东亚地区栖北散白蚁的遗传分化研究
2016-12-09邓锋刘延胜庞正平平正明王建国
邓锋, 刘延胜, 庞正平, 平正明, 王建国*
(1.江西农业大学农学院,南昌330045;2.常州市武进区白蚁防治所,江苏常州213100;3.广东省昆虫研究所,广州510260)
东亚地区栖北散白蚁的遗传分化研究
邓锋1, 刘延胜1, 庞正平2, 平正明3, 王建国1*
(1.江西农业大学农学院,南昌330045;2.常州市武进区白蚁防治所,江苏常州213100;3.广东省昆虫研究所,广州510260)
本研究通过收集中国的栖北散白蚁Reticulitermessperatus(Kolbe)种类,进行线粒体DNA 细胞色素氧化酶Ⅱ(COⅡ)基因扩增测序,将所得序列与日本和韩国的R.speratusCOⅡ序列进行比较分析,进而比较东亚地区R.speratus种类的亲缘关系和迁徙路线。结果显示中国与韩国、中国与日本分别存在相同的R.speratus亚种,且R.speratus在东亚地区的最有可能迁徙路线是先从中国迁徙到日本,再从日本传播到韩国。
栖北散白蚁;COⅡ;传播路线;亲缘关系
白蚁是生态系统中重要的分解者,影响土壤的结构、稳定性以及有机质的分布和质量,在有机质循环中占有非常重要的地位。但同时,白蚁对木构件和建筑的破坏,也使其成为一种经济害虫。
散白蚁属Reticulitermes由Holmgren于1913年定名,属内共138种,主要分布在东洋区、古北区、新北区等(程冬保,杨兆芬,2014),中国拥有其中大部分的种类,共计111种(黄复生等,2000)。其中,栖北散白蚁Reticulitermessperatus(Kolbe)最早在1885年由Kolbe定名,模式产地为日本,后因属的变动,由Snyder在1949年确定为现在的名字,并沿用至今(黄复生等,2000)。因其地理分布和分类地位,R.speratus在亚洲散白蚁种类中显得尤为有趣。在东亚地区,R.speratus同时分布在中国、日本和韩国。在日本,Morimoto(1968)报道了R.speratus的5个亚种,分别为R.s.speratus、R.s.leptolabralis、R.s.kyushuensis、R.s.okinawanus和R.s.yaeyamanus。但Takematsu和Yamaoka(1999)等认为R.s.okinawanus和R.s.yaeyamanus可用碳氢化合物的方法明显区分开来,但其他3个亚种R.s.speratus、R.s.leptolabralis、R.s.kyushuensis都不能通过形态和化学的方法明显区分开。因此,目前认为日本只存在3个R.speratus亚种。经过Kim等(2012)和Park等(2013)的调查研究,确定韩国只有R.s.kyushuensis1个亚种。在中国,R.speratus并没有进行亚种的细分。另外,R.speratus为中国分布最北的1个白蚁物种,分布在辽宁省丹东市(黄复生等,2000)。李小荣(2012)报道了R.speratus在吉林省公主岭市的分布,从而改写了中国白蚁分布的北界。
R.speratus为东亚广布种,因而其系统发育关系和迁徙路线备受关注(Parketal.,2006;Kimetal.,2012)。Park等(2006)通过分子系统发育分析、古地理学假设等方法,分析了韩、日两国R.speratus的种类以及传播途径,提到其传播可能和中国有关,并提出R.speratus是从中国迁徙到日本,然后再从日本迁徙到韩国的假设。但由于缺乏中国地区的R.speratus种类,其关于R.speratus迁徙路线的假设可能存在问题。因此,本研究通过加入中国地区的R.speratus种类,对R.speratus进行线粒体细胞色素氧化酶Ⅱ(COⅡ)基因扩增,并将所得序列与日、韩已提交NCBI的序列进行比较,对中、日、韩三国R.speratus的系统发育关系进行分析,探讨东亚地区栖北散白蚁的起源和扩散路线,并对其进化机制进行研究。
1 材料和方法
1.1 实验标本信息
所用标本采集于中国辽宁省和山东省的6个市,每个市采集3~5个巢群,其中公主岭市标本为大连市白蚁防治办公室李小荣女士赠送。所有标本分2份保存,1份常温保存于75%乙醇中用于形态鉴定,1份-20 ℃保存于100%乙醇中用于DNA提取。实验随机选择每个市的1~2个地理种群进行DNA提取,标本详细信息见表1。
1.2 形态学鉴定
利用NIKON SMZ1500显微观测设备对标本进行拍照和测量,将获得的测量数据与《中国动物志 昆虫纲 第十七卷 等翅目》(黄复生等,2000)中的原始数据进行比较分析,获得初步的形态鉴定结果,然后将标本和结果送白蚁分类专家平正明老师复核,最终确定种名。
1.3 基因组DNA提取和检测
采用组织基因组DNA提取试剂盒[天根生化科技(北京)有限公司]提取散白蚁基因组DNA,并用琼脂糖凝胶电泳和紫外分光光度法检测DNA的纯度和浓度,-20 ℃保存备用。
1.4 mtDNA序列扩增和测序
为了验证形态鉴定的结果,并进行后续的系统发育分析,我们对线粒体DNA COⅡ基因进行了部分测序。COⅡ基因片段广泛用于欧洲的白蚁研究(Marini & Mantovani,2002;Luchettietal.,2007;Leniaudetal.,2010)。
COⅡ基因片段扩增引物序列为:TL-J-3037(5’-ATGGCAGATTAGTGCAATGG-3’),TK-N-3785(5’-G-TTTTAAGAGACCAGTACTTG-3’)(Xingetal.,2001)。
PCR程序为:94 ℃5 min,45 ℃1 min,65 ℃3 min,35个循环。PCR产物于-20 ℃保存。
PCR产物经1%琼脂糖凝胶电泳检测后,送生工生物工程(上海)股份有限公司进行双向测序。
1.5 数据分析
用DNAStar对所得序列进行拼接和编辑,去除两端引物;然后用MEGA 6.0对序列进行比对,将比对后的序列用DAMBE进行碱基替换饱和性检测,再将适合建树的序列进行分析,计算其碱基组成、核苷酸差异、平均碱基含量等。之后用MEGA基于Kimura 2-parameter模型计算遗传距离,并用最大似然法(ML)和邻接法(NJ)在Kimura 2-parameter模型下构建系统发育树,对系统发育树各进化支进行Bootstrap 1 000次的置信度检验,比较不同方法构建的系统发育树,分析散白蚁的系统发育关系。为了明确中国R.speratus种类的分类地位,下载了GenBank中韩国和日本提交的R.speratus序列用于比较分析(表1)。

表1 散白蚁标本信息

图1 栖北散白蚁兵蚁头部和前胸背板的特征
Fig.1 Characteristics of the soldiers’ pronotum and the head ofReticulitermessperatus
a.头部正面, b.头部背面, c.头部侧面, d.前胸背板。
a.Head, front view, b.Head, dorsal view, c.Head, lateral view, d.pronotum.
2 结果
2.1 形态鉴定结果
经形态数据比对,本研究所用标本为R.speratus种类。图1为兵蚁头部和前胸背板的特征。
2.2R.speratus的DNA序列分析
对样本进行测序分析后,选取长度为655 bp的序列与日、韩两国已提交GenBank的序列一起进行系统发育分析,其中保守位点581个,变异位点74个,简约信息位点62个。A、T、C、G的平均比例分别为39.6%、23.6%、23.3%、13.5%;G+C平均含量为37%,A+T平均含量为63%。个体间的遗传距离详见表2。
序列通过ML和NJ构建系统发育树,所得结果见图2和图3。结果显示日本南部和韩国的R.speratus种类聚为一支,日本北部的R.speratus种类单独聚为一支,中国的R.speratus种类在两支均有分布。
3 讨论
3.1R.speratus在东亚地区的分布及亲缘关系
R.speratus种类为亚洲广布种,中国的辽宁、河北、天津和北京等地都有分布(黄复生等,2000),日本主要分布在吐噶喇海峡以北(Yasudaetal.,2000),而韩国几乎全国都有分布(Kimetal.,2012;Parketal.,2013)。但由于地理生态环境的差异以及扩散因素的影响,各地分布的亚种并不相同。日本R.speratus有3个亚种,其分布因地理而异,北部以R.s.speratus为主,南部因吐噶喇海峡的存在,在吐噶喇海峡以南无R.speratus的分布,可能是吐噶喇海峡阻碍了R.speratus的扩散。韩国的R.speratus种类与日本的R.s.kyushuensis相同。中国的R.speratus并未细分到亚种,但结合遗传距离和系统发育树,对中国的R.speratus进行以下2种分析:第一种分析,从图2和图3中可以看到中国的R.speratus分布在3个大支,可以认为有3个亚种的存在,即中国大部分R.speratus种类与韩国和日本南部的种类聚在一起,构成第一个亚种,第二个亚种是与日本北部R.speratus种类聚在一起的2个种群,第三个亚种是从GenBank中下载的中国R.speratus种类,它们单独成一支存在;第二种分析,从遗传距离来看,前面2个亚种与前一种分析一致,但在GenBank中下载的中国R.speratus序列构成的第三个亚种有待讨论,因为这个亚种与前2个亚种的种内遗传距离大于前2个亚种与其他种类的种间遗传距离,理论上可以认为是2个不同物种,即中国的R.speratus只存在2个亚种,但Austin等(2002)在比较分析北美和欧洲的散白蚁时,发现有的种类因为起源的不同,确实存在种内遗传距离差异很大的情况。综上可知,中国的R.speratus种类至少存在2个亚种,且在日本和韩国都有相同种类分布。
3.2R.speratus在东亚地区的传播路线
R.speratus在中、日、韩三国都有分布,因而其迁徙路线备受关注。通过分析3个国家R.speratus的分子系统发育关系,并结合古地理学假设,我们对R.speratus在东亚地区的传播路线进行了2个假设:第一个假设,在更新世时期,东海陆架盆地可能是在水面之上的,R.speratus通过东海陆架盆地从中国东部迁徙到日本的主要岛屿,然后分布到日本的北部和南部,日本种群再扩散到韩国,形成现在的韩国R.speratus种群(Parketal.,2006)。R.speratus从中国迁徙到日本后,由于第四纪冰川的反复出现,使得迁徙到日本北方的种群经历了多次的寒冷气候,因此,出现了形态变化和遗传变异来适应环境的变化。在韩国,R.speratus的分布纬度与日本南部R.speratus的差不多,均在35°N左右,因而其与日本南部种类遗传差异较小。但是,在进化树中可以看到中国青岛和大连的R.speratus与日本北部的遗传变异也较小,我们推测可能是与青岛和大连的地理位置有关,青岛和大连是中国重要的港口,与外国贸易往来非常多,可能是日本北部的R.speratus通过贸易往来被重新人为地带回了中国。

表2 散白蚁个体间的遗传距离

图2 基于线粒体COⅡ序列构建的栖北散白蚁最大似然树
▲ 本研究中作者采集的中国标本; 下图同。
▲ The specimens were collected in China; the same below.
另外一种假设是,中国R.speratus本身就存在几个亚种,几个亚种都迁徙到日本,然后几个亚种在日本广泛分布,并扩散到韩国。但是,在第四纪冰川时期,部分种类出现了形态变化和遗传变异来适应环境的变化,最终形成了现在的分布情况。如果能增加日本、韩国等地更多的地理种群标本,再针对某个基因在不同地理种群中的表达差异进行分析,可能会得到更好的结果。
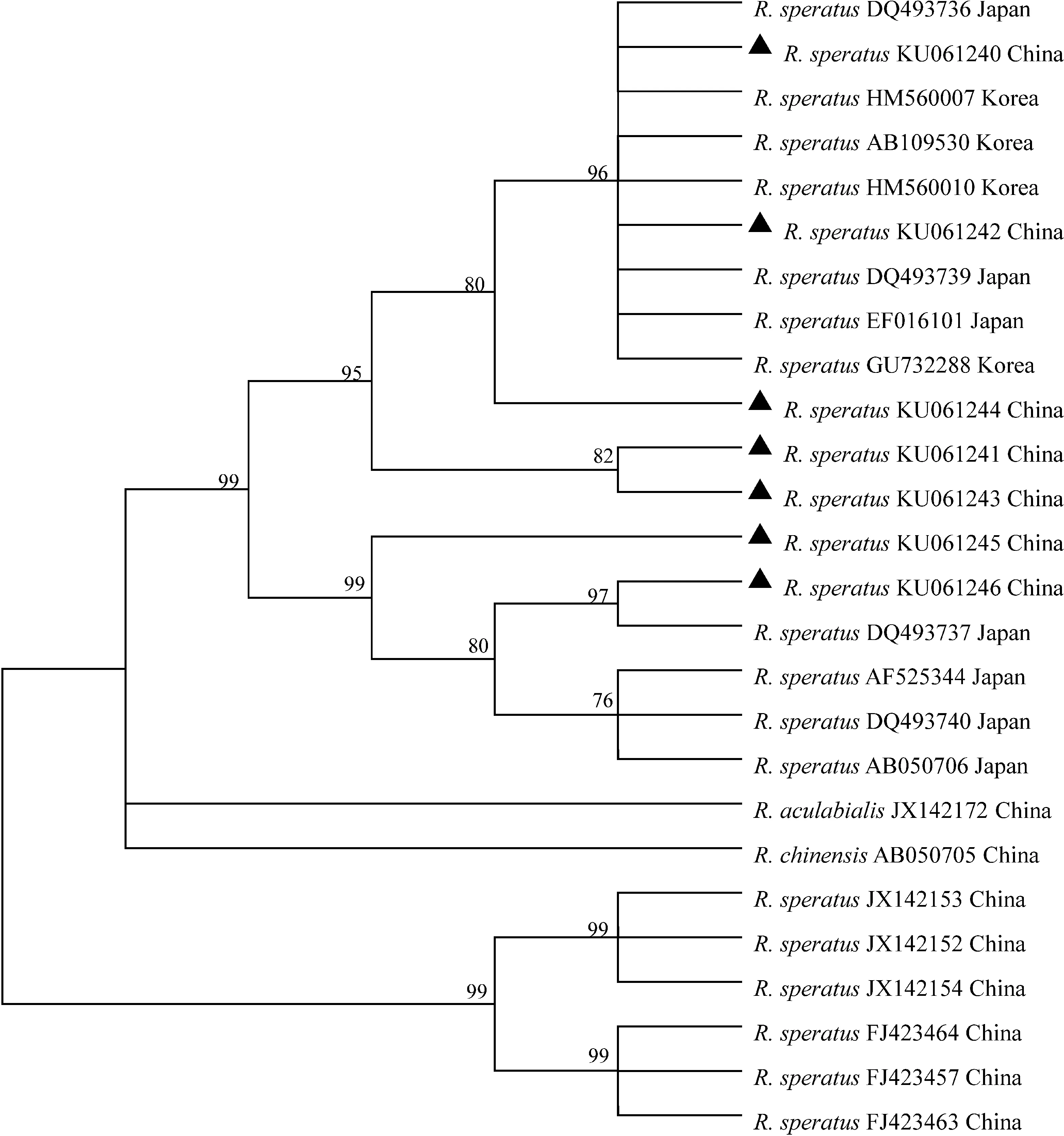
图3 基于线粒体COⅡ序列构建的栖北散白蚁邻接树
Fig.3 Neighbor-joining tree ofReticulitermessperatusbased on mitochondrial COⅡ sequences
致谢:感谢青岛白蚁防治研究所李万红、烟台白蚁防治办公室陈同石、大连白蚁防治办公室宋宝臣和李小荣对北方标本采集给予的极大支持和帮助。
程冬保, 杨兆芬. 2014. 白蚁学[M]. 北京: 科学出版社.
黄复生, 平正明, 何秀松, 等. 2000. 中国动物志 昆虫纲 第十七卷 等翅目[M]. 北京: 科学出版社.
李小荣. 2012. 吉林省公主岭市的白蚁危害改写了我国白蚁分布的北界[J]. 城市害虫防治, (1): 35-37.
Austin JW, Szalanski AL, Uva P,etal. 2002. A comparative genetic analysis of the subterranean termite genusReticulitermes(Isoptera: Rhinotermitidae)[J]. Annals of the Entomological Society of America, 95(6): 753-760.
Kim M, Choi Y, Lee J,etal. 2012. Molecular characteristics of subterranean termites of the genusReticulitermes(Isoptera: Rhinotermitidae) from Korea[J]. Annals of the Entomological Society of America, 105(1): 97-102.
Leniaud L, Dedeine F, Pichon A,etal. 2010. Geographical distribution, genetic diversity and social organization of a new European termite,Reticulitermesurbis(Isoptera: Rhinotermitidae)[J]. Biological Invasions, 12(5): 1389-1402.
Long YH, Xiang H, Xie L,etal. 2009. Intra- and interspecific analysis of genetic diversity and phylogeny of termites (Isoptera) in east China detected by ISSR and COⅡ markers[J]. Sociobiology, 53(2A&2B): 411-430.
Luchetti A, Marini M, Mantovani B. 2007. Filling the European gap: biosystematics of the eusocial systemReticulitermes(Isoptera, Rhinotermitidae) in the Balkanic Peninsula and Aegean area[J]. Molecular Phylogenetics & Evolution, 45(1): 377-383.
Marini M, Mantovani B. 2002. Molecular relationships among European samples ofReticulitermes(Isoptera, Rhinotermitidae)[J]. Molecular Phylogenetics & Evolution, 22(3): 454-459.
Morimoto K. 1968. Termites of the genusReticulitermesof Japan and Taiwan[J]. Bulletin of Government Forest Experiment Station, 217: 43-73.
Ohkuma M, Yuzawa H, Amornsak W,etal. 2004. Molecular phylogeny of Asian termites (Isoptera) of the families Termitidae and Rhinotermitidae based on mitochondrial COⅡ sequences[J]. Molecular Phylogenetics & Evolution, 31(2): 701-710.
Park HJ, Bae CH, Lee JJ,etal. 2013. A comparative phylogenetic analysis ofReticulitermessperatuskyushuensis(Isoptera: Rhinotermitidae) on Jeju Island, Korea[J]. Journal of Agriculture & Life Sciences, 47(1): 137-146.
Park YC, Kitade O, Schwarz M,etal. 2006. Intraspecific molecular phylogeny, genetic variation and phylogeography ofReticulitermessperatus(Isoptera: Rhinotermitidae)[J]. Molecules and Cells, 21(1): 89-103.
Takematsu Y, Yamaoka R. 1999. Cuticular hydrocarbons ofReticulitermes(Isoptera: Rhinotermitidae) in Japan and neighboring countries as chemotaxonomic characters[J]. Applied Entomology and Zoology, 34(1): 179-188.
Xing LX, Maekawa K, Miura T,etal. 2001. A reexamination on the taxonomic position of ChineseHeterotermesaculabialis(Isoptera: Rhinotermitidae) based on the mitochondrial cytochrome oxidase Ⅱ gene[J]. Entomological Science, 4(1): 53-58.
Yashiro T, Matsuura K. 2007. Distribution and phylogenetic analysis of termite egg-mimicking fungi “termite balls” inReticulitermestermites[J]. Annals of the Entomological Society of America, 100(4): 532-538.
Yasuda I, Nakasone Y, Kinjo K,etal. 2000. Morphology and distribution of termites in Ryukyu islands and north and south Daito islands[J]. Japanese Journal of Entomology (New Series), 3(4): 139-156.
Genetic Differentiation ofReticulitermessperatusin East Asia
DENG Feng1, LIU Yansheng1, PANG Zhengping2, PING Zhengming3, WANG Jianguo1*
(1.School of Agricultural Sciences, Jiangxi Agricultural University, Nanchang 330045, China;2.Changzhou Wujing Institute of Termites Control, Changzhou, Jiangsu Province 213100, China;3.Guangdong Entomological Institute, Guangzhou 510260, China)
In this study,Reticulitermessperatus(Kolbe) were collected in China and a portion of the mitochondrial gene encoding for the cytochrome oxidase subunit Ⅱ (COⅡ) was sequenced.In order to compare the phylogenetic relationship and migration route ofR.speratusin east Asia, GenBank data of Korean populations, southern and northern Japanese populations were used.According to the comparative analysis of the sequences, it was suggested that the sameR.speratussubspecies existed in China and Japan, China and Korea, respectively, andR.speratusmigrated into the Japanese main islands from east China, and Korean populations diverged from Japanese populations.
Reticulitermessperatus; COⅡ; migration route; phylogenetic relationship
2015-10-12 接受日期:2016-01-30 基金项目:江西省研究生创新专项资金项目(YC2014-S180); 国家自然科学基金项目(31160380,30960223); 江苏省常州市武进区白蚁所横向项目(WJ 20130418)
邓锋(1991—), 男, 硕士研究生, 主要从事白蚁系统发育研究, E-mail:dfeng0827@163.com
*通信作者Corresponding author,E-mail:ppdjxau@gmail.com
10.11984/j.issn.1000-7083.20150313
Q951; Q969
A
1000-7083(2016)03-0361-07
