13个铁线莲品种的核型比较分析
2016-12-09任佳伦季梦成刘志高
任佳伦,季梦成,赵 爽,刘志高
(浙江农林大学 风景园林与建筑学院,浙江 临安 311300)
13个铁线莲品种的核型比较分析
任佳伦,季梦成,赵 爽,刘志高
(浙江农林大学 风景园林与建筑学院,浙江 临安 311300)
铁线莲属Clematis为观赏价值极高的藤本植物。铁线莲品种数量繁多,形态变异丰富,遗传背景也相对复杂,品种间存在杂交不亲和性的障碍,对杂交亲本的染色体核型分析等细胞学特征的研究显得十分重要。采用酶解法-去壁低渗火焰干燥法与Giemsa-C分带染色技术,以13个铁线莲品种自然生长的幼嫩根尖为试材,进行染色体参数和核型分析。结果表明:13个品种的染色体均为2n=2x=16,均为二倍体;染色体核型分为2种类型:2n= 2x=16=10m+4st+2t,2n=2x=16=10m+6st;随体均在第6条,第7条或第8条染色体上,且位置均在短臂上;13个品种的核型类型均为 “2A”,具有很大的相似性,说明杂交不亲和现象由其他因素导致。图3表3参18
植物细胞遗传学;铁线莲属;染色体;核型
1 材料与方法
1.1 试验材料
13个铁线莲品种均取自浙江农林大学铁线莲种质资源圃(表1),所取试材均为自然生长状态下幼嫩根尖。
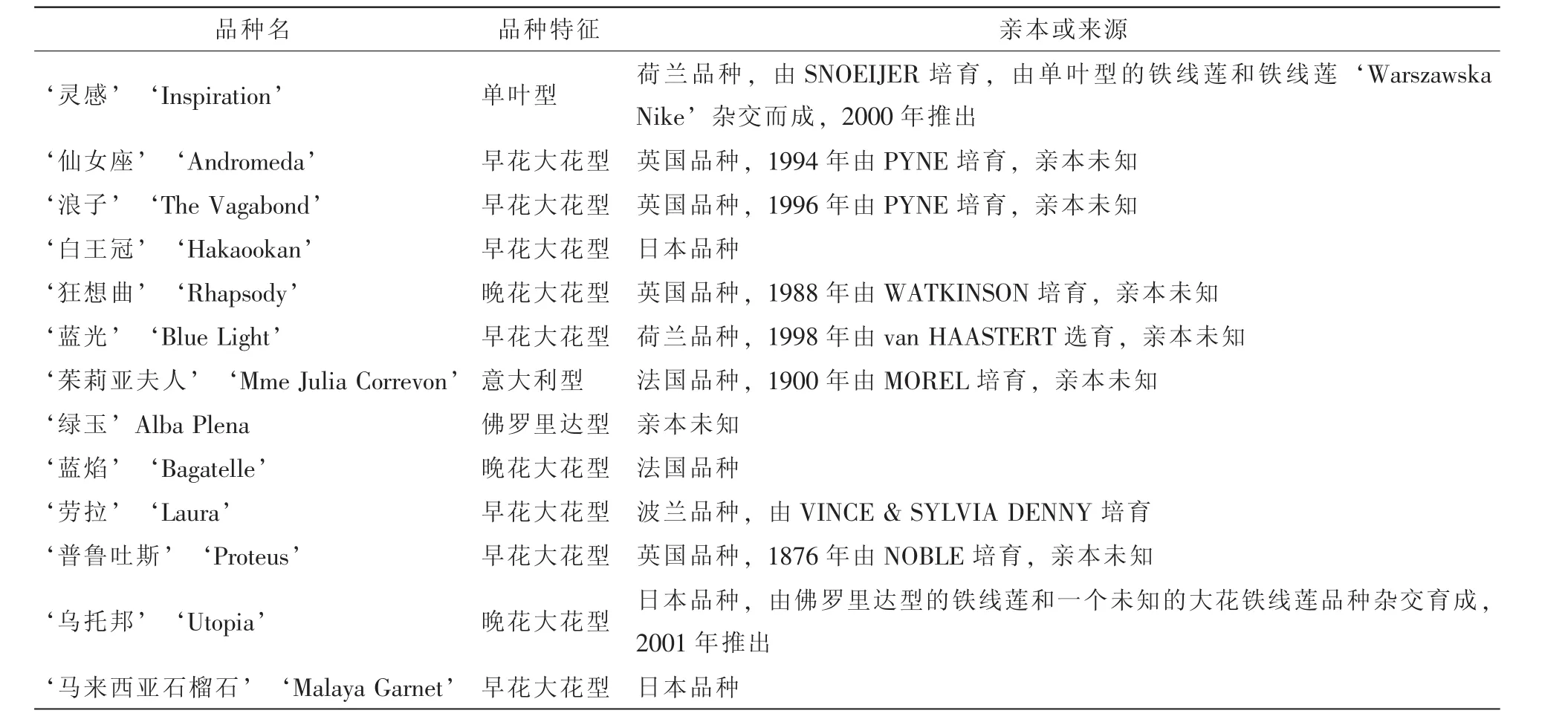
表1 13个铁线莲品种的基本信息Table 1 Information of thirteen Clematis cultivars
1.2 试验方法
1.2.1 染色体制片 采用酶解法与去壁低渗火焰干燥法制片[8]。将材料放入0.002 mol·L-18-羟基喹啉溶液中预处理4.0~4.5 h,预处理后以0.075 mol·L-1氯化钾溶液进行前低渗处理,0.5 h后在V(甲酸)∶V(冰乙酸)=3∶1的根尖固定液下固定数小时。再以体积分数为2.5%纤维素酶和2.5%果胶酶混合液对根尖进行4.0~5.0 h的酶解去壁,酶解完全后去除上清液和杂质,再固定0.5 h后用滴管将此黏稠溶液滴于预先在乙醇溶液中冷冻-20℃的载玻片上,用乙醇灯火焰微热烘干,即得到含有根尖细胞的玻片。
1.2.2 染色 采用染色体Giemsa-C分带技术[9]进行染色。将0.2 mol·L-1氯化氢和2×柠檬酸钠(SSC)缓冲液分别放入60℃恒温水浴锅中加热,将制好的载玻片放入0.2 mol·L-1氯化氢中处理160 s。之后将玻片放入水中洗去残留的氯化氢,再放入氢氧化钡溶液中处理420~430 s,后转入水龙头下将氢氧化钡溶液冲洗掉。最后将玻片放入2×SSC水浴锅中加热,1.0 h后放入20.0 mL磷酸氢二钠+10.0 mL磷酸二氢钾+50~60滴Giemsa染液的混合溶液中染色数小时。
1.2.3 镜检 染色后用蒸馏水轻轻冲洗玻片,空气中干燥后即可进行染色体观测和拍照。先在40倍物镜下找到分裂较好的染色体,向载玻片滴2滴二甲苯,盖玻片滴2滴镜油,再转至100倍物镜下拍照。
1.2.4 参数计算及制作核型模式图 统计细胞分裂相30个·品种-1,其中85%以上的分裂相中染色体一致的数目定为该品种的染色体数[10]。分析时,各个品种取1个染色体形态好且分散好的分裂相,染色体相对长度的计算按照LEVAN等[11]的方法。核型类型以染色体长度比和臂比值来衡量,方法参照STEBINNS[12]的分类标准。着丝点位置的命名按照LEVAN等[11]的方法。核不对称系数(As.K.,%)的计算按照ARANO[13]的方法。染色体相对长度系数(I.R.L)的分类参照KUO等[14]的方法。以Adobe Photoshop CS测量染色体的长短臂与随体的相对长度[15],再以Excel计算染色体的相对长度[16],臂比值,臂比均值(AAR),臂比值大于2的染色体比例(PCA),臂指数(N.F值),着丝点指数等,并制作核型模式图。
2 结果与分析
经观察发现:13个铁线莲品种均为二倍体,染色体数为2n=2x=16,染色体基数为8,且品种间的核型公式极为相似,核型类型均为2A。13个铁线莲品种的核型参数见表2。由表2可知:核型公式‘灵感’为2n=2x=16=10m+4st+2t(2SAT), ‘仙女座’为2n=2x=16=10m+6st(4SAT), ‘浪子’为2n=2x=16=10m+4st(2SAT)+2t, ‘白王冠’为2n=2x=16=10m+4st+2t(2SAT), ‘狂想曲’为2n=2x=16=10m+6st(2SAT), ‘蓝光’为2n=2x=16=10m+4st+2t(2SAT), ‘茱莉亚夫人’为2n=2x=16=10m+6st(2SAT),‘绿玉’为2n=2x=16=10m+4st+2t(2SAT), ‘蓝焰’为2n=2x=16=10m+6st(2SAT), ‘劳拉’为2n=2x=16=10m+6st(2SAT), ‘普鲁吐斯’为2n=2x=16=10m+4st+2t(2SAT), ‘乌托邦’为2n=2x=16=10m+6st(2SAT), ‘马来西亚石榴石’为2n=2x=10m+2st+4t(4SAT)。
13个品种的臂指数均为26,臂比值大于2的染色体比例均为37.50%。由图1~图3可知, ‘灵感’‘白王冠’ ‘狂想曲’ ‘绿玉’ ‘蓝焰’在第8对染色体上具有随体, ‘仙女座’ ‘马来西亚石榴石’于第6对与第8对染色体上具有随体, ‘浪子’ ‘茱莉亚夫人’ ‘普鲁吐斯’ ‘乌托邦’于第7对染色
体上具有随体, ‘蓝光’ ‘劳拉’在第6对染色体上具有随体。其他染色体组的核型参数比较见表3。

表2 13个铁线莲品种的染色体参数Table 2 Chromosomes parameters of thirteen Clematis cultivars

表2 (续)Table 2 Continued
3 讨论
核型分析是细胞分类学的一个重要指标,能够识别染色体的形态特征,对研究系统的演化、种间亲缘关系及分类有重要的意义[17]。本研究报道了13个铁线莲栽培品种的染色体数目与核型,均为首次详细报道。13个品种均为二倍体(2n=2x=16),染色体基数为8,说明铁线莲品种的染色体在数目上是比较恒定的,这与彭绿春等[7]的研究结果一致,这证明铁线莲属的染色体基数比较恒定的表现为x=8。本试验中13个铁线莲品种的核型类型均为 “2A”,这与张镱锂等[2,4]的研究结果有所不同。
13个品种的臂指数值均为26,核型结构为:第1~5对均为具中部着丝点(m)染色体,第6对为均为具近端(st)着丝点染色体,第7至8对为具近端(st)着丝点染色体或端部(t)着丝点染色体,这说明铁线莲品种的核型基本模式极为一致,这与张镱锂等[4]研究一致。但这13个品种所包含的m,st,t这4类染色体的数量却不尽相同,从核型模式图中可明显看出这种差异。核型分析结果表明,13个铁线莲品种都能观察到随体染色体,除 ‘仙女座’和 ‘马来西亚石榴石’具有2对随体外,其他品种均具有1对随体,且位置都在短臂上,这与前人的研究结果也一致[2-4]。在被子植物中,核型进化的基本趋势是由对称向不对称方向发展的,系统演化上处于比较古老或原始的植物往往具有较对称的核型,不对称的核型通常出现在较进化或特化的植物中[12]。本研究中13个铁线莲品种的核不对称系数均相对较低(61.78%~64.78%),变异很小,其按进化程度由高到低的顺序为 ‘绿玉’ ‘茱莉亚夫人’ ‘浪子’ ‘劳拉’ ‘乌托邦’ ‘普鲁吐斯’ ‘马来西亚石榴石’ ‘狂想曲’ ‘蓝光’ ‘蓝焰’ ‘仙女座’ ‘灵感’‘白王冠’。此结果与杨光穗等人的研究结果基本一致[18]。
本研究13个铁线莲品种的核型特征极为相似,说明杂交不亲和可能是温度、湿度或人为等其他因素造成,有待于进一步探究。13个品种间的核型参数间存在的差异,可为铁线莲杂交育种提供细胞学资料。本研究中各染色体显带都不明显,因此,未将染色体Giemsa-C分带技术应用到铁线莲品种的核
型分析中。

图1 13个铁线莲品种的染色体中期分裂相(×1 000)Figure 1 Metaphase cleavage phase of chromosomes of thirteen Clematis cultivars(×1 000)

表3 13个铁线莲品种染色体参数比较Table 3 Chromosomes parameters of thirteen Clematis cultivars

图2 13个铁线莲品种的染色体核型图Figure 2 Karyotype of thirteen Clematis cultivars

图3 13个铁线莲品种的核型模式图Figure 3 Chromosomes karyotype mode of thirteen Clematis cultivars
[1] 蔡艳飞,李世峰,李涵,等.中国铁线莲属植物研究进展[J].中国农学通报,2009,25(4):195-198.
CAI Yanfei,LI Shifeng,LI Han,et al.Advance in the research of Climatis L.[J].Chin Agric Sci Bull,2009,25(4):
195-198.
[2] 张镱锂,贺士元.国产6种铁线莲的染色体研究[J].武汉植物学研究,1990,8(2):115-121.
ZHANG Yili,HE Shiyuan.Chromosome studies on 6 species of Clematis in China[J].J Wuhan Bot Res,1990,8(2): 115-121.
[3] 龚维忠,龙雅宜,李懋学.北京地区铁线莲属植物的研究[J].武汉植物学研究,1985,3(4):371-379.
GONG Weizhong,LONG Yayi,LI Maoxue.Karyotype studies on Clematis from Beijing,China[J].J Wuhan Bot Res, 1985,3(4):371-379.
[4] 张镱锂.7种铁线莲的染色体研究[J].武汉植物学研究,1991,9(2):107-111.
ZHANG Yili.Chromosome studies on 7 species of Clematis in China[J].J Wuhan Bot Res,1991,9(2):107-111.
[5] 杨亲二,罗毅波,洪德元.湖南7种毛茛科植物的核型研究[J].广西植物,1994,14(1):27-36.
YANG Qinger,LUO Yibo,HONG Deyuan.A karyotype study of six species in the Ranunculaceae from Hunan,China[J].Guihaia,1994,14(1):27-36.
[6] 杨亲二.国产毛茛科银莲花族17种植物的细胞学研究[J].植物分类学报,2002,40(5):396-405.
YANG Qinger.Cytology of ten species in Anemone,one in Anemoclema and six in Clematis(Trib.Anemoneae,Ranunculaceae)from China[J].Acta Phytotaxon Sin,2002,40(5):396-405.
[7] 彭绿春,于恒隽,余娜,等.10种铁线莲的核型特征及核型似近系数聚类分析[J].湖南农业大学学报:自然科学版,2012,38(6):617-622.
PENG Lüchun,YU Hengjuan,YU Na,et al.Karyotype features of 10 species of Clematis and cluster analysis of karyotypies resemblance-near coefficient[J].J Hunan Agric Univ Nat Sci,2012,38(6):617-622.
[8] 宁华.植物细胞核不同制备方法的比较研究[J].华中师范大学学报:自然科学版,2009,43(2):308-311.
NING Hua.Compared study on methods of plant nucleus preparation with different ways[J].J Huazhong Norm Univ Nat Sci,2009,43(2):308-311.
[9] 高猛,安玉麟,孙瑞芬.向日葵染色体Giemsa C-分带研究[J].植物遗传资源学报,2012,13(6):1078-1081.
GAO Meng,AN Yulin,SUN Ruifen.Giemsa C-banding patterns analysis of sunflower chromosomes[J].J Plant Genet Resour,2012,13(6):1078-1081.
[10] 李懋学,陈瑞阳.关于植物核型分析的标准化问题[J].武汉植物学研究,1985,3(4):297-302.
LI Maoxue,CHEN Ruiyang.A suggestion on the standardization of karyotype analysis in plants[J].J Wuhan Bot Res,1985,3(4):297-302.
[11] LEVAN A,FREDGA K,SANDBERG A A.Nomenclature for centromeric position on chromosomes[J].Hereditas, 1964,52(2):201-220.
[12] STEBBINS G L.Chromosome Evolution in Higher Plants[M].London:Edward Amold,1971.
[13] ARANO H.Cytological studies in subfamily Carduoidae(Comositae)of Japan(Ⅸ)the karyotype analysis and phylogenic consideration on Pertya and Ainsliaea[J].Bot Mag Tokyo,1963,76(895):32-39.
[14] KUO S R,WANG T T,HUANG T C.Karyotype analysis of some formosan gymnosperms[J].Taiwania,1972,17(1): 66-80.
[15] 卢玉飞,蒋建雄,易自力.利用Photoshop软件处理芒属植物染色体图像的方法探讨[J].草业科学,2013,30(6):922-926.
LU Yufei,JIANG Jianxiong,YI Zili.The protocol for processing of the chromosomal images of Misanthus for karyotype analysis using Adobe Photoshop software[J].Pratacultural Sci,2013,30(6):922-926.
[16] 刘泉,傅祖芸.一种植物染色体图像核型分析系统[J].计算机工程与应用,2000,36(3):71-74.
LIU Quan,FU Zuyun.A system of analyzing plant chromosome image’s karyotype[J].Comput Eng Appl,2000,36(3):71-74.
[17] 杨汉民.细胞生物学实验[M].北京:高等教育出版社,1997.
[18] 杨光穗,尹俊梅,冷青云,等.鹤蕉属植物8个品种间的核型比较分析[J].热带作物学报,2010,31(12):2118-2123.
YANG Guangsui,YIN Junmei,LENG Qingyun,et al.Karyotypes comparison of eight varieties of Heliconia spp.[J]. Chin J Trop Crops,2010,31(12):2118-2123.
Karyotypes in 13 Clematis cultivars
REN Jialun,JI Mengcheng,ZHAO Shuang,LIU Zhigao
(School of Landscape Architecture,Zhejiang A&F University,Lin’an 311300,Zhejiang,China)
Clematis has extremely high ornamental value,complicated genetic background and crossincompatibility phenomena.Studies on chromosome karyotype analysis of hybrid parent is very important. Chromosome types and karyotypes with natural growth of tender root tip materials in 13 Clematis cultivars were studied by the method of cell wall degradation and hypotonic treatment for chromosome preparations.Results indicated that all chromosomes for the 13 provenances were diploids and the chromosome number was 2n=2x =16.There were 2 types of chromosome karyotypes classified from the morphological observation:2n=2x= 16=10m+4st+2t and 2n=2x=16=10m+6st.Satellites were found in chromosomes of each cultivar for numbers 6,7,and 8.Both karyotypes were “2A”.Thus,both chromosome karyotypes had a great deal of similarity.So cross incompatibility phenomena may caused by other factors.[Ch,3 fig.3 tab.18 ref.]
plant cytogenetics;Clematis;chromosome;karyotype
S682.39;Q943
A
2095-0756(2016)06-1033-07
2015-12-22;
2016-02-12
浙江省科学技术重大资助项目(2012C12909-16)
任佳伦,从事野生园林植物资源分类与应用。E-mail:1416001262@qq.com。通信作者:刘志高,副教授,从事观赏植物栽培与育种技术研究。E-mail:1692362952@qq.com
10.11833/j.issn.2095-0756.2016.06.015
毛茛科Ranunculaceae铁线莲属Clematis为观赏价值极高的藤本植物,全世界大约有300种,中国约110种,分布于华中西南地区。铁线莲属植物花色美丽,花型特别,有 ‘攀援植物皇后’的美称,是非常优秀的垂直绿化材料,因具多种抗逆性,也是杂交育种的优良资源[1]。早在1885年,就有科学家GURIGNARD对铁线莲属植物进行了细胞学研究,证实C.recta的单倍染色体n=8,MEURMAN等在1939年研究发现了四倍体、六倍体与杂合染色体。据张镱锂等[2]的统计,该属约有106种与变种的染色体数目及部分种的核型有过记载,其他并未见报道。对铁线莲属的细胞学研究多集中在野生种上[2-6],而有关栽培品种的的研究则鲜有报道[7]。通过自然选择及人工杂交选育,铁线莲品种数量繁多,形态变异丰富,遗传背景也相对复杂,为铁线莲种质资源调查、品种分类与鉴定等工作带来了一定的难处。针对杂交育种工作中存在子一代生长周期较长,且本研究的13个品种间存在杂交不亲和性的障碍,因此,
对杂交亲本的染色体核型分析等细胞学特征的研究显得十分重要。本研究采用酶解法、去壁低渗火焰干燥法和Giemsa-C分带技术对13个铁线莲品种进行核型分析,研究核型差异是否是导致其杂交不亲和原因。
