不同苹果矮化砧木苗抗寒性比较
2016-12-20冯梅曹亚军陈刚肖丽娟王华强黄鹏
冯梅,曹亚军,陈刚,肖丽娟,王华强,黄鹏
(兵团第一师农业科学研究所,新疆阿拉尔843300)
不同苹果矮化砧木苗抗寒性比较
冯梅,曹亚军,陈刚,肖丽娟,王华强,黄鹏
(兵团第一师农业科学研究所,新疆阿拉尔843300)

本试验通过对不同苹果矮化砧木苗叶片的可溶性糖、可溶性蛋白、游离脯氨酸和丙二醛含量以及过氧化物酶活性的测定,采用隶属函数值综合评价方法,对8种砧木苗进行综合评价。结果表明,八棱海棠+SH1抗寒性最强,YB0009次之,平邑甜茶最差。在基砧上嫁接中间砧,可有效提高砧木苗的抗寒性;不同砧木苗中,在相同中间砧的条件下,八棱海棠为基砧的砧木苗的抗寒性高于以平邑甜茶为基砧砧木苗。
苹果;矮化砧;生理生化指标;抗寒性
中国苹果的栽培面积及产量均居世界第一位,是当之无愧的全球苹果第一生产大国[1],而新疆是我国苹果重要的产区之一。目前世界苹果主要以矮化密植栽培为主,而矮化砧木是矮化密植栽培的主要方式。由于不同砧木的特性各不相同,抗寒性也有差别,且我国复杂的地理条件及气候条件,如果矮化砧木选择不当,在生产过程中,会造成矮化苹果植株遭受不同程度的冻害,尤其是在高纬度栽培区,其抗寒力决定着苹果植株的越冬安全、树势及产量等。因此,目前迫切需要选育抗寒性强的矮化砧木,以促进新疆乃至北方地区苹果矮化密植集约化栽培进程。
植物的抗寒性具有明显的季节性[2],若与环境极端低温的年周期不同步[3],就极易形成冻害。目前,植物主要是经过或不经过冷冻处理的方法来估测其抗寒性[2],且在抗寒机理、抗寒性鉴定、抗寒资源评价及抗寒性遗传育种等领域做了大量研究工作,并取得了良好的进展[4],认为植株叶片或枝条的电导率大小[5-9]、可溶性糖、可溶性蛋白、丙二醛和游离脯氨酸含量以及酶活性等[10-18]与植物抗寒能力存在相关性,可作为衡量植物抗寒能力的主要指标。随着苹果的矮砧密植趋势的加大,一些科研机构及院所通过引入国内外优良矮化砧木,对其进行抗寒性鉴定和抗寒生理的评价等,最终提出部分抗寒力较强的矮化砧木[19-21],由于我国苹果栽培区域化严重。因此,结合各地区区域特点,筛选抗寒能力强的砧木是决定苹果植株越冬安全、树势及产量的关键。目前,关于矮化砧资源的抗寒性研究报道较少。本研究对从国内各地引入的苹果抗寒矮化砧进行抗寒性的比较试验和评价,以期为新疆南疆乃至北方地区整个寒地苹果抗寒矮化栽培发展有所裨益。
1 材料与方法
1.1试验材料
供试苹果矮化砧木品种为:八棱海棠、八棱海棠+SH1、八棱海棠+SH12、八棱海棠+SH6、八棱海棠+Y-1、Y-B0009、Y-C0002、平邑甜茶、平邑甜茶+SH1、平邑甜茶+Y-1,均采自于新疆兵团第一师农业科学研究所的苹果矮砧资源圃。2015年9月初选取生长健壮、粗细一致的一年生枝条上的叶片,叶片健康、完整,其中中间砧为SH系的砧木叶片表面有一层绒毛。
1.2测定方法
将采集的叶片依次用自来水、蒸馏水冲洗,并用纱布擦干,放入塑料袋中,于-80℃下保存待用。可溶性蛋白含量测定采用考马斯亮蓝G-250染色法[22]、可溶性糖含量的测定采用蒽酮比色法[23]、游离脯氨酸含量的测定采用磺基水杨酸法[24]、过氧化物酶(POD)含量测定采用愈创木酚比色法[25]、丙二醛含量的测定采用硫代巴比妥酸[26]。测定时每品种均重复3次。
1.3数据处理
采用Exce12003和DPS7.05进行数据处理与统计分析,多重比较采用LSD法。同时将整理后的数据用模糊数学隶属度公式[27-28]进行定量转换,再将各指标隶属函数值取平均值进行无性系间相互比较,其计算公式为:

如果某一指标与评判结果为负相关,则用反隶属函数进行定量转换,其计算公式为:
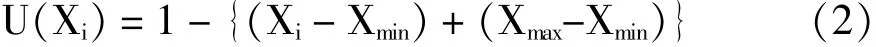
式中(1)(2)中:U(Xi)为隶属函数值,Xi为无性系某项指标测定值,Xmax和Xmin为所有参试无性系中某一指标的最大值和最小值。
2 结果与分析
2.1不同砧木苗叶片可溶性蛋白含量的差异
蛋白质含量的增加,有助于加强细胞的保水力,提高细胞内的束缚水含量,降低冰点,并可导致细胞液过冷的形成,对发展高水平的抗寒性具有重要作用。从图1可以看出,不同砧木苗叶片可溶性蛋白含量YB0009最高,八棱海棠+SH12最低。同一基砧不同中间砧叶片可溶性蛋白含量存在显著差异:以平邑甜茶为基砧的砧木苗中,可溶性蛋白含量平邑甜茶+Y1显著高于平邑甜茶+SH1和对照平邑甜茶;在自根砧苗中,可溶性蛋白含量YB0009显著高于YC0002。在同一中间砧不同基砧苗中叶片可溶性蛋白含量存在差异:以Y1为中间砧的苗中,叶片可溶性蛋白含量平邑甜茶+Y1显著高于八棱海棠+Y1。说明,中间砧和基砧的种类共同影响砧木苗叶片可溶性蛋白含量,且在基砧上嫁接中间砧可提高砧木苗叶片可溶性蛋白含量。
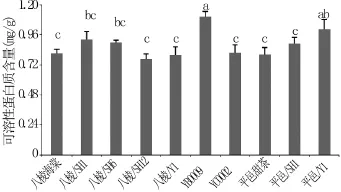
图1 不同砧木苗叶片可溶性蛋白含量的差异
2.2不同砧木苗叶片可溶性糖含量的差异
可溶性糖的含量与抗寒性存在平行现象,抗寒性强的品种,可溶性糖的含量高[29]。从图2可以看出,不同砧木苗中叶片可溶性糖含量八棱海棠+Y1最高,八棱海棠+SH6最低。同一基砧不同中间砧砧木苗叶片可溶性糖含量存在显著差异;以八棱海棠为基砧的砧木苗中,各砧木苗显著低于对照八棱海棠,且各砧木苗间也存在显著性差异;以平邑甜茶为基砧的砧木苗中,叶片可溶性糖含量平邑甜茶+ SH1显著低于对照平邑甜茶,且平邑甜茶+SH1与平邑甜茶+Y1间存在显著性差异。在同一中间砧不同基砧砧木苗叶片可溶性糖含量存在显著差异;以SH1为中间砧的砧木苗中,叶片可溶性糖含量八棱海棠+SH1显著高于平邑甜茶+SH1;以Y1为中间砧的砧木苗中,叶片可溶性糖含量八棱海棠+Y1显著高于平邑甜茶+Y1。说明,中间砧和基砧的种类共同影响砧木苗叶片可溶性糖含量,且在基砧上嫁接中间砧可降低砧木苗叶片可溶性蛋白含量。
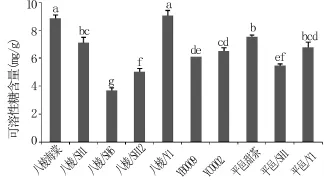
图2 不同砧木苗叶片可溶性糖含量的差异
2.3不同砧木苗叶片过氧化物酶活性的差异
过氧化物酶是植物体内普遍存在活性较高的一种酶[30],它能够清除超氧自由基形成的H2O2,还有分解叶绿素及生长素的功能,POD活性越高抗寒能力越强。从图3可以看出,不同砧木苗中叶片过氧化物酶活性八棱海棠+Y1最高,八棱海棠+SH12最低。在同一基砧不同中间砧砧木苗叶片过氧化物酶活性存在显著差异:以八棱海棠为基砧的砧木苗中,叶片过氧化物酶活性八棱海棠+Y1显著高于对照八棱海棠;八棱海棠+SH12显著低于对照八棱海棠,且八棱海棠、八棱海棠+SH1与八棱海棠+SH6、八棱海棠+SH12、八棱海棠+Y1间以及八棱海棠+ SH6、八棱海棠+SH12、八棱海棠+Y1间存在显著性差异;在自根砧苗中,叶片过氧化物酶活性YB0009显著低于YC0002。在同一中间砧不同基砧砧木苗叶片过氧化物酶活性存在显著差异:以Y1为中间砧的砧木苗中,叶片过氧化物酶活性八棱海棠+Y1显著高于平邑甜茶+Y1。说明,中间砧和基砧的种类共同影响砧木苗叶片过氧化物酶活性。
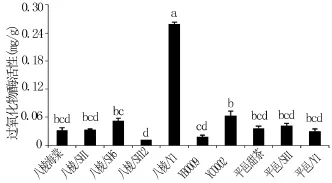
图3 不同砧木苗叶片过氧化物酶活性的差异
2.4不同砧木苗叶片丙二醛含量的差异
丙二醛是膜脂质过氧化的产物,其含量的多少是膜脂质过氧化作用强弱的一个重要指标。从图4可以看出,不同砧木苗中叶片丙二醛含量YC0002最高,八棱海棠+SH12最低。在同一基砧不同中间砧砧木苗叶片丙二醛含量存在显著差异:以平邑甜茶为基砧的砧木苗中,叶片丙二醛含量平邑甜茶+ SH1、平邑甜茶+Y1显著高于对照平邑甜茶。在同一中间砧不同基砧砧木苗叶片丙二醛含量存在显著差异;以SH1为中间砧的砧木苗中,叶片丙二醛含量平邑甜茶+SH1显著高于八棱海棠+SH1。说明,中间砧和基砧的种类共同影响砧木苗叶片丙二醛含量。

图4 不同砧木苗叶片丙二醛含量的差异
2.5不同砧木苗叶片脯氨酸含量的差异

图5 不同砧木苗叶片脯氨酸含量的差异
从图5可以看出,不同砧木苗中叶片脯氨酸含量八棱海棠+SH6最高,YB0009最低。在同一基砧不同中间砧砧木苗叶片脯氨酸含量存在显著差异:以八棱海棠为基砧的砧木苗中,各砧木苗叶片脯氨酸含量与对照八棱海棠间无显著差异,但八棱海棠+SH6与八棱海棠+SH1、八棱海棠+Y1存在显著性差异。在同一中间砧不同基砧砧木苗叶片脯氨酸含量存在差异。说明,中间砧和基砧的种类共同影响砧木苗叶片脯氨酸含量。
2.6苹果矮化砧木品种间叶片各生理指标间的相关性分析
从表1可以看出,5个生理生化指标之间存在相关性,其中蛋白质含量与可溶性糖、脯氨酸和POD含量呈负相关,与丙二醛含量呈正相关;可溶性糖含量与POD含量呈正相关,与丙二醛和脯氨酸含量呈负相关;POD含量与丙二醛含量呈正相关,与脯氨酸含量呈负相关;丙二醛含量与脯氨酸含量呈负相关。丙二醛以及脯氨酸含量与可溶性蛋白含量相关性最高,分别为0.503 9、-0.515 7。
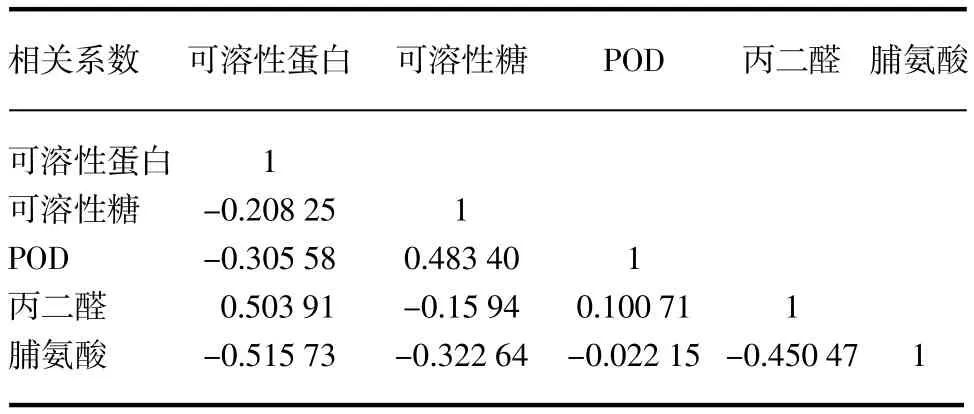
表15 个生理生化指标的相关系数矩阵

表2 不同砧木苗叶片生理指标隶属度的综合评价
2.7不同砧木苗抗寒性的综合评价
不同植物其抗寒性不同,目前可通过可溶性糖、游离脯氨酸、丙二醛和可溶性蛋白质等含量以及过氧化物酶活性来估测植物的抗寒性[10-18],但单从某一指标含量去衡量其抗寒性,不科学,具有明显的片面性。在该研究中,不同砧木苗的生理指标含量均存在差异(如图1~5所示)。因此采用隶属函数值综合评价方法,就10种砧木苗的生理指标进行综合评价,即将各指标的平均数值换算成隶属函数值,取各指标隶属度的平均值作为10种砧木苗的综合评定标准。经综合评价发现八棱海棠+Y1抗寒性最强,YB0009次之,平邑甜茶最差。在基砧上嫁接中间砧,可有效提高柘木苗的抗寒性;不同砧木苗在相同中间砧的条件下,八棱海棠为基砧的苗抗寒性高于以平邑甜茶为基砧的苗。
2.8不同砧木苗越冬抗寒性鉴定
在冬季露地不加任何保护措施的条件下,于次年4月通过对砧木的越冬存活率进行调查(见表3),发现除平邑甜茶+SH1和平邑甜茶+Y-1外其余的均未受到冻害,并能正常生长,其中平邑甜茶+ SH1受冻率严重,达到54.55%,且不可恢复。
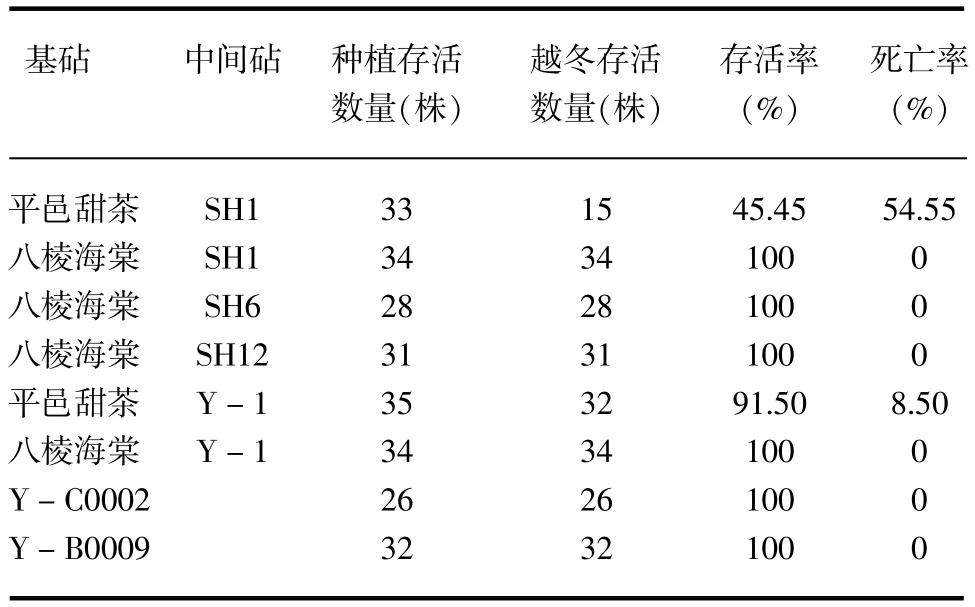
表3 不同矮化苹果砧木越冬存活率比较
3 结论与讨论
抗寒性是植物可以忍受严寒的冷冻温度。是一系列的反应体现在植物生理生化反应中的结果[3],往往受植物体内许多基因共同调控,同时也可能受外部的环境因素的影响或限制[31-32]。
不同树种间、同一果树生长周期的不同阶段以及果树不同组织器官的抗寒性也存在差异。而测得果树抗寒性的信息,对于研究抗寒育种的意义重大[32]。
当气温降低时,植物生长受到影响,植物体内一方面诱导活性氧清除酶类的活性升高及抗氧化物质含量的增加,有效地清除活性氧[33];另一方面,通过积累或分解渗透调节物质,调节细胞渗透平衡[34],来共同缓解逆境胁迫对植物的伤害。一般认为,可溶性糖、游离脯氨酸、丙二醛和可溶性蛋白质等含量以及过氧化物酶活性与植物的抗逆性密切相关[10-18],较高含量的脯氨酸、可溶性糖、可溶性蛋白质、游离脯氨酸和过氧化物酶活性等以及较低含量的丙二醛是植物抗寒性增强的内在原因。因此,通过检测其含量,来作为预测果树的抗寒性的重要指标。但是由于植物的生理过程较为复杂,植物的抗寒性受很多因素影响,孤立地用单一指标很难反映植物的抗寒本质。因此,本研究通过对矮化砧木苗叶片的可溶性蛋白、可溶性糖、丙二醛和游离脯氨酸含量以及过氧化物酶活性的测定,采用隶属函数值综合评价方法,将各指标的平均数值换算成隶属函数值,取各指标隶属度的平均值作为10种砧木苗的综合评定标准。经综合评价发现八棱海棠+Y1抗寒性最强,YB0009次之,平邑甜茶最差。在基砧上嫁接中间砧,可有效提高苗的抗寒性;不同砧木苗中,在相同中间砧的条件下,八棱海棠为基砧苗的抗寒性高于以平邑甜茶为基砧苗。
[1]中华人民共和国国家统计局.中国统计年鉴(2008)[M].北京:中国统计出版社,2008.
[2]张钢.国外木本植物抗寒性测定方法综述[J].世界林业研究,2005,18(5):14-20.
[3]邱乾栋,吕晓贞,臧德奎,等.植物抗寒生理研究进展[J].山东农业科学,2009,8:53-57.
[4]潘杰,简令成.植物寒害和抗寒机制中膜与蛋白质研究的进展[J].植物学通,1990,7(1):1-5.
[5]李喜森,荣志祥,李恩生,等.苹果矮化砧木77-34选育研究[J].北方果树,2004(4):4-9.
[6]沙广利,黄粤,李凌.苹果新矮砧抗寒性电导法测定[J].中国农学通报,2000,16(1):51-52.
[7]曲柏宏,朴红权,代志国,等.利用电导法测定苹果新品种的抗寒性[J].北方果树,1998(1):5-6.
[8]郁俊谊,张继谢.秦冠和富士苹果临界致死低温测定[J].陕西农业科学,1994(2):28.
[9]牛立新,张延龙.苹果品种抗寒性测定及其评价方法探讨[J].落叶果树,1996(增刊):1-2.
[10]刘大林.低温胁迫下番石榴叶片生理生化变化的探讨[J].林业科学,2003,39(1):38-41.
[11]张基德,李玉梅,陈艳秋,等.梨品种枝条可溶性糖、脯氨酸含量变化规律与抗寒性的关系[J].延边大学农学学报,2004,12(4):281-285.
[12]王丽雪,李荣富,张福仁.葡萄枝条中蛋白质(过氧化物酶活性)变化与抗寒性的关系[J].内蒙古农牧学院学报,1996,17(1):45-49.
[13]姚胜蕊,曾骧,简令成.桃花芽越冬过程中糖积累和质壁分离动态与品种抗寒性的关系[J].果树科学,1991,8(1):13-18.
[14]周碧燕,陈杰忠,季作梁,等.香蕉越冬期间SOD活性和可溶性蛋白质含量的变化[J].果树科学,1999,16(3):192-196.
[15]刘艳,赵虎成.梨枝条中淀粉(还原糖及脂类物质的动态变化与抗寒性关系[J].内蒙古农业大学学报,2002,23(1):57-60.
[16]何开跃,李晓储,黄利斌,等.冷冻胁迫对福建柏苗可溶性糖和丙二醛(MDA)含量的影响[J].江苏林业科技,2000(6):6-8.
[17]武维华.植物生理学[M].北京:科学出版社,2003.
[18]谢晓金,李映雪,张耀鸿.园林植物抗寒性测定技术的研究[J].福建林业科技,2008(9):244-247.
[19]李登科.SH系苹果矮化砧的主要性状及其利用技术[J].北方果树,1994(1):8-10.
[20]林凤起.苹果矮化砧GM-256经济性状的研究[J].北方园艺,1993(2):5-8.
[21]杨廷桢.早果矮化苹果矮化砧木Y系选育研究初报[J].中国农学通报,2006,22(6):273-276.
[22]张志良.植物生理学实验指导(第二版)[M].北京:高等教育出版社,1990.160-162.
[23]上海植物生理学会.植物生理学实验手册[M].上海:上海科学技术出版社,1985.134-136.
[24]张殿忠,汪沛洪,赵会贤.测定小麦叶片游离脯氨酸的方法[J].植物生理学通讯,1990,26(4):62-65.
[25]高俊凤.植物生理学实验技术[M].西安:世界图书出版公司,2000.
[26]赵世杰,史国安,董新纯.植物生理学实验指南[M].北京:中国农业科学技术出版社,2002.
[27]高建社,王军,周永学,等.5个杨树无性系抗旱性研究[J].西北农林科技大学学报:自然科学版,2005,33(2):112-116.
[28]武媛林.酸枣种质资源的评价及选优[D].保定:河北农业大学,2008.
[29]粟本文.茶树抗寒性研究概况[J].茶树科学简报,1991(4)22-28.
[30]孙昌祖,刘家琪.低温胁迫对青杨叶片O2、MDA、膜透性、叶水势及保护酶的影响[J].内蒙古林学院学报,1998,20(3):32-36.
[31]Lindén L.Measuring cold hardiness in woody plants[D]. Doctorial Thesis,2002.
[32]Zhang G.A review of methods for measuring frost hardiness in woody plants abroad[J].World Forest Science and Research,2005,18(5):14-20.
[33]邓雪柯,乔代蓉,李良,等.低温胁迫对紫花首蓓生理特性影响的研究田[J].四川大学学报:自然科学版,2005,42(1):190-194.
2016—10—11
