相关,花序长与花序柄长呈显著正相关,花药大小与花被片大小和花粉数量均呈显著正相关。这些结果表明,花部特征之间共同作用,以适应花期的改变,导致雌雄植株出现对花期提前的不对称响应机制。
作为植物的有性生殖器官,花在形态上的差异可能会导致传粉机制的改变,从而影响植物的生殖过程及种群发展[1,29,30]。桑树花既是虫媒花,也是风媒花,其花在成熟后,花药破裂,所含的大量花粉喷出,并且它的花粉互不粘连[31],故扩散地越广,雌花的接收面积越广,对它的传粉及繁殖越有利。从本实验结果来看,花期提前后,雌花的子房大小和柱头宽显著降低,这将减小雌花对花粉的接收面积和接收效率。但为了适应环境变化,保证生殖成功,植物有其相应的适应性机制。花期提前后,雌花花被片的减小以及雄花花粉数量的增加,正好增加了花粉落到较小柱头上的几率,这种雌雄花对花期提前的不对称响应,可减小由于花期提前对其传粉以及繁殖带来的不利影响。
综上,花期提前后对花器官资源分配的减少,既适应了环境改变,又保证了正常花期对于资源的需求,并且,雌雄花对花期提前的不对称响应,可减小由于花期提前对其传粉以及繁殖带来的不利影响。
[1] FENSTER C B,ARMBRUSTER W S,WILSON P,et al.Pollination syndromes and floral specialization[J].Annual Review of Ecology,Evolution and Systematics,2004,12(35):375-403.
[2] 李肖夏.淫羊藿属植物的花部特征及其传粉适应[D].武汉大学,2013.
[3] 黄自然,杨 军,吕雪娟.桑树作为动物饲料的应用价值与研究进展[J].蚕业科学,2006,32(3):377-385.
[4] 张志强,李庆军.花寿命的进化生态学意义[J].植物生态学报,2009,33(3):598-606.
[5] 袁一斌,慕军鹏,彭幼红,等.多齿马先蒿与刺齿马先蒿花部形态、昆虫访花频率及种子产量比较研究[J].应用与环境生物学报,2011,17(4):467-472.
[6] IRWIN R E,ADLER L S,AGRAWAL A A.Community and evolutionary ecology of nectar[J].Ecology,2004,85(6):1477-1478.
[7] 张桂莲,张顺堂,肖浪涛,等.花期高温胁迫对水稻花药生理特性及花粉性状的影响[J].作物学报,2013,39(1):177-183.
[8] GOLDHAMER D,SMITH T.Single-season drought irrigation strategies influence almond production[J].California Agriculture,1995,49(1):19-22.
[9] PANDEY D M,GOSWAMI C L,KUMAR B.Physiological effects of plant hormones in cotton under drought[J].Biologia Plantarum,2003,47(4):535-540.
[10] 刘建福,陈李林,汤青林,等.不同土壤水分胁迫对澳洲坚果花期生长的影响[J].西南农业大学学报(自然科学版),2004,26(6):735-739.
[11] 陈正洪,肖 玫,陈 璇.樱花花期变化特征及其与冬季气温变化的关系[J].生态学报,2008,28(11):5209-5217.
[12] 徐维新,辛元春,张 娟,等.近20年青藏高原东北部禾本科牧草生育期变化特征[J].生态学报,2014,34(7):1781-1793.
[13] 郑景云,葛全胜,赵会霞.近40年中国植物物候对气候变化的响应研究[J].中国农业气象,2003,24(1):28-32.
[14] 杨 帅,王碧霞,胥 晓,等.葎草雌雄植株开花物候和花器官对干旱的响应差异[J].植物分类与资源学报,2014,36(5):653-660.
[15] 蒋高明,韩兴国,林光辉.大气CO2浓度升高对植物的直接影响—国外十余年来模拟实验研究之主要手段及基本结论[J].植物生态学报,1997,21(6):2-15.
[16] 秦 俭,何宁佳,黄先智,等.桑树生态产业与蚕丝业的发展[J].蚕业科学,2010,36(6):984-989.
[17] 刘 芸.桑树在三峡库区植被恢复中的应用前景[J].蚕业科学,2011,37(1):93-97.
[18] 中国科学院中国植物志编辑委员会.中国植物志[M].北京:科学出版社,1998.
[19] 陈梦华,秦 芳,刘 刚,等.桑树雌雄幼苗抗氧化酶系统和光合色素对UV-B辐射的响应差异[J].西华师范大学学报(自然科学版),2014,35(4):327-332.
[20] RUDALLl P J,BATEMAN R M,FAY M F,et al.Floral anatomy and systematics of Alliaceae with particular reference to Gilliesia,a presumed insect mimic with strongly zygomorphic flowers[J].American Journal of Botany,2002,89(12):1867-1883.
[21] 张栩佳,胡灵芝,陈哲皓,等.花器官大小调控机制的研究进展[J].植物生理学报,2014,50(6):691-697.
[22] INOUYE D W.Effects of climate change on phenology,frost damage,and floral abundance of montane wildflowers[J].Ecology,2008,89(2):353-62.
[23] MU J,PENG Y,XI X,et al.Artificial asymmetric warming reduces nectar yield in a Tibetan alpine species of Asteraceae[J].Annals of Botany,2015.(online)
[24] LEGAVE J M,GU ÉON Y,MALAGI G,et al.Differentiated responses of apple tree floral phenology to global warming in contrasting climatic regions[J].Frontiers in Plant Science,2015,60(50):1678-1685.
[25] MARC J,GIFFORD R M.Floral initiation in wheat,sunflower,and sorghum under carbon dioxide enrichment[J].Canadian Journal of Botany,1984,62(1):9-14.
[26] 杨建莹,梅旭荣,刘 勤,等.气候变化背景下华北地区冬小麦生育期的变化特征[J].植物生态学报,2011,35(6):623-631.
[27] FITTER A H, FITTER R S R. Rapid changes in flowering time in British plants[J]. Science, 2002, 296(5573): 1689-1691.
[28] 王桔红,陈 文.四种菊科植物开花期构件生物量及表型可塑性比较[J].生态学杂志,2014,33(8):2031-2037.
[29] 孙 颖.百子莲繁殖生物学研究[D].东北林业大学,2009.
[30] GOODWILLIE C,SARGENT R D,ECKERT C G,et al.Correlated evolution of mating system and floral display traits in flowering plants and its implications for the distribution of mating system variation[J].New Phytologist,2010,185(1):311-321.
[31] 代君君,范 涛,章玉萍,等.桑树蜜源植物的特性与利用[J].中国蜂业,2012,34(63):27-29.
Effects of Advancing Flowering Date on Floral Morphology and Biomass in Male and Female Mulberry
WU Qiuping1,LIU Gang2,HE Chunbo1,WANG Ailing1,ZHOU Sha1,ZENG Zhen1,LI Dadong1,XU Xiao1,DONG Tingfa1
(1.a.College of Life Sciences,b.Key Laboratory of Southwest China Wildlife Resources Conservation,China West Normal University,Nanchong Sichuan 637009,China;2.Sericultural Research Institute,Sichuan Academy of Agricultural Sciences,Nanchong Sichuan 637000,China)
In recent years,the phenomenon of advancing flowering date (AF) caused by global climate change is observed in the Mulberry (MorusalbaL.) species,an important economic and ecological restoration species in China.To evaluate the potential effects of AF on the reproductive organs of Mulberry,the floral morphology and biomass in male and female Mulberry in late January (advancing flowering date,AF) or mid-March (normal flowering date,NF) were investigated.The results showed that:1) inflorescence length in AF was significantly lower than that in NF,while inflorescence stalk length and number of single flowers per inflorescence were not significantly different between AF and NF;2) compared to flowers in NF,the ovary size,tepal size and stigma width of female flowers in AF were lower,but the pollen number of male flowers in AF were higher;3) dry mass of single flower and inflorescence of female and male flowers in AF were significantly lower than that in NF;4) in addition, inflorescence dry mass in female plants was positively correlated with ovary size,tepals size and dry mass of single flowers,and inflorescence length was positively correlated with ovary size,tepals size and dry mass of single flowers;in male plants,inflorescence dry mass was significantly positively correlated with dry weight of single flowers and tepal size.The results suggest that part traits of female floral morphology and biomass of female and male flowers in AF were lower than that in NF,while the pollen number of male flowers in AF were higher,and flowers suffered more negative effects in morphology and biomass than males in AF.The asymmetric responses between female and male flowers of Mulberry by AF may ensure its reproductive success.
dioecy;Mulberry;flowering phenology;floral traits
1673-5072(2016)03-0258-06
2016-04-15 基金项目:四川省青年科技基金资助项目(2012JQ0045);四川省教育厅创新团队项目(14TD0015) 作者简介:吴秋平(1990—),女,四川资阳人,硕士研究生,主要从事植物生理生态研究。 通讯作者:董廷发(1985—),男,云南曲靖人,博士,主要从事植物生理生态研究。E-mail:dongfar@163.com
Q944.3
A
10.16246/j.issn.1673-5072.2016.03.005
