青藏高原两种雪雀惊飞距离的比较研究
2016-12-09张贺李文靖胡延萍张堰铭
张贺, 李文靖, 胡延萍, 张堰铭*
(1.中国科学院西北高原生物研究所,高原生物适应与进化重点实验室,西宁810008;2.中国科学院大学,北京100049)
青藏高原两种雪雀惊飞距离的比较研究
张贺1, 2, 李文靖1, 2, 胡延萍1, 张堰铭1*
(1.中国科学院西北高原生物研究所,高原生物适应与进化重点实验室,西宁810008;2.中国科学院大学,北京100049)
在分析鸟类对人类干扰耐受程度的研究中,惊飞距离是常用的衡量指标之一,它是指人在鸟类惊飞之前能接近鸟类的距离,反映了鸟类对人为侵扰的适应程度。本文通过对青藏高原广泛分布的白腰雪雀Montifringillataczanowskii和棕颈雪雀M.ruficollis惊飞距离的比较研究,旨在探讨2种鸟类对人类干扰适应性的差异。结果显示,体型较大的白腰雪雀的惊飞距离(12.50 m ± 0.36 m,n=46)显著大于棕颈雪雀的惊飞距离(11.03 m±0.43 m,n=33);觅食个体的惊飞距离显著小于观望个体(P<0.001);随着入侵者起始距离的增加,2种雪雀惊飞距离逐渐增大(P<0.001)。结果表明,鸟类初始状态和入侵者起始距离能够显著影响鸟类的惊飞距离,体型较大的鸟类对人类干扰的适应性较差。
惊飞距离;人类干扰;白腰雪雀;棕颈雪雀;青藏高原
随着人类活动范围和程度的扩大,人为干扰已经成为鸟类生存面临的主要威胁(Heiletal.,2007;Mller,2008)。惊飞距离是衡量动物对人类干扰耐受程度的重要指标,它是指人在鸟类惊飞之前能接近鸟类的距离(Ydenberg & Dill,1986;Cooper & Frederick,2007)。大量综述文献及荟萃分析(meta-analyses)结果表明:入侵干扰者特征(如起始距离、接近速度、靠近方向等)、目标动物特性(如体型形态、个性特征、年龄经验等)以及外部环境条件(如栖息地特征、与隐蔽处距离等)均能显著影响鸟类的惊飞距离(Stankowich & Blumstein,2005;Stankowich,2008;Samiaetal.,2016)。
雪雀属Montifringilla是典型的高原鸟类,主要分布于我国青藏高原及其毗邻地区(屈延华等,2002)。其中白腰雪雀M.taczanowskii和棕颈雪雀M.ruficollis共同分布于海拔3 000 m以上,有些甚至可达5 500 m,是青藏高原的特有种(Leietal.,2003)。2种雪雀均在高原鼠兔Ochotonacurzoniae洞穴中营巢,形成了奇特的“鸟鼠同穴”现象。由于白腰雪雀体型明显大于棕颈雪雀,在种间竞争中具有优势,因此白腰雪雀常常占据食物资源更为有利的地形(Zeng & Lu,2009)。虽然有研究曾调查过青藏公路及青藏铁路附近人类干扰活动对3种雪雀惊飞距离的影响(Geetal.,2011),但并未考虑人为侵扰的起始距离和动物的原始状态对惊飞距离的影响,也没有深入讨论雪雀对人类干扰的适应性问题。
野生动物的惊飞距离研究在科学管理野生动物旅游业,促进野生动物保护等方面具有重要的实践意义。本文旨在研究同域共存的2种雪雀惊飞距离的差异及其对不同因素的响应,为青藏高原小型地栖型鸟类的管理、保护及相关政策的制定提供科学依据。
1 材料与方法
1.1 研究区域概况

1.2 野外调查方法
野外实验选择晴朗无风的日子,时间为08∶00—18∶00。选择1只目标雪雀并记录其原始行为(觅食或者观望),然后以恒定的步速和步幅(0.5 m/步)接近目标个体,并注意目标动物的行为变化。当雪雀逃跑或者飞离原始位置时,实验者迅速记录此时步数,并以此为起点继续按原速步行至雪雀离开前所在的位置,再记录此时步数。第二次记录的步数距离即为该雪雀的惊飞距离,2次记录的距离之和即为人为侵扰的起始距离。
为避免伪重复,每次实验后实验者离开样地100 m之外记录数据,并准备下一次实验,以排除重复测量同一个体及其他雪雀个体注意到实验过程从而影响数据的真实性。每2次实验间隔3 d以避免动物的适应性。所有实验均由同一人穿着同一件衣服完成。
1.3 数据统计分析
为分析入侵者起始距离、动物原始状态对2种雪雀惊飞距离的影响,采用一般线性模型(general linear model,GLM)对数据进行分析。惊飞距离为因变量,原始状态(觅食vs.观望)和物种(白腰雪雀vs.棕颈雪雀)为固定变量,起始距离作为协变量。如果差异有统计学意义,采用Bonferroni进行多重比较。
数据处理与分析由SPSS 19.0完成,显著水平为0.05,数值以平均值±标准误表示。
2 结果
实验期间,共获得白腰雪雀有效数据46组,棕颈雪雀有效数据33组。一般线性模型分析表明,白腰雪雀的惊飞距离(12.50 m±0.36 m)极显著大于棕颈雪雀(11.03 m±0.43 m)(F=6.792,P=0.011;表1,图1)。
雪雀的惊飞距离受到其初始状态的影响,觅食个体惊飞距离(10.66 m±0.42 m)显著小于观望个体(12.87 m±0.38 m)(F=94.860,P<0.001;表1,图1)。初始状态与物种的交互作用对惊飞距离影响的差异无统计学意义(F=2.850,P=0.096)。
入侵者起始距离对雪雀惊飞距离影响的差异有高度统计学意义(F=94.860,P<0.001;表1)。随着起始距离的增加,雪雀惊飞距离也随之增大(图2)。回归系数分别为白腰雪雀r=0.66,棕颈雪雀r=0.81。

表1 白腰雪雀和棕颈雪雀惊飞距离一般线性模型分析结果
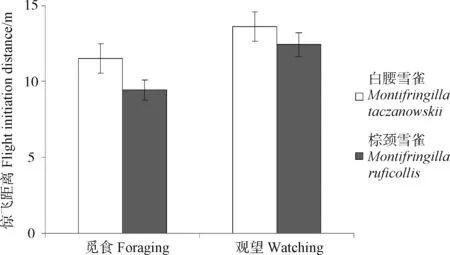
图1 白腰雪雀和棕颈雪雀在不同状态下的惊飞距离
Fig.1 The flight initiation distance ofMontifringillataczanowskii

图2 入侵者起始距离与白腰雪雀和棕颈雪雀惊飞距离的关系
3 讨论
对2种雪雀惊飞距离的分析结果表明,起始距离和原始状态能够显著影响惊飞距离。随着起始距离的增加,惊飞距离逐渐增大。“尽快逃走避免慌张”(FEAR)假说认为动物观察到入侵者后会立即逃走,以降低持续侦查的能量消耗(Blumstein,2010)。Dumont等(2012)质疑起始距离与惊飞距离的关系,认为二者之间存在的只是数学关系而非生物学效应。随后Chamaillé-Jammes和Blumstein(2012)采用稳健方法论证明了二者之间不可能存在数学关系。Samia等(2013)整合了鸟类、哺乳类和两栖类动物的大量数据,采用荟萃分析证明了二者之间存在的生物学效应以及FEAR假说的真实性。本研究也完全支持该假说并证实了起始距离对惊飞距离影响的正效应。
本研究结果表明觅食个体的惊飞距离小于观望个体,这与前人研究结果类似(Blumsteinetal.,2004)。一方面,根据最优逃避理论(optimal escape theory)(Ydenberg & Dill,1986;Cooper & Frederick,2007),动物面临风险时需要在逃跑或留在原地之间进行不断权衡。当动物在觅食过程中受到入侵者干扰,离开觅食地点意味着本次获取食物的机会丧失,并且再次回到觅食点摄食又需要消耗时间与能量。因此,觅食个体能够对人为侵扰有较高的容忍度。另外,已有的研究表明,动物在觅食过程中会减少对环境风险的监视(Dukas & Kamil,2000),当动物处于觅食、玩耍或打斗时,对风险源的注意放松会导致惊飞距离降低(Krause & Godin,1996;Blumstein,1998;Brick,1998)。
白腰雪雀的惊飞距离显著大于棕颈雪雀,这与Ge等(2011)在青藏铁路、青藏公路附近的调查结果一致。体型可能影响鸟类的惊飞距离,即体型较大的鸟类,惊飞距离也较大(王彦平等,2004;Mller & Erritze,2010)。鸟类眼睛的大小与体型成正比,体型较大的个体眼眶也较大,能够获得更好的视野。同时,大型个体的视线也不易被遮蔽物所遮拦,因此往往能够提前发现入侵者,躲避捕食风险。
研究鸟类对人为侵扰容忍度的影响不仅有助于防止或减少人为侵扰对鸟类的影响,还可为极易受干扰的物种保护提供理论依据,对于维持和提高鸟类多样性也有重要意义(王彦平等,2004)。特别是在青藏高原地区,逐渐增加的人类活动对野生动物带来了巨大的影响。本文的研究结果提示该地区应减少生态旅游等人为干扰,并加大环境教育力度。在加强野生动物的监测和研究中,即使是同域分布的近缘物种,也要针对各自的生物学特性制定不同的保护管理策略。而未来研究的重点将集中于集群效应对于野生动物惊飞距离的影响,特别是由异种个体组成的混合群对人类干扰的适应研究,这对于野生动物的管理和保护更有意义(Westonetal.,2012)。
屈延华, 雷富民, 尹祚华. 2002. 雪雀属鸟类栖息地在中国的分布[J]. 动物学报, 48(4): 471-479.
曲家鹏, 杨敏, 李文靖,等. 2008. 高原鼠兔家群结构的季节变异[J]. 兽类学报, 28(2): 144-150.
王彦平, 陈水华, 丁平. 2004. 惊飞距离——杭州常见鸟类对人为侵扰的适应性[J]. 动物学研究, 25(3): 214-220.
王长庭, 王启基, 龙瑞军, 等. 2004. 高寒草甸群落植物多样性和初级生产力沿海拔梯度变化的研究[J]. 植物生态学报, 28(2): 240-245.
Blumstein DT, Runyan A, Seymour M,etal. 2004. Locomotor ability and wariness in yellow-bellied marmots[J]. Ethology, 110(6): 615-634.
Blumstein DT. 1998. Quantifying predation risk for refuging animals: a case study with golden marmots[J]. Ethology, 104(6): 501-516.
Blumstein DT. 2010. Flush early and avoid the rush: a general rule of antipredator behavior?[J]. Behavioral Ecology, 21(3): 440-442.
Brick O. 1998. Fighting behaviour, vigilance and predation risk in the cichlid fish (Nannacaraanomala)[J]. Animal Behaviour, 56(2): 309-317.
Chamaillé-Jammes S, Blumstein DT. 2012. A case for quantile regression in behavioral ecology: getting more out of flight initiation distance data[J]. Behavioral Ecology and Sociobiology, 66(6): 985-992.
Cooper WE, Frederick WG. 2007. Optimal flight initiation distance[J]. Journal of Theoretical Biology, 244(1): 59-67.
Dukas R, Kamil AC. 2000. The cost of limited attention in blue jays[J]. Behavioral Ecology, 11(5): 502-506.
Dumont F, Pasquaretta C, Réale D,etal. 2012. Flight initiation distance and starting distance: biological effect or mathematical artefact?[J]. Ethology, 118(11): 1051-1062.
Ge C, Li ZQ, Li J,etal. 2011. The effects on birds of human encroachment on the Qinghai-Tibet Plateau[J]. Transportation Research Part D: Transport and Environment, 16(8): 604-606.
Heil L, Fernández-Juricic E, Renison D,etal. 2007. Avian responses to tourism in the biogeographically isolated high Córdoba Mountains, Argentina[J]. Biodiversity & Conservation, 16(4): 1009-1026.
Krause J, Godin J-GJ. 1996. Influence of prey foraging posture on flight behavior and predation risk: predators take advantage of unwary prey[J]. Behavioral Ecology, 7(3): 264-271.
Lei FM, Qu YH, Lu JL,etal. 2003. Conservation on diversity and distribution patterns of endemic birds in China[J]. Biodiversity & Conservation, 12(2): 239-254.
Samia DS, Blumstein DT, Stankowich T,etal. 2016. Fifty years of chasing lizards: new insights advance optimal escape theory[J]. Biological Reviews of the Cambridge Philosophical Society, 91(2): 349-366.
Samia DS, Nomura F, Blumstein DT. 2013. Do animals generally flush early and avoid the rush? A meta-analysis[J]. Biology Letters, 9(2): 20130016.
Stankowich T, Blumstein DT. 2005. Fear in animals: a meta-analysis and review of risk assessment[J]. Proceedings of the Royal Society B: Biological Sciences, 272(1581): 2627-2634.
Stankowich T. 2008. Ungulate flight responses to human disturbance: a review and meta-analysis[J]. Biological Conservation, 141(9): 2159-2173.
Weston M, McLeod EM, Blumstein D,etal. 2012. A review of flight-initiation distances and their application to managing disturbance to Australian birds[J]. Emu: Austral Ornithology, 112(4): 269-286.
Ydenberg RC, Dill LM. 1986. The economics of fleeing from predators[J]. Advances in the Study of Behavior, 16(C): 229-249.
Zeng XH, Lu X. 2009. Interspecific dominance and asymmetric competition with respect to nesting habitats between two snowfinch species in a high-altitude extreme environment[J]. Ecological Research, 24(3): 607-616.
Research on the Flight Initiation Distance of Two Snowfinch Species on the Qinghai-Tibetan Plateau
ZHANG He1, 2, LI Wenjing1, 2, HU Yanping1, ZHANG Yanming1*
(1.Key Laboratory of Adaptation and Evolution of Plateau Biota, Northwest Institute of Plateau Biology, Chinese Academy of Sciences, Xining 810008, China; 2.University of the Chinese Academy of Sciences, Beijing 100049, China)
Flight initiation distance (FID) is an excellent metric to quantify the tolerance degree of bird against human intrusion.It refers to the distance of a bird begins to flee from an approaching threat, and reflects the adaptation ability to human intrusion.The present work investigated the FID of the white-rumped snowfinch (Montifringillataczanowskii) and rufous-necked snowfinch (M.ruficollis) on the Qinghai-Tibetan Plateau, and discussed the adaption of these two species to the human disturbance.The results indicated that the FID ofM.taczanowskii(12.50 m ± 0.36 m,n=46), which was the largest species among its genus, was significantly larger than that ofM.ruficollis(11.03 m±0.43 m,n=33).Foraging snowfinch tolerated a closer distance than watching individuals, and there was a significant positive relation between the starting distance of intruder and the FID of both species.In conclusion, the initial activity of animals and the starting distance of intruder can influence the FIDs, and the birds with larger body size had relatively lower adaption ability against human intrusions.
flight initiation distance; human intrusion;Montifringillataczanowskii;Montifringillaruficollis; Qinghai-Tibetan Plateau
2015-11-24 接受日期:2016-03-02
国家自然科学基金项目(31270467); 中国科学院青年创新促进会基金项目(2014386)
张贺(1989—), 男, 博士研究生, 研究方向: 动物生态学, E-mail:zhanghe072@163.com
*通信作者Corresponding author,E-mail:ymzhangnwipb@126.com
10.11984/j.issn.1000-7083.20150372
Q959.7; Q958.1
A
1000-7083(2016)03-0368-04
