巴山榧树种子特性与幼苗生长规律
2016-12-04长师范学院生命科学与技术学院重庆涪陵40800武陵山区特色资源开发与利用研究中心重庆涪陵40800
, , , , , , (.长师范学院生命科学与技术学院, 重庆 涪陵 40800;2.武陵山区特色资源开发与利用研究中心, 重庆 涪陵 40800)
巴山榧树种子特性与幼苗生长规律
江波1,2,周先容1,2,龚练1,秦明一1,2,盛亚莉1,郑永红1,李枝1
(1.长江师范学院生命科学与技术学院, 重庆 涪陵 408100;2.武陵山区特色资源开发与利用研究中心, 重庆 涪陵 408100)
采用室内和田间试验,研究了巴山榧树3个地理种群种子形态特征、种子萌发特性及幼苗生长规律。结果表明,巴山榧树种子形态特征在种群内和种群间均存在一定程度的变异,重庆南川种群种子变异较大,甘肃武都种群相对较小;3个种群种子大小顺序表现为,甘肃武都gt;四川万源gt;重庆南川,长度、宽度和重量在种群间差异显著或极显著,长宽比在重庆南川种群和甘肃武都种群间差异显著,重庆南川种群种子在形状上趋向椭圆形,甘肃武都种群种子则更趋圆球形。巴山榧树种子具有休眠现象,萌发率较低;1年生幼苗苗高呈“S”型曲线增长;3个种群种子萌发率、幼苗苗高与种子大小顺序表现一致,种子大小对种子萌发和幼苗生长有明显影响。应加强巴山榧树的保护和繁育。
巴山榧树; 种子特性; 育苗试验; 种子萌发; 幼苗生长
表1 巴山榧树种子的采集信息

采集地点经纬度海拔(m)坡向坡度群落类型样本数量(株)采种数量(粒)重庆南川金佛山107°08′E,28°56′N1340~1650SE5°35°灌丛15336四川万源花萼山108°08′E,32°07′N1200~1650NW75°30°针阔叶混交林12204甘肃武都洛塘镇105°17′E,33°01′N1580~1720NE25°45°针阔叶混交林9178
巴山榧树(TorreyafargesiiFranch.)又名篦子杉、紫柏、托杉、铁头枞和球果榧等,隶属于红豆杉科(Taxaceae)榧树属(Torreya),是我国特有的裸子植物,1999年被列为Ⅱ级重点保护野生植物[1]。目前,该物种零星分布于四川、贵州、陕西、甘肃、湖北、湖南、河南、安徽、重庆等省市海拔800~2 700 m的亚热带山区[2-3]。巴山榧树木材坚硬,结构细致,可做家具、农具等,种子可榨油,同时也是我国亚热带山区森林更新和荒山造林优良树种,具有较高的经济价值和生态价值[4]。由于巴山榧树的生境破碎化严重,种群自然更新能力差,其分布范围不断缩小,天然种群及规模急剧下降,物种处于易危状态[2-3]。
到目前为止,对巴山榧树的研究主要集中在地理分布[2]、资源[3]、遗传多样性[5-7]和群落生态[8-10]等方面,但关于巴山榧树种子特性与育苗技术的研究鲜有报道。种子和幼苗是植物整个生活史中十分关键的阶段,探讨濒危植物的种子特性及幼苗生长规律,对于濒危植物保护、苗木培育及森林营建具有重要的意义[11-14]。本研究以巴山榧树3个不同地理种群的种子为种源,探讨其种子特性、育苗技术和幼苗生长规律,为深入研究该物种的遗传多样性和种苗培育奠定基础,对于巴山榧树的保护和种群恢复具有十分重要的理论和实践意义。
1 材料与方法
1.1 材 料
供试的巴山榧树种子来源见表1。2013年9月上旬种子成熟时采集种子,由于巴山榧树结实植株和成熟种子数量稀少,因此尽可能将每个种源地样本植株的所有成熟种子全部采集,并按照种源地和样本植株分装编号,带回实验室备用。每个种源地的样本植株数量和采种数量见表1。
1.2 方 法
1.2.1 种子形态特征的测定
将采回的巴山榧树种子置于阴凉通风处,及时去除假种皮并漂洗干净,自然风干后测定种子的形态指标。随机选取饱满的种子,每个种源地设3个重复,每个重复50粒种子,用精度为0.01 cm的数显游标卡尺测量种子最长和最宽处作为种子的长度和宽度,计算长宽比及各指标的平均值。同时用万分之一电子天平测定种子重量,计算平均值。
1.2.2 种子萌发试验
于2013年10月12日开始种子萌发试验。用1 g/L高锰酸钾溶液分别对不同种源地的种子消毒2 h,经清水冲洗干净后,采用湿沙层积法处理种子,湿沙与种子按体积比5∶1混合拌匀,堆积在通风的室温环境中进行种子萌发试验。试验期间,适时适量喷水保持湿沙的含水量在70%左右,并翻动通气。种子发芽以胚根突破种皮为标志,每周定时观察记录发芽种子数,并记录种子萌发开始和结束时间,统计种子萌发率。种子萌发率(%)=最终萌发种子数/供试种子总数×100%。种子萌发试验于2015年3月10日结束,共计76周。
1.2.3 播种育苗试验
1) 苗圃地选择:试验地设在长江师范学院校园,位于重庆市涪陵城区,属亚热带东部湿润季风气候,年均气温18 ℃,年均有效积温5 100 ℃,年均降水量1 200 mm,年均日照时数1 248 h,年无霜期310~320 d[15]。选择土层深厚、疏松、肥沃的沙壤土苗圃地。
2) 播种育苗:清除杂草和石砾,深挖翻土20 cm以上,整地做成苗床,苗床高20 cm,宽1.2 m,周围挖40 cm宽的排水沟。播种前适量施农家肥,并进行土壤消毒,将1 g/L高锰酸钾溶液均匀喷洒于床面,随后用塑料薄膜覆盖苗床,2 d后将湿沙层积发芽种子按不同种源地分区播于苗床上。于3月初播种,采用条播,种子间距10 cm×10 cm,种子横放,胚根向下,上面覆盖2 cm左右的细沙,浇透水。搭建简易塑料棚,以提高土壤温度,促进根系生长和幼芽发育。幼苗出苗前,每隔4 d左右浇透水1次。幼苗出土后,随着气温升高、光照增强,拆除塑料薄膜,及时换上遮阳网,避免强光和高温对幼苗的危害;勤浇水,保持土壤湿润;及时除草、疏松土壤;每7 d喷洒1 g/L多菌灵可湿性粉剂,预防立枯病;用呋喃丹拌土施入苗床,诱杀防治金龟子、地老虎等有害生物。播种后,每天观察出苗及幼苗生长情况,记录出苗开始和结束时间,统计出苗率和幼苗成活率。
表2 巴山榧树不同地理种群种子形态特征

种群 长 度 宽 度 长宽比 重 量 平均值(cm)变异系数(%)平均值(cm)变异系数(%)平均值变异系数(%)平均值(g)变异系数(%)重庆南川2.1256±0.0284cC12.331.7644±0.0078cC11.881.2092±0.0188aA8.981.6697±0.0085cB24.34四川万源2.5169±0.0536bB10.892.1362±0.0381bB10.311.1811±0.0248abA7.472.0992±0.0193bB18.11甘肃武都2.9345±0.0582aA10.092.5233±0.0411aA9.431.1653±0.0226bA6.903.0374±0.0304aA19.67
注:表中同列数值后不同大、小写字母分别表示在0.01和0.05水平上差异显著。下同。
3) 幼苗年生长规律观测:选取不同种源地出苗基本一致的幼苗各20株,每月定期测定每株幼苗的苗高,计算平均值。苗高用直尺测量,精确至0.1 cm。
1.2.4 数据统计与分析
采用Excel软件进行数据处理与作图,DPS软件进行数据统计分析,LSD多重比较方法进行差异显著性检验。
2 结果与分析
2.1 巴山榧树种子形态特征
巴山榧树不同地理种群种子的形态特征见表2。在种群内部,以位于南方的重庆南川种群种子的各项形态指标变异较大,位于北方的甘肃武都种群相对较小。3个种群间以甘肃武都种群种子最大最重,平均长约2.93 cm,宽约2.52 cm,重量约3.04 g;重庆南川种群种子最小最轻,平均长约2.13 cm,宽约1.76 cm,重量约1.21 g。3个种群间种子长度和宽度的差异达到极显著水平(plt;0.01),重量的差异则达到显著水平(plt;0.05),仅甘肃武都种群与其他2个种群间种子的重量差异极显著。巴山榧树种子的长宽比以重庆南川种群最高,甘肃武都种群最低,两者间差异显著,重庆南川种群种子在形状上趋向椭圆形,甘肃武都种群种子则更趋圆球形。因此,巴山榧树种子形态存在明显的地理变异,种子大小顺序表现为,甘肃武都gt;四川万源gt;重庆南川;种子的长宽比则与之相反,其顺序为,重庆南川gt;四川万源gt;甘肃武都。
2.2 巴山榧树种子萌发规律
巴山榧树不同地理种群种子的萌发进程及萌发率见图1。3个种群的种子经过湿沙层积10~11周,即在当年12月中下旬开始萌发,重庆南川种群萌发较早,四川万源种群和甘肃武都种群的种子萌发较重庆南川种群滞后1周;13~18周,即第2年1月上旬至2月中旬为种子萌发高峰期;26~57周,即4月中旬至11月中旬,为种子萌发停滞期;58~67周,每个种群均有少数种子萌发,第3年1月下旬后种子不再萌发。巴山榧树种子萌发率较低,3个种群的平均萌发率为31.53%,且不同地理种群间存在明显差异,其中甘肃武都种群的萌发率最高,为45.51%;重庆南川种群的萌发率最低,为22.62%,几乎仅为甘肃武都种群的1/2。因此,巴山榧树种子萌发特性的地理差异表现较为明显,萌发率由高到低的顺序为,甘肃武都gt;四川万源gt;重庆南川,萌发率顺序表现出与种子大小一致的特点。

图1 巴山榧树不同地理种群种子萌发进程及萌发率
2.3 巴山榧树幼苗生长规律
播种25 d后,幼苗陆续出土。3个种群的平均出苗率为83.46%,甘肃武都种群的出苗率最高,达92.59%,重庆南川种群的出苗率最低,为76.32%。3个种群1年生幼苗的成活率差异不大,介于90.67%~91.38%之间。巴山榧树1年生幼苗苗高呈“S”型曲线增长(图2),其生长经历了一个“慢—快—慢”的过程,生长初期增长较缓慢,6月后进入高生长期,9月后增长变缓,直至11月上旬幼苗停止生长,生长期近8个月,属于全期生长类型。3个种群1年生幼苗苗高平均为18.85 cm,其中甘肃武都种群最高,达21.03 cm;重庆南川种群最低,为17.28 cm。因此,巴山榧树3个不同地理种群间种子出苗率和幼苗苗高的大小顺序与种子大小表现一致。
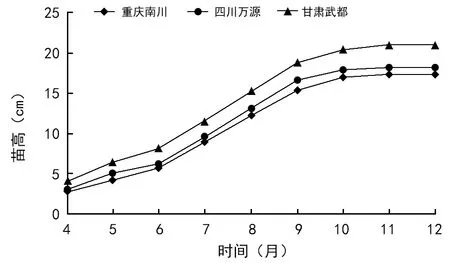
图2 巴山榧树不同地理种群1年生幼苗苗高
3 结论与讨论
3.1 巴山榧树种子形态变异
来自重庆南川、四川万源和甘肃武都的3个巴山榧树地理种群,其种子形态存在明显的地理变异,3个地理种群间种子平均长度、宽度和重量差异均达到显著或极显著水平,种子大小顺序为,甘肃武都gt;四川万源gt;重庆南川;种子长宽比在重庆南川和甘肃武都种群间差异显著,重庆南川种群种子在形状上趋向椭圆形,甘肃武都种群种子更趋圆球形。已有研究表明,种子形态变异既受遗传的控制,又受环境中生物因素(如捕食、寄生、竞争等)和非生物因素(光、温、水、海拔、纬度等)的影响[16-17]。通常认为,随着纬度的北移和环境干燥度的提高,种子趋大变重[18]。国内研究者已在白皮松(Pinusbungeana)[18]、乌桕(Sapiumsebiferum)[19]等植物种子形态变异的研究中得到印证。在本研究中,巴山榧树也表现出相同的趋势,来自北方(甘肃武都)种群的种子最大最重,而南方(重庆南川)种群的种子最小最轻。然而,长叶榧(Torreyajackii)[12]、青冈(Cyclobalanopsisglauca)[20]等物种却呈现出相反的规律,即种子随着纬度的南移而变大。这表明影响种子大小变异的因素十分复杂,不能简单地归于纬度这一个因素。目前,种子变异的机制尚不明晰,有待进一步研究。
3.2 巴山榧树种子休眠特性
种子休眠是具有正常活力的种子在适宜的环境条件下仍不能萌发的现象,其原因大致可归为由内源因素引起的胚休眠和由外源因素引起的种壳休眠[21]。本研究采用广泛使用的湿沙层积法[22-24]贮藏巴山榧树种子,试验结果表明,种子萌发高峰期在第2年1月上旬至2月中旬,在经历长达7个月的萌发停滞期后,于第2年11—12月和第3年1月仍有少数种子萌发。因此,巴山榧树种子具有休眠现象。巴山榧树种子虽有坚韧的种壳,但经过湿沙层积10周后,种子即开始萌发,这说明种壳的限制不是巴山榧树种子休眠的主要原因。史忠礼等研究发现,香榧(Torreyagrandis‘Merrillii’)采收时其外部形态虽已表现成熟,但胚的分化不完善,尚处于原胚时期,经10~20 ℃湿沙层积2个月后,胚器官分化完成[24]。因此,巴山榧树种子休眠的主要原因可能与同属物种香榧一样,是由于胚休眠引起的。胚休眠包括胚形态发育不全、生理后熟或存在抑制物质等,可以用低温层积、变温处理和激素处理等方法解除[21]。本试验采用湿沙层积法解除巴山榧树种子休眠,但巴山榧树种子萌发率较低。因此,巴山榧树种子休眠的具体原因及其解除方法有待进一步研究。
3.3 种子大小对种子萌发和幼苗生长的影响
许多研究表明,种子大小能够影响种子的萌发能力和幼苗的建成,较大的种子相对较小的种子具有较高的萌发率,产生更大更高的幼苗[17,25]。在本研究中,巴山榧树3个地理种群的种子萌发率和幼苗苗高与种子大小顺序表现一致。相关性分析显示,种子萌发率与种子重量呈正相关(r=0.988 1)。柯文山等[17]发现,四川大头茶(Gordoniaacuminata)种子萌发率还与纬度有关,南方种子萌发率相对较北方低。巴山榧树种子萌发率表现出与此相似的规律,来自北方(甘肃武都)种群的萌发率最高,而南方(重庆南川)种群的萌发率最低,种子萌发率与纬度正相关(r=0.778 7)。同时,巴山榧树种子萌发率与种子重量、纬度的相关性均未达到显著水平,推测种子萌发率除受种子大小和纬度的影响外,还可能与采种母树年龄及其生境因子的影响有关。巴山榧树幼苗苗高与种子重量呈正相关(r=0.997 8),达到显著水平。由于巴山榧树采种困难,本研究的样本量偏少,可能对结果分析有一定影响。
3.4 巴山榧树繁育展望
野外调查发现,巴山榧树天然种群中结实植株稀少,种子早落,且遭受鸟类、松鼠等动物的取食,种子成熟时数量少。种子萌发试验表明,巴山榧树种子萌发率低。这些因素可能是导致巴山榧树天然种群中幼苗稀少,种群更新不良的原因,同时也给种子育苗工作造成了一定困难。今后在进一步摸索巴山榧树种子育苗技术的基础上,应积极开展扦插繁殖、组织培养等无性繁殖试验,扩大巴山榧树种苗生产规模,为巴山榧树种质资源保护和种群恢复奠定基础。
[1]于永福.中国野生植物保护工作的里程碑:国家重点保护野生植物名录(第一批)出台[J].植物杂志,1999(5):3-11.
[2]周先容,余岩,周颂东,等.巴山榧树地理分布格局及潜在分布区[J].林业科学,2012,48(2):1-8.
[3]周先容,张薇,何兴金,等.巴山榧树(Torreyafargesii)资源及其保护[J].东北林业大学学报,2012,40(2):42-46.
[4]郑万钧,傅立国.中国植物志(第七卷)[M].北京:科学出版社,1978:457-467.
[5]周先容,周颂东,何兴金,等.大娄山区巴山榧树遗传多样性的RAPD分析[J].东北林业大学学报,2011,39(2):24-27.
[6]马俊,陈发波,周先容,等.不同地理种群巴山榧树及近缘种叶绿体trnL-trnF序列的分析[J].基因组学与应用生物学,2014,33(5):1 053-1 058.
[7]周先容,尚进,陈发波,等.巴山榧树及近缘种的psbA-trnH序列分析[J].西部林业科学,2015,44(1):16-21.
[8]汪建华,周先容,何兴金,等.金佛山巴山榧树灌丛群落的种类组成及结构分析[J].西南大学学报(自然科学版),2013,35(7):32-38.
[9]汪建华,周先容,尚进,等.金佛山巴山榧树灌丛群落主要木本植物种群生态位特征[J].生态学杂志,2014,33(5):1 135-1 141.
[10]汪建华,周先容,尚进,等.金佛山喀斯特山地灌丛巴山榧树群落特征[J].四川师范大学学报(自然科学版),2014,37(3):389-395.
[11]金珊珊,李建辉,金则新,等.濒危植物长叶榧种子化学成分分析[J].种子,2008,27(3):22-25.
[12]刘丽丽,汪恩锋,李建辉.濒危植物长叶榧种子形态变异研究[J].种子,2012,31(1):31-33.
[13]韩建伟,张智勇,王恩茂,等.大别山五针松种子特性及促进种子萌发的研究[J].中国农学通报,2014,30(1):5-10.
[14]郑国增.降香黄檀播种及幼苗生长规律研究[J].福建林业科技,2015,42(1):123-125.
[15]周先容,尚进,江波,等.大木山自然保护区植被类型初步研究[J].安徽农业科学,2008,36(5):1 837-1 839.
[16]张世挺,杜国祯,陈家宽.种子大小变异的进化生态学研究现状与展望[J].生态学报,2003,23(2):353-364.
[17]柯文山,钟章成,席红安,等.四川大头茶地理种群种子大小变异及对萌发、幼苗特征的影响[J].生态学报,2000,20(4):697-701.
[18]李晓洁,徐化成.白皮松种子发芽习性及其种源变异的研究[J].林业科学,1989,25(2):97-105.
[19]黄雪方,金雅琴,李冬林.乌桕不同种源种子性状的地理变异[J].西南林业大学学报,2011,31(4):44-48.
[20]蔡永立,王希华,宋永昌.中国东部亚热带青冈果实形态变异的研究[J].生态学报,1999,19(4):581-586.
[21]杨期和,叶万辉,宋松泉,等.植物种子休眠的原因及休眠的多形性[J].西北植物学报,2003,23(5):837-843.
[22]王昌腾.野生长叶榧生物学特性与保护研究[J].林业实用技术,2005(10):6-7.
[23]陈海云,杨文仙.濒危植物珙桐的种子育苗技术[J].福建林业科技,2011,38(3):110-112.
[24]史忠礼,赵同芳.香榧种子休眠的研究[J].植物学报,1973,15(2):279-280.
[25]武高林,杜国祯.植物种子大小与幼苗生长策略研究进展[J].应用生态学报,2008,19(1):191-197.
Research on Seed Characteristics and Seedling Growth Law ofTorreyafargesii,an Endangered Plant in China
JIANGBo1,2,ZHOUXianrong1,2,GONGLian1,QINMingyi1,2,SHENGYali1,ZHENGYonghong1,LIZhi1
(1.College of Life Science and Technology,Yangtze Normal University,Fuling Chongqing 408100,China;2.Research Center for Development and Utility of Unique Resources in the Wulingshan Region,Fuling Chongqing 408100,China)
Based on the indoor and field experiments on three geographical populations ofTorreyafargesii,the morphological characteristics of seeds,germination characteristics and seedling growth law are analyzed.The results indicate that a) to a certain extent,there is variability on seed morphological characteristics within every population and among populations,Chongqing Nanchuan’s population variation is larger,but Gansu Wudu’s relatively smaller.b)The seed size of three populations varies gradually,and the sequence is,Gansu Wudu’sgt;Sichuan Wanyuan’sgt;Chongqing Nanchuan’s.There are significant or very significant differences in the length,the width and the weight of the seed between the populations.There is significant difference in the length-width ratio between the Chongqing Nanchuan’s population and the Gansu Wudu’s population.The seed shape of Chongqing Nanchuan’s population trends elliptical,yet the Gansu Wudu’s spherical.c) On the seed ofT.fargesii,there are following characteristics,dormancy phenomenon,low germination rate,“S” type growth curve on the height of annual seedling.The seed germination rate and the seedling height of the 3 populations are consistent with the sequence of seed size,and the seed size has a significant effect on the seed germination and the seedling growth.So we can strengthen the protection and breeding ofT.fargesii.
Torreyafargesii; seed characteristics; seedling raising trials; seed germination;seedling growth
2016-06-25
国家自然科学基金项目(编号:31470568);重庆市自然科学基金项目(编号:cstc 2012 jjA 00006);长江师范学院科研创新团队建设计划资助项目(编号:2014 XJTD 06)。
江 波(1974—),男,重庆忠县人;讲师,主要从事植物学研究;E-mail:jblmz74@126.com。
周先容(1968—),男,四川渠县人;教授,主要从事植物学和生态学研究;E-mail:zhouxianrong@yznu.cn。
10.16590/j.cnki.1001-4705.2016.10.014
S 791.53
A
1001-4705(2016)10-0014-05
