不同含油量文冠果种子透射电镜显微结构观察
2016-11-25朱桐武创李琳琳
朱桐+武创+李琳琳


摘 要:文冠果(Xanthoceras sorbifolia Bunge)是我国所特有的一种优质的木本油料树品种,是具有发展潜力的生物能源物种,具有多方面使用价值。本试验主要针对能源植物文冠果种子中的油体进行透射电镜观测,选取不同区域栽培、含油量差异较大的文冠果种子为试材,对其油体的显微结构进行观测。结果显示,油体形状多呈球形和椭球形,少部分为不规则形状;分布数量与含油量、出油率呈正相关关系,高含油量种子中含有的油体数量较多,分布密集。
关键词:文冠果;油体;显微结构
中图分类号:S565.9 文献标识码:A DOI 编码:10.3969/j.issn.1006-6500.2016.10.001
Abstract:Wood of Shinyleaf Yellowhorn (Xanthoceras sorbifolia Bunge), wood of Shinyleaf Yellowhorn genera, single species, was a species of woody oil tree species endemic to china, This experiment selected different cultivation area of crown fruit mature seeds to test. The results showed that the oil bodies were spherical and ellipsoidal, were not uniform in size, in the cells arranged tightly focused; The oil content, oil yield efficiency and crown fruit under the microstructure of body size, quantity, positively correlated relationship. The larger quantity of high oil content of seed oil bodies and small oil body gathered in lard body forms around the oil concentration areas.
Key words:Xanthoceras sorbifolia Bunge;oil body;microstructure
文冠果属无患子科文冠果属,为落叶科灌木或小乔木。文冠果的果实为蒴果,每果有8~10粒种子。种子扁球形,暗褐色,种仁白色[1]。种子含油量很高,是我国特有的高油料作物。油体是植物种子的贮油细胞器,呈现球形体,直径为0.5~2.5 μm,一般积累于子叶、胚乳或盾片中。油体储存的能量利于种子萌生,且助力植株生长。其体形、数量因植物种类的不同而存在差异,影响因素为养分和生长条件。即使同一粒种子,油体大小也会因存在于不同组织中而产生差异[2]。一直以来油体与含油量关系都深受大家的关注[3],而很少有关于文冠果油体的相关报道。本试验采用了透射电镜技术,观察油体形态、大小,为文冠果油体研究提供基础性实验性的参考,同时为提取文冠果油或生物柴油的研究提供参考。
1 材料和方法
1.1 试验材料
试验试材为文冠果(Xanthoceras sorbifolia Bunge),是我国特有的一种优良木本实用油料树种。本试验选取种植于江苏沐阳、新疆自治区乌鲁木齐市奇台县文冠果成熟的种子。
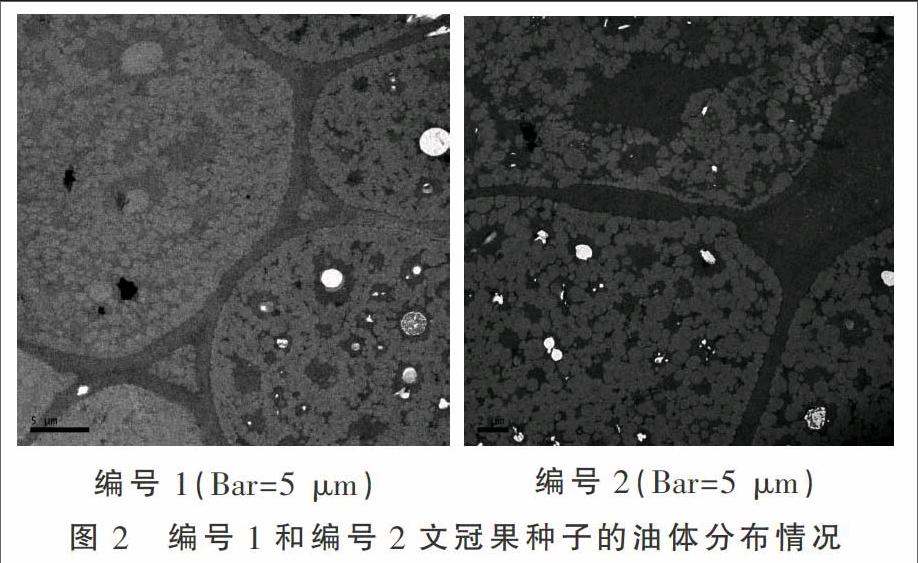
文冠果种子存在明显差异,根据文冠果形态特征和表1中的含油量、出油率,筛选出差异较大的两个种子作对比(图1):编号1和编号2(编号1三种指数都为大,编号2各指数都为小)。
1.2 透射电镜方法
文冠果种子去种壳,切割成1 mm3的组织块,用戊二醛固定,经德国 Leica EM AMW 微波组织处理机处理样品,包埋聚合,再通过德国Leica EM FC7 UC7超薄切片机对聚合块进行修块,后用双重染色法进行染色。
2 结果与分析
2.1 不同含油量文冠果种子内油体的差异性
不同含油量文冠果种子的含油量存在差异,油体分布也有差异。通过透射电镜切片的显微结构可以看出,高含油量种子,油体总体数目多,排列分布密集,大油体和小油体的集聚呈现出不规则的密集区域,分布在细胞中(图3)。而低含油量种子在细胞中出现相同规律,不同之处在于油体的总量明显小于高含油量种子,大油体小油体的聚集密度略小,细胞内出现部分空白区域(图2)。
2.2 放大后油体的观察
以编号1的文冠果种子进行放大倍数后的显微结构观察:(A)文冠果种子大油体(箭头)和部分正在融合的小油体(三角);(B)文冠果种子质体中形状不规则的油体(箭头)。油体虽然多为球形体,但也有少部分不规则形状;(C)文冠果种子油体从细胞壁向内扩充生长;(D)文冠果种子油体数量多,连成一片区域而形成油,充满整个细胞(箭头);(E)文冠果种子液泡中的油(箭头),质体中的油(三角),细胞内存在大量油体;(F) 油体蛋白(箭头)覆盖于油体表面。
2.3 油体分析
2.3.1 油体的数量和形态观察 油体多呈球形和椭球形,大小不均一,细胞内油体的大小有明显的差异性,小油体数量较多而大油体数量较少,小油体会逐渐融合成大油体。油体在细胞中的排列疏紧程度,随处在不同部位而产生差异,含量不一定相同。
2.3.2 文冠果种子中油体的发育 油贮存在种子的基本组织细胞中,由图3(C)中油体在细胞的少量分布,到图3(B)中的大量分布可知,早期细胞原生质体中出现椭球形的油体,这些油体排列松散,随机分布于原生质中。随着时间的推移和细胞形态的变化,油体数量逐渐增多,大量椭圆形、球形和不规则形状油体均匀分布于细胞质中,排列松散,直至细胞中充满油体。油体的扩增使得原生质被挤压,原生质的区域越来越小,在细胞中找到细胞核有着巨大难度。
2.3.3 文冠果种子中油体的分布 在液泡和质体中图3(E)都观察到存在着大量油体的分布,且一些小的油体会发生融合,形成一大片油的区域,且油充满细胞时呈现出多种形态。油形成于液泡和质体,且含量非常高。
3 结论与讨论
文冠果作为一种木本油料植物,它和别的油料作物一样,胚中所有细胞都积累油。起初在细胞壁和细胞核的周围发现油体的分布;随后扩增的油体逐渐向细胞中央推移;随后大量扩增,直至整个细胞充满油体。细胞中油体的形态多呈现椭球形、但也有不规则形状。由于处于不同位置,油体在细胞中的排列有疏有紧,油含量也不相同。油体在细胞内的分布与含油量有一定的关系,含油量越高,油体排列越繁多,面积越大。高含油量品种,油体总体数目多,排列分布密集,大油体和小油体的集聚呈现出不规则密集区域,分布在细胞中。而低含油量品种在细胞中也出现相同规律,不同之处在于油体的总量明显小于高含油量品种,密度略小,细胞内出现部分无油体空白区域。
油体蛋白大量覆盖于油体表面,调控油体生长。Naested 等[4]认为钙结合油素蛋白可能直接参与膜和油体的融合。Huang[5]、Sarmiento等[5-6]推测钙结合油素蛋白参与钙对油体融合的调节,使小油体通过融合变成成熟油体。钙结合油素蛋白结构的保守性意味着其功能可能受钙和磷酸化调控,现已证实钙离子/钙结合蛋白介导信号干涉植物生长的各个方面[7],包括植物生长发育和对生物非生物的应激反应[8-10]。因此,文冠果含油量高是由于钙结合油素蛋白、钙离子信号介导油体生长。随着时代高速发展,开发油体结合蛋白在生物技术及生物化学上的应用很有必要,价值不容小觑。
参考文献:
[1]方有海,孙林,赵凌泉,等.文冠果的生物生态学特性及其开发利用[J].防护林科技,2010 (1):96-97.
[2]TZEN J,CAO Y,LAURENT P,et al.Lipids, proteins, and structure of seed oil bodies from diverse species[J].Plant physiology,1993,101(1):267-276.
[3]程红焱,宋松泉.种子的贮油细胞器——油体及其蛋白[J].植物学通报,2006,23(4):418-430.
[4]NAESTED H,FRANDSEN G I,JAUH G Y,et al.Caleosins: Ca2+ binding proteins associated with lipid bodies[J].Plant molecular biology,2000,44(4):463-476.
[5]HUANG A H C. Oleosin and oil bodies in seeds and other organs[J]. Plant physiol, 1996, 110: 1055-1061.
[6]SARMIENTO C,ROSS J H,HERMAN E,et al.Expression and subcellular targeting of a soybean oleosin in transgenic rapeseed. Implications for the mechanism of oil-body formation in seeds[J].The plant journal : for cell and molecular biology,1997,11(4):783-796.
[7]DU L, POOVAIAH B W.Ca2+/calmodulin is critical for brassinosteroid biosynthesis and plant growth[J].Nature,2005,437(759):741-745.
[8]LECOURIEUX D,RANJEVA R,PUGIN A.Calcium in plant defence-signalling pathways[J].The new phytologist,2006,171(2):249-269.
[9]YANG T B,DU L Q,POOVAIAH B W.Concept of redesigning proteins by manipulating Calcium/calmodulin-binding domains to engineer plants with altered traits[J].Functional plant biology,2007,34(4):343-352.
[10]袁俊,李朝军.钙调蛋白在PC12细胞分化和迁移中的作用[J].华北农学报,2011,26(1):22-25.
