近百年全球草地生态系统净初级生产力时空动态对气候变化的响应
2016-12-06刚成诚王钊齐杨悦陈奕兆张艳珍李建龙程积民
刚成诚,王钊齐,杨悦,陈奕兆,张艳珍,李建龙*,程积民
(1.西北农林科技大学水土保持研究所,陕西 杨凌 712100;2.中国科学院水利部水土保持研究所,陕西 杨凌 712100;3.南京大学生命科学学院,江苏 南京 210093)
近百年全球草地生态系统净初级生产力时空动态对气候变化的响应
刚成诚1,2,3*,王钊齐3,杨悦3,陈奕兆3,张艳珍3,李建龙3*,程积民1,2
(1.西北农林科技大学水土保持研究所,陕西 杨凌 712100;2.中国科学院水利部水土保持研究所,陕西 杨凌 712100;3.南京大学生命科学学院,江苏 南京 210093)
气候变化是影响生态系统空间地理分布、结构和功能的主要因素。为了从长时间序列大空间尺度上了解气候变化对草地生态系统的影响及其反馈机制,本研究利用综合顺序分类法及分段模型分别模拟了1911-2010年间全球草地生态系统及净初级生产力(NPP)的时空动态,并通过相关性分析揭示草地NPP对不同气候因子的响应。结果表明,在过去的百年间,全球草地面积从1920s的5175.73万km2下降到1990s的5102.16万km2,其中冻原与高山草地类组的面积下降最多,为192.35万 km2,荒漠草地类组、典型草地类组和温带湿润草地类组的面积分别下降了14.31、34.15和70.81万km2,而热带萨王纳类组的面积增加了238.06万km2。在气候变化的影响下,大多数草地类组的重心均向北方移动,在北半球尤为明显。全球草地NPP从1920s的25.93 Pg DW/年增加到1990s的26.67 Pg DW/年。就各草地类组而言,冻原与高山草地类组、荒漠草地类组、典型草地类组、温带湿润草地类组的NPP均呈现下降趋势,分别下降了709.57、24.98、115.74和291.56 Tg DW/年,而热带萨王纳类组的NPP则增加了1887.37 Tg DW/年。从全球尺度来看,降水是影响草地NPP的主要气候因子。总的来说,近百年气候变化对全球草地生态系统产生了深刻的影响,尽管草地NPP呈现增加的趋势,但暖湿化的气候变化对草地生态系统的结构和空间分布产生了不利的影响。
草地生态系统;综合顺序分类法(CSCS);净初级生产力(NPP);时空动态;分段模型
全球变化与陆地生态系统(global change-terrestrial ecosystems, GCTE)之间的相互作用是目前全球变化研究中的热点内容之一[1]。从宏观尺度来看,气候是决定植被类型和空间分布的最主要的非生物因素,持续升高的气温和变化的降水格局对陆地生态系统结构和功能产生了深刻的影响,而植被作为陆地生态系统的主体,对气候变化的响应最为直接[2-4]。草地是陆地生态系统的重要组成部分之一,约占陆地总面积的30%。草地生态系统不仅具有防风固沙、保持水土、涵养水源等生态功能,而且是食品安全的重要组成部分,它提供了人类食用的肉制品和奶制品,是畜牧业发展的重要经济资源。同时,由于其广大的分布面积,草地对平衡全球温室气体浓度,降低陆地温室效应具有重要意义[5-6]。草地主要分布在干旱半干旱地区,对全球气候变化极为敏感。目前,国内外关于气候变化对草地生态系统的影响进行了大量深入的研究[7-10]。在我国,气候变化已导致草地覆盖度总体呈现上升趋势,青藏高原地区草地植被生长季提前[11-12]。同时,气候变暖导致青藏高原地区各类冻土发生不同程度的退化[13]。
评估气候变化与草地生态系统之间相互作用的关键在于对草地生态系统的时空动态进行定量评估。气候条件以水热条件为核心,对草地的立地条件和动植物的分布与生长起着直接、广泛而持久的影响。地带性的生物气候条件决定了生物(牧草和家畜)的立地条件,因此影响着草地生态系统土-草-畜的各个组成部分的存在与发展,这种影响在相当长的时期内会保持稳定[14-15]。综合顺序分类法(Comprehensive Sequential Classification System, CSCS)就是以此为基础,根据农业生物气候特征,以水热为指标,在大空间尺度内划分草原的高级分类方法[14-16]。CSCS的提出为研究草地生态系统的发生发展及定量描述提供了重要手段,目前已得到广泛的应用[17-19]。马轩龙等[20]利用CSCS与地理信息系统相结合的方法对甘肃省的草地类型进行了探索,并制作了甘肃省草地类型分布图。吴静等[21]利用遥感数据的土壤水热指标代替传统站点大气水热指标,对CSCS进行了改进。赵明伟等[22]基于CSCS对中国北方草地地上生物量进行了高精度模拟,并分析每一种潜在植被类型的草地生物量的内在影响因素。
净初级生产力(net primary productivity, NPP)是描述植物群落在自然环境条件下生产能力的重要指标,它是指绿色植物通过光合作用固定有机物的量,为能量和营养在整个生态系统的食物链、食物网中循环提供了物质基础和动力来源[23]。NPP能够反映植物的生长状况,其变化反映了生态系统对环境因子变化的响应,因此可以作为衡量陆地生态系统响应气候变化研究的重要指标[24-25]。草地NPP是草地生态系统中土-草-畜系统与气候(光、水、热)等环境因子之间综合作用的结果,是草地生态系统结构和功能的综合指标,也是草地生态系统固碳能力的直接表征[26-27]。研究气候变化背景下草地NPP的变化对评价气候变化对草地生态系统结构和功能的影响及碳源/汇估算具有十分重要的意义。目前,草地NPP已在不同尺度开展了广泛的研究,在样地尺度主要采用地面观测生物量的方法,而区域或全球尺度的植被NPP估算主要以模型模拟为主[28-29]。气候生产力模型主要利用气候因子与植被干物质之间建立相关性,在不同区域不同植被类型上已得到验证,由于所用参数简单,易获取,在模拟植被NPP中具有独特的优势[28,30]。
草地植被动态与气候变化关系已开展了广泛的研究,但主要集中在特定时间和空间尺度,从长时间序列大空间尺度上分析气候变化对草地生态系统的影响的研究较少。鉴于此,本研究利用综合顺序分类法和基于湿润度指数的分段模型分别模拟近百年(1911-2010年)全球草地生态系统及NPP时空动态,并分析各草地类组在不同时间段的迁移距离和方向,最后通过相关性分析揭示影响草地NPP的主要气候因子。本研究不仅有助于了解全球气候变化对草地生态系统的影响及其反馈机制,且对陆地生态系统碳循环研究具有重要意义。
1 材料与方法
1.1 全球气候数据
CRU_TS 3.22数据库是由英国东安格利亚大学气候研究中心(Climatic Research Unit, CRU)提供[31]。该数据库通过整合已有的若干个数据库,重建了一套覆盖完整、分辨率高且无缺测的月平均地表气候要素数据集,时间范围为1901-2014年,覆盖全球0.5°经纬网格的所有陆地(除南极洲),该数据集已经被广泛应用于全球气候变化研究中[32]。本研究将月降水和月气温数据重新计算为年均温(mean annual temperature,MAT)和年降水量(mean annual precipitation,MAP)。
根据IPCC的研究报告,过去的30年很可能是过去800年中最热的30年,且依据大尺度气候变化研究中关于基准气候时间段的常用标准,为了更合理的描述全球草地生态系统的时空格局及动态特征,本研究将过去100年划分为4个时期,每30年为一个时期:即1920s (1911-1940年)、1950s (1941-1970年)、1970s (1961-1990年)和1990s (1981-2010年)。
1.2 综合顺序分类法(CSCS)及其改进
综合顺序分类法(CSCS)包括3个分类等级:类,亚类和型。类,是基本的分类单位,根据水分与热量分配特点来确定,具有地带性农业生物气候特征。在具体分类中,以生物气候指标——≥0 ℃年积温(∑θ)和湿润度指数K作为分类依据,以定量数据来说明不同草原类别的生物气候条件的核心——水热状况和它们的区别,其计算公式如下:
K=MAP/(0.1×∑θ)=MAP/(0.1×AAT)
(1)
式中:MAP是年降水量(mm),0.1为模型修正系数,AAT为≥0 ℃年积温。以湿润度等级为横坐标,大于0 ℃年积温为纵坐标,做出全球草地生态系统分类检索图。
CSCS理论是通过对具有类似性质的湿润度和温度的区域通过分组或集群进行植被类型的划分。不同等级的湿润度与年积温组成了不同的组合区,即为一个特定的景观类别。根据Olson等[33]对大陆极地冰雪的分类结果,本研究对原有CSCS分类系统进行了改进,增加了对极地/冰雪的识别。改进的CSCS模型确立了8个热量带和6个湿润度等级,共组合成48个类,并依据类之间的相关关系,结合生物气候指标和使用习惯,将48个类划分成10个类组(图1)。除去极地/冰雪、荒漠和森林景观,草地生态系统共有21个类,划分为5个类组,分别是:冻原与高山草地类组、荒漠草地类组、典型草地类组、温带湿润草地类组和热带萨王纳类组,每一类草地的命名均采用确定该区的热量级与湿润度相连。
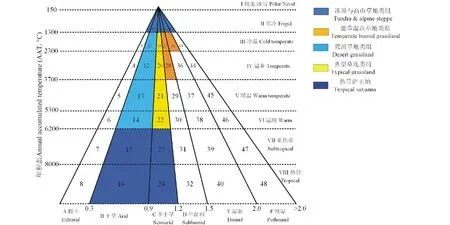
图1 改进的CSCS分类检索图Fig.1 The index chart of improved CSCS
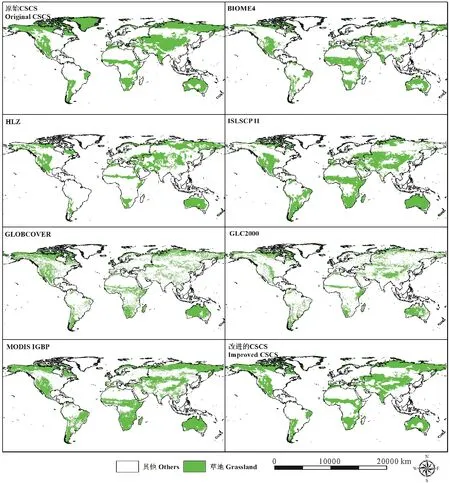
图2 改进的CSCS与其他植被覆盖数据中全球草地生态系统的分布Fig.2 The distribution of global grassland ecosystem simulated by improved CSCS and other land cover data
本研究对比了改进的CSCS与其他植被分类数据模拟的全球草地的空间分布(图2),其中CSCS、BIOME4、Holdridge Life Zone (HLZ)和The International Satellite Land-Surface Climatology Project (ISLSCP) II均为利用气温、降水和蒸散量等气候数据模拟得到的潜在草地分布,而GLOBCOVER、Global Land Cover 2000 (GLC2000)和MODIS IGBP的分类结果均是基于遥感技术得到的现实自然植被覆盖数据。根据计算结果原CSCS模拟的全球草地面积最大,为5955.55万km2,GLC2000的分类结果最小,仅为2844.91万km2,改进的CSCS模拟的全球草地分布面积为(5100.21±59.06)万km2,与GLOBCOVER的分类结果最为接近(表1)。
此外,利用验证BIOME4植被分类结果的全球3394个草地样点,本研究验证了改进的CSCS的模拟结果。该数据集依据区域尺度植被的生活型、叶型和物候等功能特征来定义群落。通过不同区域信息的融合和群落名称的标准化,共识别40个群落。为了便于对模型输出进行比较,基于结构和功能,将群落划分为 9个超级群落[45]。其中草地生态系统主要分为3个超级群落,即非木本草地&灌丛、萨王纳&木本植被、干/湿冻原,其样点数分别为1770、494和1130个(图3)。验证结果表明,共有2836个点与改进的CSCS模拟的草地植被相吻合。
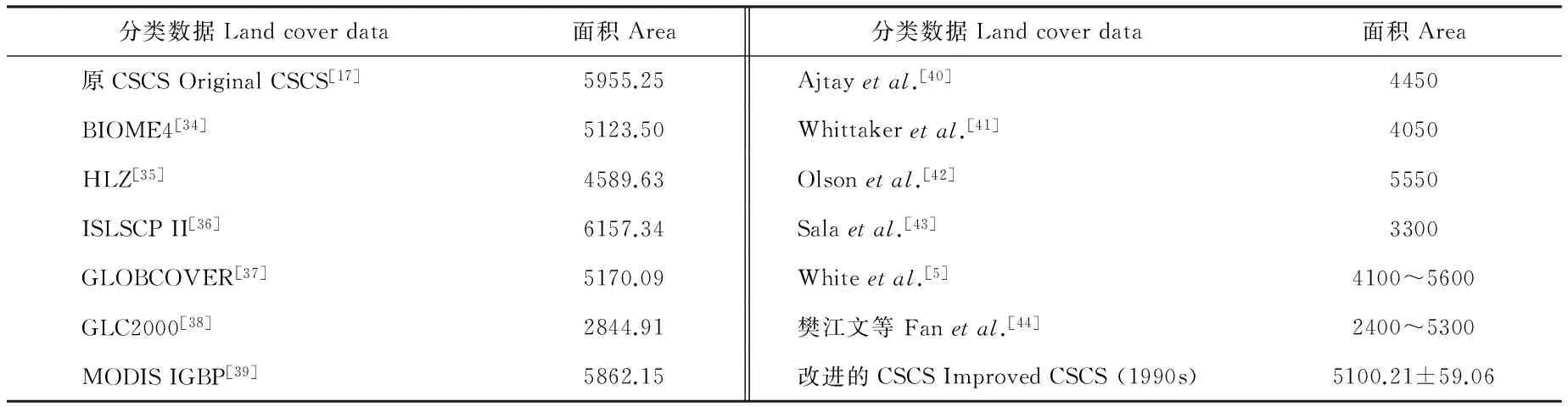
表1 改进的CSCS与其他研究中全球草地的分布面积
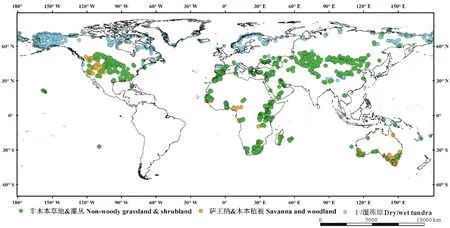
图3 全球不同草地类型样点分布图Fig.3 The spatial distribution of different grassland types across the globe
综上所述,经过对比验证,改进的CSCS不但继承了原分类系统的优势,而且可以更加准确合理的模拟全球草地植被分布。CSCS是基于土壤-植被-大气的分类方法,在模拟长时间序列草地植被动态的研究中具有重要的应用价值,尤其是对数据缺乏的时间和地区。然而,需要指出的是,CSCS并未考虑CO2施肥效应和氮沉降等气候变化对草地植被的影响,也未包含气候变化引起的火灾、虫灾和病原菌对草地分布的影响。该分类方法假设植物物种和种子能够在气候条件不利于生存时,无障碍的迁移或移动到其他地点来寻找新的生境。此外,该系统是一种自上而下的分类系统,未考虑到下垫面的因素,因此在模拟小区域尺度草地分布时,需进一步考虑地形和坡度、坡向等地理因素。尽管如此,CSCS为大尺度草地分类提供了一个新的方法,能够较为准确的反映长时间序列大空间尺度草地的地带性分布和对气候变化的响应。
1.3 重心模型
为了更好的描述气候变化对全球草地生态系统的影响,本研究利用不同草地类组重心的变化情况来反映其空间变化[17,46]。采用斑块中心函数来计算不同草地类组的重心:即通过计算同一草地类组不同斑块的中心点坐标,进而得到不同草地类组在各个时期的重心,其计算公式为:
(2)
式中:t是时间变量;Ij是指草地类组j的斑块数量;Sij(t)是指草地类组j的第i个斑块的面积;Sj(t)是指草地类组j的总面积;[Xij(t),Yij(t)]是指草地类组j的第i个斑块的面积的地理坐标;[xj(t),yj(t)]是指草地类组j的重心坐标。
根据其重心坐标,采用欧氏距离公式来计算不同草地类型分布重心在不同时段内的迁移距离,并确定其迁移方向,计算公式如下:
(3)
(4)
式中:Dj是草地类组j从t到t+1时刻的迁移距离;θj是草地类组j从t到t+1时刻的迁移方向。本研究定义0°表示正东方向,90°表示正北方向,180°和270°分别代表正西和正南方向。[xj(t),yj(t)]和[xj(t+1),yj(t+1)]分别代表草地类组j在t和t+1时刻的中心点坐标。如果0°<θj<90°,表示草地类组j从t到t+1时刻向东北方向移动,当90°<θj<180°,180°<θj<270°,270°<θj<360°时,则分别表示草地类组j向西北,西南和东南方向迁移。
1.4 NPP估算模型—分段模型
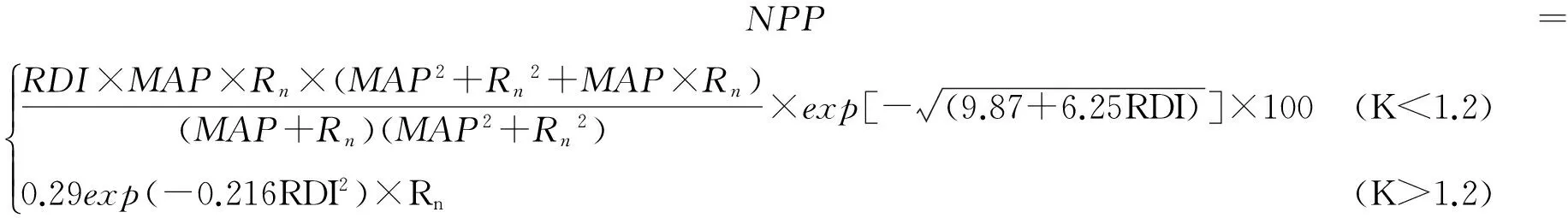
(5)
RDI=0.629+0.237PER-0.00313PER2
(6)
Rn=RDI×MAP×L×2.38×10-4
(7)
PER=PET/MAP=1.6145/K
(8)
式中:MAP是年降水量(mm),RDI是指辐射干燥度,由PER计算得到,PER是指潜在蒸散率,Rn是蒸散面的净辐射值[J/(cm2·a)],L是潜热通量,公式中采用2503 J/g,PET是潜在蒸散(mm),K是湿润度指数,由综合顺序分类法中公式求得,NPP是净初级生产力,单位是g DW/(m2·年)。
NPP实测数据来源于美国橡树岭国家实验室(Oak Ridge National Laboratories,ORNL)的净初级生产力数据库(Net Primary Production Database),该数据库覆盖了1931-1996年全球2523个样点和5164个0.5°网格数据,数据采集的样地包括了由生态区或气候区定义的多个植被类型,专门用来验证模型或者假说的结果[49-50]。模型模拟草地NPP与样地实测草地NPP对比结果表明,模型模拟结果与实测数据具有良好的一致性,分段模型用于模拟全球草地NPP具有较高的可信度(图4)。
1.5 草地NPP与气候因子的相关性分析
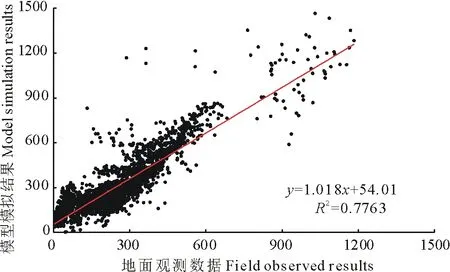
图4 分段模型模拟NPP结果与实测数据的比较Fig.4 The comparison of modelled NPP and field observation data
本研究采用基于像元的空间分析方法对草地NPP与MAT、MAP、湿润度指数K和年积温(AAT)进行Person相关性分析。如果相关系数通过显著性检验,则表示二者呈现显著(P<0.05)相关或极显著(P<0.01)相关。
2 结果与分析
2.1 近百年全球MAT与MAP时空动态
在1911-2010年间,全球MAT平均上升0.817 ℃,全球97.44%的地区的MAT均呈现上升趋势,其中亚欧大陆中高纬度的升温最为明显(图5)。1920s-1950s和1950s-1970s间气温分别上升0.099和0.035 ℃,而在1970s-1990s期间,MAT则上升了0.671 ℃。相比之下,MAP的变化呈现出较高的空间异质性。在过去的100年中,全球降水量平均增加了18.78 mm,全球65.02%的区域年降水量均增加。就全球草地分布区而言,近百年全球97.75%的草地分布区MAT呈现总体上升的趋势,平均上升0.889 ℃。同时,66.03%的草地分布区的MAP增加,平均增加了9.710 mm。总的来说,在1911-2010年间,全球64.57%的草地分布区的气候呈现暖湿化,33.18%的地区则呈现暖干化,同时冷干化和冷湿化的区域分别占0.79%和1.46%,其中暖干化的区域主要集中在非洲大陆撒哈拉地区和亚欧大陆的东部。
2.2 全球草地生态系统时空分布动态
不同时期全球草地生态系统的空间分布如图6所示。冻原与高山草地类组主要分布在北半球的高纬度和高海拔地区,如西伯利亚地区、格陵兰岛、北美洲北部及中国的青藏高原地区;温带湿润草地类组主要分布在加拿大北部以及欧亚大陆的中东部;典型草地类组是荒漠草地和温带湿润草地的过渡地带,主要分布在内蒙古、中亚部分地区以及北美洲的大平原地区;荒漠草地类组主要分布在北纬30°-60°之间的中亚地区、蒙古高原以及巴西高原的南部,并与典型草地镶嵌分布在北美大平原地区;热带萨王纳类组则主要分布在赤道热带雨林的两侧,包括印度半岛、非洲萨赫勒地区、非洲南部、北美大平原南部,巴西高原东北部,拉普拉塔平原部分地区,以及澳大利亚热带荒漠的周围。热带萨王纳类组的分布面积最大,其平均面积为(2010.05±108.32) 万km2,占草地总面积的39.14%;其次是冻原与高山草地,总面积为(1442.78±28.29) 万km2,占草地总面积的28.56%;荒漠草地和温带湿润草地的面积分别为(780.84±15.31)和(452.32±32.36) 万km2,分别占草地总面积的15.31%和8.87%;分布范围最小的是典型草地,其面积为(414.21±19.00) 万km2,占草地总面积的8.12%。
在过去的100年中,全球草地面积从5175.73万km2下降到5102.16万km2。其中,冻原与高山草地类组面积下降的最多,为192.35万 km2,荒漠草地类组、典型草地类组和温带湿润草地类组的面积分别下降了14.31、34.15和70.81万km2。具体来说,在过去的100年中,冻原与高山草地类组的分布面积逐渐下降,1970s-1990s间下降最多(153.68万km2,10.45%)。温带湿润草地的面积同样呈现持续下降的趋势,在1920s-1950s期间下降最多(55.75万km2)。荒漠草地类组和典型草地类组的变化趋势相同,即在1920s-1950s期间分布面积减少,之后逐渐增加。相比之下,气候变暖导致热带萨王纳类组的分布面积逐渐扩大,在过去的100年中共增加了238.06万km2,其中1970s-1990s期间增加了159.96万km2。
2.3 不同草地类组的迁移距离和方向
植被重心的变化进一步反映了气候变化对植被分布格局的影响。在北半球,热带萨王纳类组的重心分布在北纬20°附近,荒漠草地类组和典型草地类组的重心分布在北纬40°-45°之间,温带湿润草地类组的重心在北纬50°-52°,冻原与高山草地类组的重心则在北纬61°左右;在南半球,热带萨王纳类组的重心分布在南纬20°左右,冻原与高山草地类组、荒漠草地类组、典型草地类组和温带湿润草地类组的重心则集中在南纬30°-35°之间。

图5 近百年全球MAT和MAP的变化Fig.5 Changes of MAT and MAP in the recent 100 years
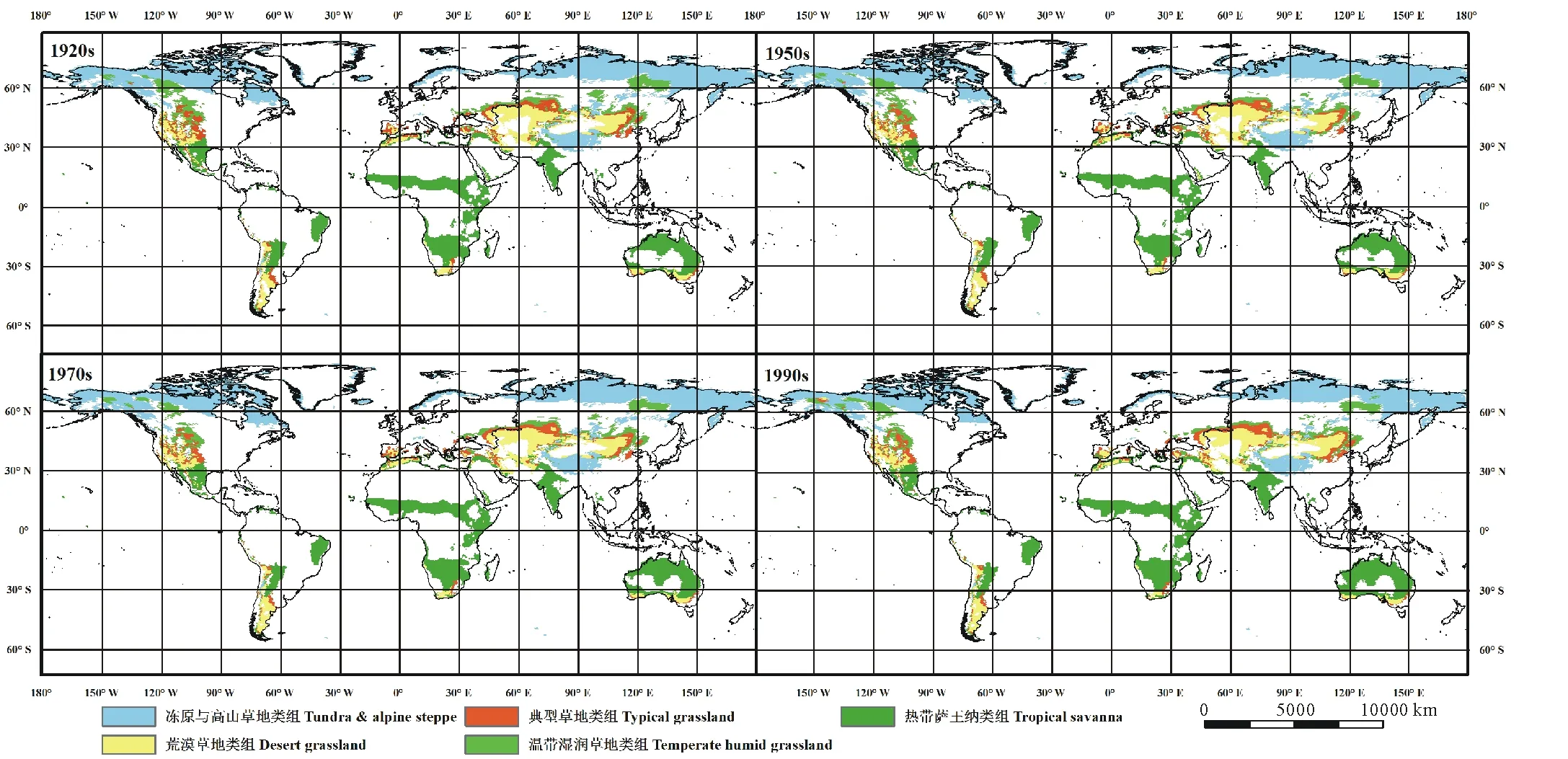
图6 近百年不同时期全球草地生态系统空间分布Fig.6 The spatial distribution of global grassland ecosystem in the recent 100 years
在1911-2010年间,气候变化导致北半球所有草地类组重心均向北移动,其中除温带湿润草地类组的重心向西北方向移动外,其余均向东北方向移动。典型草地类组重心移动距离最长,为633.11 km,冻原与高山草地类组、荒漠草地类组、温带湿润草地类组和热带萨王纳类组的重心分别移动了268.38、257.45、247.45和228.96 km。具体而言,冻原与高山草地类组与荒漠草地类组持续向东北方向移动,在1950s-1970s重心移动距离最长;典型草地类组和热带萨王纳类组的重心在1920s-1950s间向西移动,而之后向东移动,二者移动最长距离分别发生在1970s-1990s和1950s-1970s。温带湿润草地类组的重心在1920s-1950s,1950s-1970s和1970s-1990s分别向西南、东南和西北方向移动,3个时期移动距离较为接近(表2)。
在南半球,不同草地类型的迁移方向有很大差异,其中典型草地类组和温带湿润草地类组分别向西北方向迁移了102.48和233.99 km,冻原与高山草地类组则向东北方向迁移了55.38 km,荒漠草地类组的迁移距离最长,其向西南方向迁移了1289.75 km,而热带萨王纳类组则向东南方向迁移了645.10 km。典型草地类组和温带湿润草地类组重心持续向北方移动,而荒漠草地类组和热带萨王纳草地类组则持续向南移动,以上4类草地类组的重心最长移动距离均发生在1950s-1970s间。相比之下,冻原与高山草地类组重心在1920s-1950s,1950s-1970s和1970s-1990s分别向东北、西北和东南方向移动,最长移动距离发生在1970s-1990s。
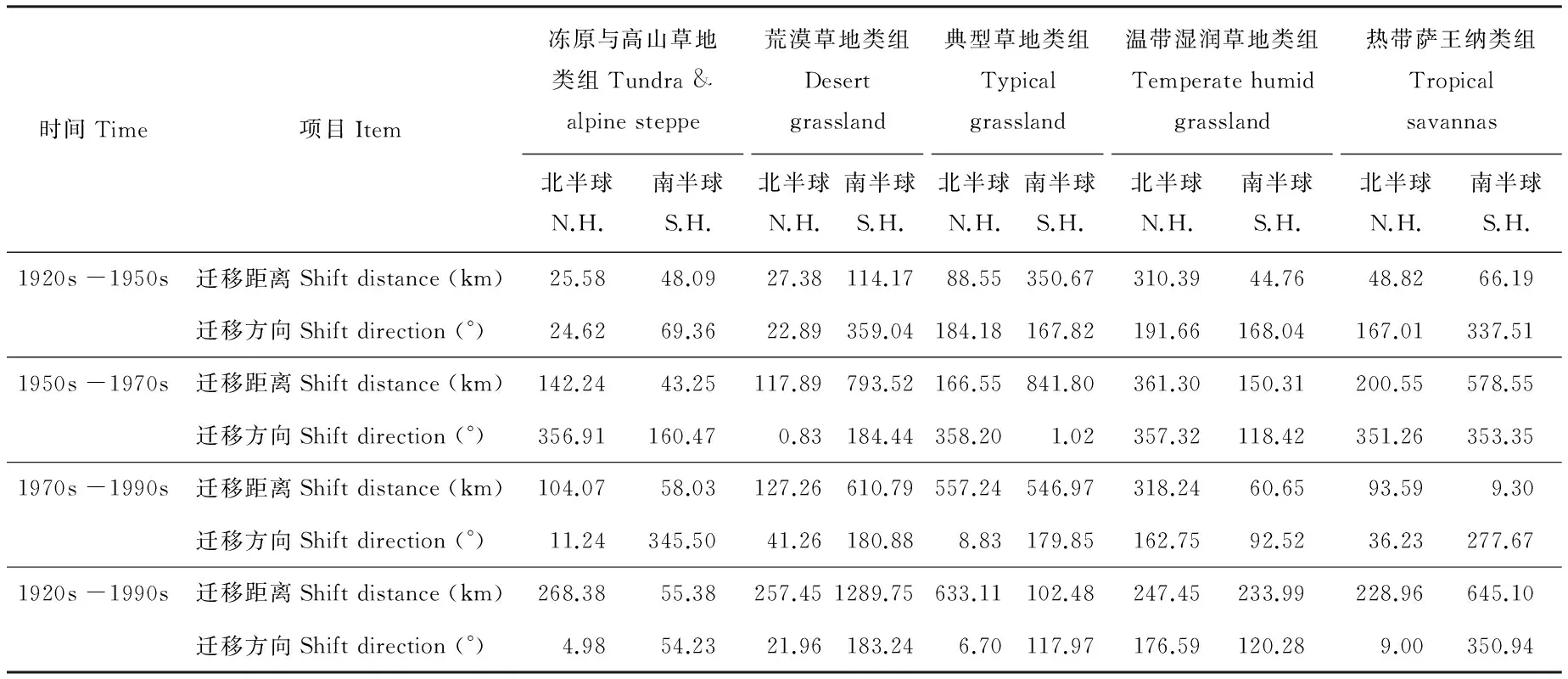
表2 近百年不同草地类组在南、北半球的迁移距离和迁移方向
2.4 近百年全球草地NPP的变化
在5个草地类组中,热带萨王纳类组的NPP最高,为(14.08±0.86) Pg DW/年,占全球草地NPP的53.97%;其次为冻原与高山草地类组,为(5.88±0.36) Pg DW/年,占全球草地NPP的22.54%;典型草地类组NPP最低,为(1.59±0.06) Pg DW/年,占全球草地NPP的6.09%;荒漠草地类组和温带湿润草地类组的NPP分别为(2.47±0.02)和(2.07±0.12) Pg DW/年,分别占全球草地NPP的9.47%和7.93%(图7)。
在过去的100年中,全球草地NPP由25.93 Pg DW/年增加到26.67 Pg DW/年,其中热带萨王纳类组的NPP呈现明显的上升趋势,而其他草地类组NPP呈现总体下降的趋势。在1911-2010年间,冻原与高山草地类组的NPP共下降了709.57 Tg DW/年,其中1970s-1990s期间下降最为明显。荒漠草地类组、典型草地类组和温带湿润草地类组的NPP分别下降了24.98、115.74和291.56 Tg DW/年。热带萨王纳类组的NPP逐渐上升,共增加了1887.37 Tg DW/年,其中1970s-1990s期间上升最多。
2.5 草地NPP与气候因子的相关性分析
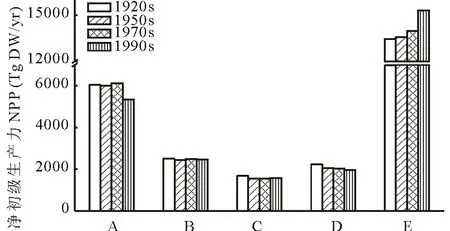
图7 近百年不同草地类组NPP变化Fig.7 The NPP change of each grassland type in the recent 100 yearsA:冻原与高山草地类组Tundra & alpine steppe;B:荒漠草地类组Desert grassland;C:典型草地类组Typical grassland;D:温带湿润草地类组Temperate humid grassland;E:热带萨王纳类组Tropical savannas.
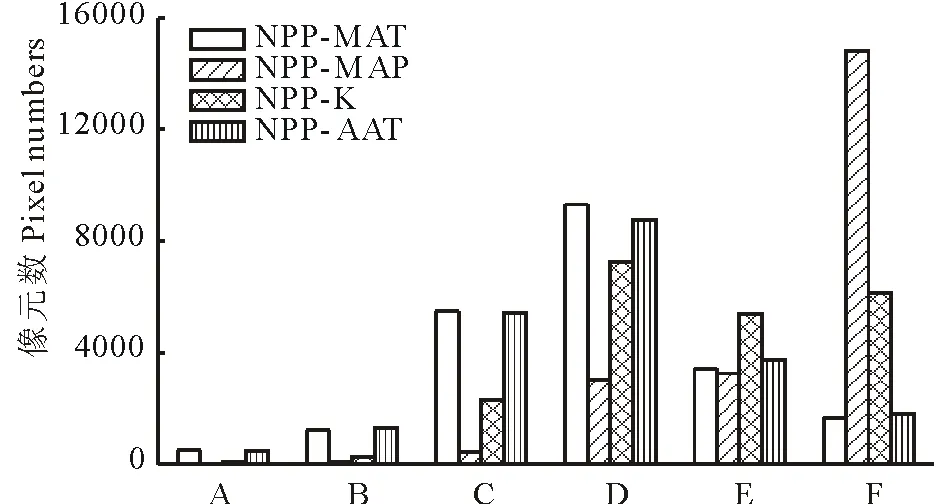
图8 全球草地NPP与MAT、MAP、K和AAT的相关性 Fig.8 The correlations between grassland NPP and MAT, MAP, K and AAT NPP: 净初级生产力Net primary productivity; MAT: 年均温 Mean annual temperature; MAP: 年降水量 Mean annual precipitation; K: 湿润度指数 Moisture index; AAT: ≥0 ℃年积温Annual accumulated temperature.A:极显著负相关Extremely significant negative correlation;B:显著负相关Significant negative correlation;C:不显著负相关Insignificant negative correlation;D:不显著正相关Insignificant positive correlation;E:显著正相关Significant positive correlation;F:极显著正相关Extremely significant positive correlation.
草地NPP与气候因子的相关性如图8所示。草地NPP与MAP的相关性最高,68.54%的草地NPP与MAP呈现极显著的正相关,置信区间在95%以上的地区达到83.95%,草地NPP与MAP呈现正相关关系的面积占97.60%;草地NPP与MAT的相关性较低,且呈现较高的空间异质性,尤其是在北半球。草地NPP与MAT相关性不显著的区域占草地总面积的68.50%。草地NPP与K的相关性高于与MAT的相关性,二者呈现正相关的区域占草地总面积的87.48%,其中达到显著正相关和极显著正相关的区域分别占25.07%和28.64%;草地NPP与AAT的相关性与MAT较为接近,二者无显著相关的面积占草地总面积的66.09%,而达到显著正相关和极显著正相关的区域分别占17.43%和8.34%。以上结果表明,从全球尺度来看,降水是影响草地NPP最主要的气候因子,NPP对降水的变化更加敏感,而在区域尺度,温度对草地NPP的影响则会更加突出。
3 讨论
近百年气候变化对全球草地生态系统产生了深刻影响,尤其是在北半球的中高纬度。在1911-2010年间,全球草地面积下降了73.58万km2,其中除热带萨王纳的分布面积增加外,其他草地类型的面积逐渐减少,其中冻原与高山草地的面积下降最多。持续的气温上升导致温带生态系统的面积不断扩大,气候变暖导致植被生长季延长,加之冬季温度上升促使温带森林向高纬度扩张,入侵冻原原始生境,冻原与高山植被的面积不断减少,这种现象在北美洲、亚洲和欧洲尤为明显[51-52]。此外,模型模拟结果显示过去百年中,冻原与高山草地类组的重心向东北方迁移了268.38 km,温带森林北缘的入侵导致冻原与高山草地的分布区整体向极地和高海拔地区迁移[53-54]。欧洲和美国西部的植被有向北方和高海拔迁移的现象,并且这种变化会持续到2100年[5,54]。热带萨王纳的扩张主要集中在非洲和大洋洲[55]。在南半球,南美洲荒漠草地面积增加,而在澳大利亚的面积减少,导致了荒漠草地的重心向西北方向移动。同时,澳大利亚热带萨王纳的分布面积下降导致其向南方移动,入侵荒漠草地的原生境。
气候变化导致温带植被向高海拔和高纬度移动,从而重新适应环境,且生产力下降。北半球中高纬度的生态系统对气候变化最为敏感,最易受影响,且适应气候变化的能力较差。温带森林入侵冻原与高山草地的南缘,致使其面积下降,单位面积草地生产力下降,导致总NPP降低。同样,典型草地分布区被荒漠草地入侵,NPP也总体下降。从不同草地类组来看,各个类组的面积变化趋势与NPP变化基本一致,冻原与高山草地类组、荒漠草地和温带湿润草地类组NPP整体呈现下降的趋势,而典型草地的分布面积较小,仅热带萨王纳的NPP逐渐增加。因此,热带萨王纳NPP主导着全球草地NPP的变化趋势。
相关性分析研究结果表明从全球尺度来看,草地NPP与MAP的相关性最高,说明降水是影响草地生产力最重要的气候因子,尤其在干旱半干旱地区,水分是草地NPP变化最主要的限制性因子[56]。但不同草地类组NPP对MAP和MAT的响应不同。冻原与高山草地类组主要分布在高纬度和高海拔地区,低温是植被生长最主要的限制因子。而对荒漠草地类组、典型草地类组和温带湿润草地类组而言,NPP与降水量的相关性较高,主要是因为水分是大陆性气候带的主导限制因子[57]。热带萨王纳类组分布区具有明显的干湿季,旱季干燥漫长,因此,降水是有机物质积累的主要限制因素。温度的变化对区域尺度草地生产力的影响更加明显,如Zhang等[30]的研究表明在过去的50年,内蒙古草地NPP的变化与温度的变化呈现明显的正相关。
4 结论
本文利用综合顺序分类法和分段模型模拟了1911-2010年间全球草地生态系统及NPP时空动态,并分析了草地NPP与不同气候因子的相关性,旨在从长时间序列大空间尺度揭示气候变化对草地生态系统的影响及其反馈机制。主要结论如下:
1) 在过去的100年中,全球草地面积由5175.73万km2下降到5102.16万km2,其中冻原与高山草地类组的面积下降最多,为192.35万 km2,荒漠草地类组、典型草地类组和温带湿润草地类组的面积分别下降了14.31、34.15和70.81万km2。热带萨王纳类组分布面积增加了238.06 万km2。
2) 在气候变化的影响下,大多数草地类组的重心均向北方移动。在南、北半球,重心迁移距离最长的草地类型分别为荒漠草地类组和典型草地类组,分别移动了1289.75和633.11 km。
3) 在1911-2010年间,全球草地NPP共上升了745.52 Tg DW/年。就各草地类组而言,冻原与高山草地类组、荒漠草地类组、典型草地类组、温带湿润草地类组的NPP均呈现下降趋势,分别下降了709.57、24.98、115.74和291.56 Tg DW/年,而热带萨王纳类组的NPP则增加了1887.37 Tg DW/年。
4) 草地NPP与气候因子的相关性研究结果表明,从全球尺度来看,降水是影响草地NPP的最主要气候因子,草地NPP对降水的变化最为敏感,而在部分地区或斑块尺度,温度的变化对草地NPP的影响更加明显。
[1] Walker B, Steffen W. IGBP Science No.1: A Synthesis of GCTE and Related Research[M]. Stockholm: IGBP, 1997: 1-24.
[2] Canadell J G, Pataki D E, Pitelka L F. Terrestrial Ecosystems in a Changing World[M]. German: Springer, 2006.
[3] Chinese Academy of Sciences. Department of Life Sciences of State Commission of Chinese National Natural Science Foundation, Global Changes and Ecological Systems[M]. Shanghai: Shanghai Science and Technology Press, 1994: 62-95.
[4] Zhang X S. A vegetation-climate classification system for global change studies in China. Quaternary Sciences, 1993, 2: 157-269.
[5] White R P, Murray S, Rohweder M,etal. Grassland Ecosystems[M]. Washington DC, USA: World Resources Institute 2000.
[6] Xie G D, Zhang Y L, Lu C X,etal. Study on valuation of rangeland ecosystem services of China. Journal of Natural Resources, 2001, 16(1): 47-53.
[7] Parton W J, Scurlock J, Ojima D S,etal. Impact of climate change on grassland production and soil carbon worldwide. Global Change Biology, 1995, 1(1): 13-22.
[8] Scurlock J, Hall D O. The global carbon sink: a grassland perspective. Global Change Biology, 1998, 4(2): 229-233.
[9] Fang J Y, Yang Y H, Ma W H,etal. Ecosystem carbon stocks and their changes in China’s grassland. Science of China: Life Science, 2010, 40(7): 566-576.
[10] Ren J Z, Liang T G, Lin H L,etal. Study on grassland’s responses to global climate change and its carbon sequestration potentials. Acta Prataculturae Sinica, 2011, 20(2): 1-22.
[11] Zhou W, Gang C C, Li J L,etal. Spatial-temporal dynamics of grassland coverage and its response to climate change in China during 1982-2010. Acta Geographica Sinica, 2014, 69(1): 15-30.
[12] Yang Y H, Piao S L. Variations in grassland vegetation cover in relation to climatic factors on the Tibetan Plateau. Journal of Plant Ecology, 2006, 30(1): 1-8.
[13] Wang G X, Hu H C, Wang Y B,etal. Response of alpine cold ecosystem biomass to climate changes in permafrost regions of the Tibetan Plateau. Journal of Glaciology and Geocryology, 2007, 29(5): 671-679.
[14] Ren J Z, Hu Z Z, Mu X D,etal. The comprehensive sequential classification system of grassland and its genesis significance. Grassland of China, 1980, 1(1): 6.
[15] Liang T G, Feng Q S, Huang X D,etal. Review in the study of comprehensive sequential classification system of grassland. Acta Prataculturae Sinica, 2011, 20(5): 252-258.
[16] Ren J Z. Classification and cluster applicable for grassland type. Acta Agrestia Sinica, 2008, 16(1): 4-10.
[17] Liang T G, Feng Q S, Cao J J,etal. Changes in global potential vegetation distributions from 1911 to 2000 as simulated by the Comprehensive Sequential Classification System approach. Chinese Science Bulletin, 2012, 57(11): 1298-1310.
[18] Ren J Z, Hu Z Z, Zhao J,etal. A grassland classification system and its application in China. Rangeland Journal, 2008, 30(2): 199-209.
[19] Lin H L, Wang X L, Zhang Y J,etal. Spatio-temporal dynamics on the distribution, extent, and net primary productivity of potential grassland in response to climate changes in China. Rangeland Journal, 2013, 35(4): 409-425.
[20] Ma X L, Li W J, Chen Q G. Preliminary exploration of native grassland classification of Gansu Province based on GIS and comprehensive and sequential grassland classification method. Pratacultural Science, 2009, 26(5): 7-13.
[21] Wu J, Li C B, Zhang D G,etal. Suggestions for improvement of moisture and temperature index in comprehensive and sequential classification system of grasslands suitable for 3S technology. Chinese Journal of Grassland, 2012, 34(4): 109-115.
[22] Zhao M W, Yue T X, Sun X F,etal. High accuracy simulation of aboveground biomass in Northern China based on IOCSG. Acta Ecologica Sinica, 2014, 34(17): 4891-4899.
[23] Odum E P, Barrett G W. Fundamentals of Ecology[M]. New York: Saunders College Publishing, 1971.
[24] Piao S L, Fang J Y. Terrestrial net primary production and its spatio-temporal patterns in Qinghai-Xizang Plateau, China during 1982-1999. Journal of Natural Resources, 2002, 17(3): 373-380.
[25] Gang C C, Zhou W, Li J L,etal. Assessing the spatiotemporal variation in distribution, extent and NPP of terrestrial ecosystems in response to climate change from 1911 to 2000. PloS One, 2013, 8(11): e80394.
[26] Lin H L, Chang S H, Li F. Research progress on grassland net primary productivity (NPP) model. Pratacultural Science, 2007, 24(12): 26-29.
[27] Scurlock J, Johnson K, Olson R J. Estimating net primary productivity from grassland biomass dynamics measurements. Global Change Biology, 2002, 8(8): 736-753.
[28] Gang C C, Zhou W, Wang Z Q,etal. Comparative assessment of grassland NPP dynamics in response to climate change in China, North America, Europe and Australia from 1981 to 2010. Journal of Agronomy and Crop Science, 2015, 201: 57-68.
[29] Ma W H, Yang Y H, He J S,etal. The temperate grassland biomass in Inner Mongolia and its relationship with environmental factors. Science of China: Life Science, 2008, 38(1): 84-92.
[30] Zhang G G, Kang Y M, Han G D,etal. Effect of climate change over the past half century on the distribution, extent and NPP of ecosystems of Inner Mongolia. Global Change Biology, 2011, 17(1): 377-389.
[31] University of East Anglia Climatic Research Unit (CRU), Jones P, Harris I. CRU Time Series (TS) High Resolution Gridded Datasets[DB/OL]. NCAS British Atmospheric Data Centre. (2015-12-22) [2016-03-01]. http://badc.nerc.ac.uk.
[32] Harris I, Jones P D, Osborn T J,etal. Updated high resolution grids of monthly climatic observations-the CRU TS3.10 Dataset. International Journal of Climatology, 2014, 34(3): 623-642.
[33] Olson D M, Dinerstein E, Wikramanayake E D,etal. Terrestrial ecoregions of the world: a new map of life on earth: a new global map of terrestrial ecoregions provides an innovative tool for conserving biodiversity. BioScience, 2001, 51(11): 933-938.
[34] Kaplan J O. Geophysical Applications of Vegetation Modeling[D]. Skane: Lund University, 2001.
[35] Holdridge L R. Determination of world plant formations from simple climate data. Science, 1947, 105: 367-368.
[36] Ramankutty N, Foley J A. ISLSCP II Potential Natural Vegetation Cover. ISLSCP Initiative II Collection, Data set[DB/OL]. Oak Ridge National Laboratory Distributed Active Archive Center. (2010-01-25) [2016-03-10]. http://daac. ornl. gov.
[37] Bontemps S, Defourny P, Bogaert E V,etal. GLOBCOVER 2009-Products Description and Validation Report[R]. Belgiqu: Université Catholique de Louvain, 2011.
[38] Bartholomé E, Belward A S. GLC2000: a new approach to global land cover mapping from Earth observation data. International Journal of Remote Sensing, 2005, 26(9): 1959-1977.
[39] Justice C O, Townshend J, Vermote E F,etal. An overview of MODIS Land data processing and product status. Remote Sensing of Environment, 2002, 83(1): 3-15.
[40] Ajtay G L, Ketner P, Duvigneaud P. Terrestrial primary production and phytomass. The Global Carbon Cycle, 1979, 13: 129-182.
[41] Whittaker R H, Likens G E. Primary production: the biosphere and man. Human Ecology, 1973, 1(4): 357-369.
[42] Olson. Major World Ecosystem Complexes Ranked by Carbon in Vegetation[DB/OL]. Oak Ridge National Laboratory Distributed Active Archive Center. (1985-09-10) [2016-03-10]. http://daac. ornl. gov.
[43] Sala O E, Parton W J, Joyce L A,etal. Primary production of the central grassland region of the United States. Ecology, 1988, 69(1): 40-45.
[44] Fan J W, Zhong H P, Liang B,etal. Carbon stock in grassland ecosystem and its affecting factors. Grassland of China, 2003, (6): 52-59.
[45] Harrison S P, Prentice I C. Climate and CO2controls on global vegetation distribution at the last glacial maximum: analysis based on palaeovegetation data, biome modelling and palaeoclimate simulations. Global Change Biology, 2003, (9): 983-1004.
[46] Yue T X, Fan Z M, Chen C F,etal. Surface modelling of global terrestrial ecosystems under three climate change scenarios. Ecological Modelling, 2011, 222(14): 2342-2361.
[47] Zhou G S, Zhang X S. A natural vegetation NPP model. Journal of Plant Ecology, 1995, 19(3): 193-200.
[48] Uchijima Z, Seino H. Agroclimatic evaluation of net primary productivity of natural vegetation. 1. Chikugo model for evaluating net primary productivity. Journal of the Meteorological Society of Japan, 1985, 40: 343-352.
[49] Olson R J, Scurlock J, Prince S D,etal. NPP Multi-Biome: Global Primary Production Data Initiative Products, R2. Data set[DB/OL]. Oak Ridge National Laboratory Distributed Active Archive Center. (2013-08-29) [2016-03-10]. http://daac. ornl. gov.
[50] Jager H I, Hargrove W W, Brandt C C,etal. Constructive contrasts between modeled and measured climate responses over a regional scale. Ecosystems, 2000, 3(4): 396-411.
[51] Hickler T, Vohland K, Feehan J,etal. Projecting the future distribution of European potential natural vegetation zones with a generalized, tree species-based dynamic vegetation model. Global Ecology and Biogeography, 2012, 21(1): 50-63.
[52] Cruz R V, Harasawa H, Lal M,etal. Climate Change 2007: Impacts, Adaptation and Vulnerability. Contribution of Working Group II to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change[M]. Cambridge: Cambridge University Press, 2007: 469-506.
[53] Berthelot M, Friedlingstein P, Ciais P,etal. Global response of the terrestrial biosphere to CO2and climate change using a coupled climate-carbon cycle model. Global Biogeochemical Cycles, 2002, 16(4): 1084.
[54] Callaghan T V, Björn L O, Chapin Iii F S,etal. Arctic Tundra and Polar Ecosystems[M]//Arctic Climate Impact Assessment. Cambridge: Cambridge University Press, 2005: 243-252.
[55] Wigley B J, Bond W J, Hoffman M. Thicket expansion in a South African savanna under divergent land use: local vs. global drivers. Global Change Biology, 2010, 16(3): 964-976.
[56] Ran G, Wang X, Ouyang Z,etal. Spatial and temporal relationships between precipitation and ANPP of four types of grasslands in northern China. Journal of Environmental Sciences, 2006, 18(5): 1024-1030.
[57] Liu X N, Ren Z C, Li C B,etal. Net primary productivity of grasslands in China under climate change. Grassland and Turf, 2010, 30(3): 7-14.
[6] 谢高地, 张钇锂, 鲁春霞, 等. 中国自然草地生态系统服务价值. 自然资源学报, 2001, 16(1): 47-53.
[9] 方精云, 杨元合, 马文红, 等. 中国草地生态系统碳库及其变化. 中国科学: 生命科学, 2010, 40(7): 566-576.
[10] 任继周, 梁天刚, 林慧龙, 等. 草地对全球气候变化的响应及其碳汇潜势研究. 草业学报, 2011, 20(2): 1-22.
[11] 周伟, 刚成诚, 李建龙, 等. 1982-2010年中国草地覆盖度的时空动态及其对气候变化的响应. 地理学报, 2014, 69(1): 15-30.
[12] 杨元合, 朴世龙. 青藏高原草地植被覆盖变化及其与气候因子的关系. 植物生态学报, 2006, 30(1): 1-8.
[13] 王根绪, 胡宏昌, 王一博, 等. 青藏高原多年冻土区典型高寒草地生物量对气候变化的响应. 冰川冻土, 2007, 29(5): 671-679.
[14] 任继周, 胡自治, 牟新待, 等. 草原的综合顺序分类法及其草原发生学意义. 中国草原, 1980, 1(1): 6.
[15] 梁天刚, 冯琦胜, 黄晓东, 等. 草原综合顺序分类系统研究进展. 草业学报, 2011, 20(5): 252-258.
[16] 任继周. 分类, 聚类与草原类型. 草地学报, 2008, 16(1): 4-10.
[20] 马轩龙, 李文娟, 陈全功. 基于GIS与草原综合顺序分类法对甘肃省草地类型的划分初探. 草业科学, 2009, 26(5): 7-13.
[21] 吴静, 李纯斌, 张德罡, 等. 适用于3S技术的草地综合顺序分类法水热指标改进. 中国草地学报, 2012, 34(4): 109-115.
[22] 赵明伟, 岳天祥, 孙晓芳, 等. 基于草地综合顺序分类系统(IOCSG)的中国北方草地地上生物量高精度模拟. 生态学报, 2014, 34(17): 4891-4899.
[24] 朴世龙, 方精云. 1982-1999年青藏高原植被净第一性生产力及其时空变化. 自然资源学报, 2002, 17(3): 373-380.
[26] 林慧龙, 常生华, 李飞. 草地净初级生产力模型研究进展. 草业科学, 2007, 24(12): 26-29.
[29] 马文红, 杨元合, 贺金生, 等. 内蒙古温带草地生物量及其与环境因子的关系. 中国科学: 生命科学, 2008, 38(1): 84-92.
[44] 樊江文, 钟华平, 梁飚, 等. 草地生态系统碳储量及其影响因素. 中国草地, 2003, (6): 52-59.
[47] 周广胜, 张新时. 自然植被净第一性生产力模型初探. 植物生态学报, 1995, 19(3): 193-200.
[57] 柳小妮, 任正超, 李纯斌, 等. 气候变化下中国草地 NPP 的研究. 草原与草坪, 2010, 30(3): 7-14.
The NPP spatiotemporal variation of global grassland ecosystems in response to climate change over the past 100 years
GANG Cheng-Cheng1,2,3*, WANG Zhao-Qi3, YANG Yue3, CHEN Yi-Zhao3, ZHANG Yan-Zhen3, LI Jian-Long3*, CHENG Ji-Min1,2
1.InstituteofSoilandWaterConservation,NorthwestA&FUniversity,Yangling712100,China; 2.InstituteofSoilandWaterConservation,ChineseAcademyofSciencesandMinistryofWaterResources,Yangling712100,China; 3.SchoolofLifeSciences,NanjingUniversity,Nanjing210093,China
Climate change has a significant effect on the spatial distribution, structure and function of ecosystems. To understand the effects and feedback mechanisms of climate change on a large spatial and temporal scale, the distribution, shift ranges and net primary production (NPP) of grassland ecosystems across the globe from 1911 to 2010 have been simulated using the Comprehensive Sequential Classification System (CSCS) and a segmentation model. Correlation analysis was also conducted to reveal the responses of grassland types to different climate variables. The results showed that the total global area of grassland ecosystems declined from 5175.73×104km2in the 1920s to 5102.16×104km2in the 1990s. The largest decrease, 192.35×104km2, occurred in tundra & alpine steppe ecosystems. The areas of desert grassland, typical grassland and temperate humid grassland decreased by 14.31, 34.15 and 70.81×104km2respectively, while tropical savanna expanded by 238.06×104km2. Climate warming forced most grasslands to shift northwards, particularly in the northern hemisphere. Global grassland NPP increased from 25.93 Pg DW/yr in the 1920s to 26.67 Pg DW/yr in the 1990s. In terms of each grassland type, the NPP of the tundra and alpine steppe, desert grassland, typical grassland and temperate humid grassland decreased by 709.57, 24.98, 115.74 and 291.56 Tg DW/yr respectively. The NPP of tropical savanna increased by 1887.37 Tg DW/yr. At the global scale, precipitation was the dominant factor affecting grassland NPP. In general, grassland ecosystems have been substantially affected by climate change over the past 100 years. Although the global grassland NPP showed an overall increasing trend, the structure and distribution of particular grassland ecosystems had been adversely affected by the warmer and wetter climate.
grassland ecosystem; comprehensive sequential classification system (CSCS); net primary productivity (NPP); spatiotemporal dynamics; the segmentation model
10.11686/cyxb2016148
http://cyxb.lzu.edu.cn
2016-04-05;改回日期:2016-05-10
国家自然科学基金(31602004),西北农林科技大学博士科研启动基金项目(2452015339)和国家重点基础研究发展计划项目(2010CB950702)资助。
刚成诚(1986-),男,满族,辽宁灯塔人,助理研究员,博士。E-mail: gangcheng024@gmail.com*通信作者Corresponding author. E-mail: gangcheng024@gmail.com, jianlongli@gmail.com
刚成诚, 王钊齐, 杨悦, 陈奕兆, 张艳珍, 李建龙, 程积民. 近百年全球草地生态系统净初级生产力时空动态对气候变化的响应. 草业学报, 2016, 25(11): 1-14.
GANG Cheng-Cheng, WANG Zhao-Qi, YANG Yue, CHEN Yi-Zhao, ZHANG Yan-Zhen, LI Jian-Long, CHENG Ji-Min. The NPP spatiotemporal variation of global grassland ecosystems in response to climate change over the past 100 years. Acta Prataculturae Sinica, 2016, 25(11): 1-14.
