史氏鲟的早期发育异速生长模式及生态学意义
2016-12-16韩骥王云山李文龙胡光源李育东
韩骥+王云山+李文龙+胡光源+李育东

摘要:使用显微镜和解剖镜对史氏鲟仔鱼期、稚鱼期发育过程进行连续观察,使用目微尺测量仔鱼多项数量性状,利用观察到的形态和数据统计分析结果进行生态学研究。实验结果表明,史氏鲟的早期发育分两个阶段,即胚后期孵出仔鱼(体长10.4~11.2 mm)到开食阶段(体长21.1~22.3 mm)和稚鱼阶段(指外源营养阶段到体态形成阶段)。仔鱼期较稚鱼期和幼鱼期在形态发生和分化上更强烈。优先发展重要的器官,然后发展次要器官是一种真骨鱼类共有的发育模式,我们把这种发育模式称之为异速生长模式。通过研究史氏鲟仔鱼的异速生长模式,讨论了异速生长的生态学意义,证明了早期生命阶段的形态发育和增长模式与当前需要有着密切关系的假设。
关键词:史氏鲟;仔鱼;异速生长;生态学
史氏鲟(Acipenser schrencki Brandt)属鲟科(Acipenseridae)鲟属,原产于黑龙江及其相通的泡沼湖泊,松花江和乌苏里江也有少量分布,是一种经济价值极高的鱼类。20世纪80年代末开始,黑龙江省水生动物资源增殖保护站(原黑龙江省特产鱼类研究所)对史氏鲟人工繁殖、人工增养殖技术进行了大量的卓有成效的研究,并将其技术推广到全国范围,获得了极大的成功[1]。实践证明史氏鲟适合于在全国范围内推广养殖,发展前景广阔。但是,史氏鲟苗种培育难度很大,技术要求很高。从1998年起,作者就在全国范围内率先突破了史氏鲟生产性育苗的技术难关[2],并且对史氏鲟仔鱼发育特点及生态学意义作了深入研究。
在史氏鲟早期生活史中,仔鱼是兼营内源营养和外源营养的,在两种营养形式转换过程中导致仔鱼从孵出起就有了剧烈的形态变化,表现在趋流性、趋光性和游动能力等方面的几个跳跃性行为改变。在许多真骨鱼类早期发生阶段,主要的行为改变是与其形态变化同步的。这种同步变化可以确保鱼类在发育阶段能够占有各自的生存环境[3]。鱼类在发育的早期阶段有很多数量性状发生改变,刚孵出的个体、仔鱼的体形到幼鱼、成鱼都有明显的进行性变化,一般认为为适应生存而进行的选择性生长是真骨鱼类仔鱼的共性[4]。鱼类身体比例的巨大变化与鱼类生长密切相关,其相对生长被定义为异速生长。这在仔鱼发育过程中是必然的,保证基本功能的重要器官优先发育,接下来发育的是对生存起次要作用的器官。一般假设这种异速生长的相对选择与生存的需要是密切相关的。本文试图通过研究鲟鱼仔鱼、稚鱼的异速生长证实上述假设。
1材料与方法
1.1试验材料
1.1.1试验鱼本实验的史氏鲟来自于黑龙江下游的自然野生亲鱼,经人工繁殖而得,初孵仔鱼体长在 10.4~11.2 mm。
1.1.2试验池试验在黑龙江省水生动物资源增殖保护站(原黑龙江特产鱼类研究所)桦川渔场进行。用面积为3.14 m2,深度为1.5 m的圆形玻璃钢水槽作为试验池。
1.1.3试验用水为经过臭氧消毒和曝气的地下水,水温为19~23 ℃,pH为8.3,DO为7.9~8.9 mg/L
1.1.4试验饵料试验用饵料为水丝蚓。
1.2试验方法
1.2.1试验鱼的饲养方法在6个圆形玻璃钢槽中,加水至0.7 m,饲养鲟鱼苗。在试验过程中,前期日投饵8次,投饵时间分别为 3:00、6:00 、9:00 、12:00 、15:00 、18:00 、21:00、24:00;后期日投饵6次投饵时间分别为2:00、6:00、10:00、14:00、18:00、22:00。日投饵量根据鲟鱼的生长需要,逐渐增加。在孵出后9 d(21.1~22.3 mm)第一次用活饵投喂每天投饵率为10%,孵出15 d后投饵率降至8%直到试验结束。观察鲟鱼的异常行为和死亡率,每天记录死亡情况,孵出后35 d存活88.2%。
1.2.2样本测定方法试验用鱼苗2万尾,每天随机取样20尾,用麻醉剂(MS-222)麻醉并保存在10%的甲醛溶液中,估测味觉摄食和游动系统发育情况,在40倍的解剖镜下观察体外器官的发育情况,如眼,须,口,鼻,鳃,侧线,鳍和卵黄的吸收情况,确定其发育情况。
体重 :用电子天平测的精确到0.1 mg。
体长 : 用解剖镜目微尺测定,精确到01 mm。
全长 :吻端至尾部末端。
眼径 :是眼球从前至后的直径长度。
口裂宽 :口张后的宽度。
头长 :吻端至鳃盖末端。
尾长 :从泄殖孔至尾末端。
头高 :头部的最高长度。
躯干长 :鳃盖末端至泄殖孔的长度。
胸鳍长 :沿体轴水平线测的其长度。
1.2.3数据的处理方法史氏鲟仔鱼期和稚鱼期的异速生长指数方程直接用全长算得:y=axb,y是依变量,x是自变量,a 是截距,b是生长系数。体长等速生长时b=1,体积等速生长时b=3,当异速正生长时b值大于等速生长值,否则小于等速生长值。令y′=logy,x′=logx,a′=loga,则有y′=a′+bx′。因此对所测数据取对数,然后按最小二乘法进行直线回归,b=SPxy/SSx a′=y′-bx′,列出生长方程。拐点的确定是根据作为全长的x值增长进行分类得回归曲线,计算xmin,xintermediate,xmax,xintermediate在xmin+2到xmax-2之间变动,再用极差检验来验查xmin到xintermediate与xintermediate到xmax之间的生长系数是否显著,在x中值附近表现,将T最大值所对应的x中值定义为拐点[5]。
2结果
2.1早期发育过程
2.1.1仔鱼期在仔鱼期,呼吸系统,游动,味觉和摄食系统的分化和形态发育都很强烈,初孵仔鱼在头底部表面可见孵化腺,口裂和鳃裂还没形成,尾是尾芽状态,躯干和尾被原始的鳍褶分开,尤其在尾部区域,感觉器官未分化,嗅觉器官是只有一个圆形体外开口,眼是一个黑色素点,震动感觉系统,侧线感觉系统只见于头区,在孵出1~2 d后,胸鳍是呈小褶状,口裂开须基可见。在半透明的鳃盖后可见鳃裂原基。其游泳行为是简单的不连续的垂直游动。在孵出后3 d,眼点变黑色,上下唇以小褶状出现在口裂四周,鳍褶开始着色,着色标志着未来鳍条的走向,鳍基在躯干腹侧宽,在尾柄处窄向前,鳍基慢慢伸展发育成将来的背鳍,尾鳍,臀鳍。在孵出后4 d出现了外鳃丝,口与须接触,须基发育增大,与此同时侧线系统达腹部中间背鳍基及鳍条数18~19根出现在背鳍褶中,胸鳍基出现,并分化为窄的皮褶。4 d仔鱼的游泳行为发生了变化,能够在水中水平游动而且具有了趋流性。
在孵出后5~6 d间,覆盖于卵黄囊中后部的血管网变少,几乎消失。胸鳍发育完毕,嗅觉器官分叶,融合成某种神经表皮泡,出现在胸鳍表面上,侧线达到消化管的后部。在孵出的第7 d,口鼻部变得水平,呈腹状,须变长起出吻端,胸鳍条基7~9根、臀鳍条基9~10根出现在鳍褶中。8 d在上下颌分别长出10个犬齿状牙,营外源营养的捕食和感觉器官发育完毕,侧线到达了胸鳍的后缘背鳍臀鳍,从尾鳍和可见的胸鳍基条8~10根分离出来。
2.1.2稚鱼期、幼鱼期孵出的第9 d(21.1~22.3 mm),稚鱼期开始,新的形态特征开始出现,卵黄囊明显消失,多数仔鱼的胃内有水丝蚓,鳍褶中背骨板雏形清晰可见,外鳃丝被外鳃盖覆盖。从11~18 d形态上几乎没什么变化,在11 d在头部背表面和胸鳍区域,可见皱形尖状物。在13 d后,在口鼻部侧背面出现了几行感觉坑。在18~19 d间,象小钉状的侧骨板两行和腹骨板两行,原基开始出现,躯体表面侧面和腹面。在20~35 d之间形态发生了变化,某些幼鱼的新特征出现,背侧,腹骨板,行数发育很大,尾鳍条基19~21根出现并发育成一定的结构,头部尖状物数量增大,口鼻部顶端变的很尖,在34~35 d前臀鳍褶全部消失,此时鲟形态上和成鲟一样。
2.2生长方程
2.2.1异速生长
2.2.1.1眼径眼径的生长可以分两个明显的阶段从孵出到3日龄其生长是正异速b=5.971 4生长方程为y=7E-0.8 x5.9714,见图1。从拐点的14.0 mm体长到35 d的眼径的增长接近等速b=0.981 6生长方程为y=0.039 5 x0.9816,见图2。
2.2.1.2体重体重的异速生长是负相关,在仔鱼期(b=1.152 7)生长方程y=1.193 x1.1527,见图3。从仔鱼形态到稚鱼形态转变是渐进的,主要发生在9 d,从这个拐点的20.4 mm体长到第35 d,稚鱼和幼鱼前期的生长系数无显著不同(P>005)这段期间生长加速,近于等速生长,(b=28575)生长方程为y=0.008 x2.8575,见图4。
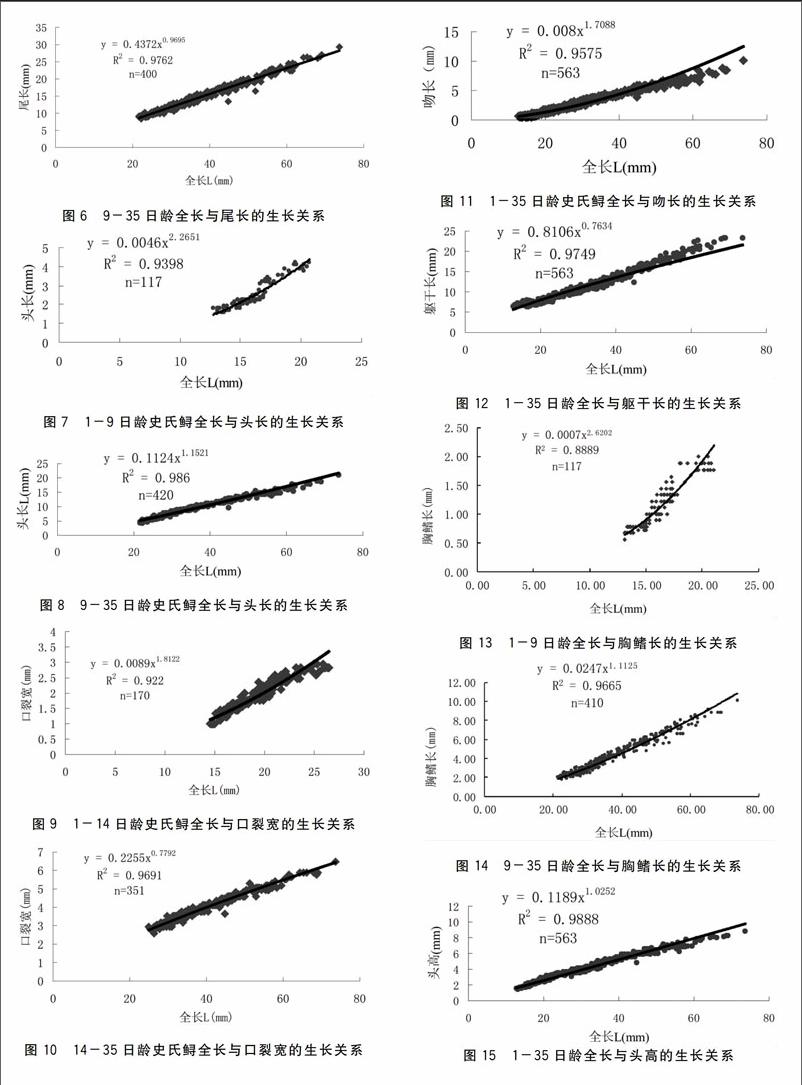
2.2.1.3尾长尾的长度增长是正异速b=1222 2,见图5,直到体长21.1~22.3 mm日龄达9 d,从这个拐点的21.7 mm体长而后生长近于等速b=0.969 5,见图6 。
2.2.1.4头长从孵出到9日龄的仔鲟头长生长正异速(b=2.265 1)生长方程为y=0.004 6 x2.2651,见图7。从拐点的21.5 mm到第35 d异速生长急速下降接近等速生长b=1.1521 生长方程为y=0.112 4 x1.1521,见图8。
2.2.1.5口裂宽在第3 d体长14.5 mm左右时,口裂张开而且生长在26.5 mm体长的第14日龄,生长均是正异速关系,生长方程为y=0008 9 x1.8122,见图9,
2.2.1.8胸鳍长2日龄胸鳍开始发育其生长为正异速b=2.620 2,见图13,直到体长达21.1~22.3 mm 日龄达9 d,从这个拐点的21.5 mm体长到35 d异速生长下
2.2.2等速生长
3讨论
实验结果表明,史氏鲟的早期发育分两个阶段,即胚后期孵出仔鱼(体长10.4~11.2 mm)到开食阶段(体长21.1~22.3 mm)和稚鱼阶段(指外源营养阶段到体态形成阶段)。仔鱼期较稚鱼期和幼鱼期在形态发生和分化上更强烈、更明显。一些形态特征在仔鱼孵出后不久(4 d或9 d)就直接进入异速增长期,而后变成非异速增长。这些形态特征增长的变化与鳃呼吸、外源营养、游泳能力的变化有关系。
在生命的早期阶段,被捕食与饥饿是造成鱼类死亡的主要原因。当史氏鲟仔鱼完成了前期机能的发育后,他们就应该能够逃避敌害生物的捕食。与此同时,仔鱼也应具有了捕获食物的能力,从而增加了鲟鱼生存的机率。数据分析发现在4 d(体长13.5~14.3 mm)前后,仔鱼的头部和尾部就同步进入了异速增长。这就证明了仔鱼同时具有了逃避敌害和捕获食物的能力。
逃避敌害的能力涉及到感觉系统、运动系统、呼吸系统的变化。
实验发现,眼睛直径的在孵出后1 d就开始了正异速增长,到了4 d转变成匀速增长。同样是到了4 d,仔鱼的游泳行为也发生了转变,可以在水中自由游动并且具有了趋流性,但是在夜间仔鱼的趋流性并不表现。逃避敌害首先要能够发现敌害生物的位置,然后要能够主动游泳逃避敌害生物的捕食。仔鱼构造相对简单的视网膜只包括“感绿”视网膜以及视锥细胞(Govardovskii et al.,1991; Dettlaff et al.,1993; Loew & Sillman,1993),这种视网膜及视锥细胞保证了鲟鱼在夜里感知光的色饱和度以及光信号,同时也可辨别有敌害生物存在的水域与安全水域的差异。
游泳行为的变化与胸鳍的发育有直接的关系。鱼类在水中水平游动胸鳍发挥着主要作用。实验结果发现,胸鳍在2日龄开始发育,呈现正异速增长(b=2.620 2),直到9日龄接近匀速增长(b=1.112 5)。
在孵出后的1-4日龄,仔鱼主要依靠发育不完全的鳃、体表呼吸以及覆盖在卵黄囊后部的动脉血管网来维持呼吸。到了5-6日龄,动脉血管网基本消失,皮肤呼吸在呼吸系统中也不再发挥重要作用了。因为这时(4 d)外鳃丝开始发育,到了9日龄鳃盖已经完全覆盖了外鳃丝。
头部长度的增长也是在1-9 d为正异速增长,这与鳃盖的正异速增长是完全吻合的。对于头部长度的迅速增长,有一种解释说:由于卵黄囊储备的营养被耗尽(9 d),稚鱼不得不开始争夺外部食物。因此,他们需要一个高性能的大容量的食物纳入器官。由于在仔鱼孵化阶段头部发育相对不完全,因此头部长度的增长是为了适应不断增大的食物颗粒的需要,同时也是为了更容易追求更大的食物颗粒来加快生长的速度。当然,这种追求更大食物的相对选择也反映出了鲟鱼精力旺盛、身体机能良好。
头部长度与鳃盖的异速增长的这种巧合可以说明:气体交换从皮肤到鳃的转变已经完成。这是由鲟鱼的各个器官得到良好的氧气供应,以及游泳行为和活动模式的转变而引起的。
在9 d前后,史氏鲟鱼苗开始摄食外源营养,在这一时期鱼苗的胃中还有少量的卵黄,肠道前部和中部还有脂肪残留。这表明从9 d到14 d,从鲟鱼开始摄食到内源营养储备被鲟鱼耗尽之前是一个混合营养时期。从1日龄开始至14日龄,口裂宽度就呈正异速增长。这结果也表明了,在这一时期,口裂的宽度的正异速增长与完全转变为捕食外部食物时同时进行的。同时证明,在早期个体发育期间,形态与形态测定的转变伴随着生理上的变化,这也就保证了食物的获得和吸收能够到预期的觅食效果。在混合营养时期,嗅觉、触觉、味觉以及感觉系统的完全发育也证实了上述事实。
最新的结论表明,鱼鳍展开分化为不对称鳍的时期与尾部长度的异速增长变化点相近,然而从仔鱼期开始到35 d尾部增长迅速减慢变为非异速增长。在这一阶段,史氏鲟的行为从根本上发生了变化:幼鱼期的游泳能力比仔鱼期和稚鱼期有了很大进步,鱼的活力也有所增加。这表明尾部长度的匀速增长、分化为不对称鳍以及尾部长度的迅速增长是与鱼游泳能力的改进同步发生的。这就保证了鲟鱼可以分散到更广阔的区域,允许他们拥有更大的觅食区和栖息地来觅食、逃避敌害。鲟鱼身体前部和后部的异速增长优先于身体的躯干部分的发育,也可以解释为一种减少游泳消耗的生理适应。头部和尾部的快速增长对减少身体的阻力是有利的。
鲟鱼与一些硬骨鱼类发育的有一个明显差异。史氏鲟的胸鳍分化比鲤鱼晚,且胸鳍发育变化点与第一次觅食同时发生。对于鲤鱼来说,胸鳍在觅食方面起着很重要的作用,既帮助调整头部的角度来瞄准猎物。然而,对鲟鱼而言,胸鳍的发育变化点与外源营养的开始没有联系,因为鲟鱼是没有视觉,捕食完全依靠触觉和嗅觉。
最后,现在的研究表明在史氏鲟早期发育阶段,重要的形态与形态变化是由于营外源营养、游泳、感觉和呼吸系统的发育引起的。头部与尾部的发育同步异速增长保证了鲟鱼可以在不同时期占据不同的栖息地。这一研究结论同样对应其他现代硬骨鱼类的研究结果,这证明了早期生命阶段的形态发育与增长模式与当前需要有着密切的关系的假设。
参考文献:
[1] 韩骥.黑龙江施氏鲟繁殖群体现状调查[J].水生态学杂志,2012,33(01):144-148
[2] 李文龙.鲟鱼苗中早期死亡原因分析[J].中国水产, 2003(05):50-52
[3] 殷名称,鱼类生态学[M].北京:中国农业出版社,48-52
[4] 石振广,王云山,李文龙.鲟鱼与鲟鱼养殖[M].黑龙江科学技术出版社,2000,03:26-31
[5] 杨运清,王义通,肖艳红. 生物统计学[M].哈尔滨:哈尔滨工程大学出版社,175-176,142-144
(收稿日期:2016-08-11)
