温度和授粉影响东北蒲公英花开闭的初步研究
2016-11-19张建周存宇费永俊等
张建 周存宇 费永俊 等
摘要:东北蒲公英(Taraxacum ohwianum Kitam.)花序重复开放3~4 d后,就永久性闭合。其授粉与否对花的寿命影响不大,但授粉过程会加速花朵闭合,减少花朵展示时间。温度和授粉均影响花朵开闭,东北蒲公英在20 ℃左右保持开放状态,低于15 ℃花序开始闭合。小花开放数量以及天气情况都会影响昆虫种类和数量,从而进一步影响花朵闭合进程;授粉过程可大大缩短花朵展示时间,平均减少4 h/d左右。传粉昆虫数量越多、温度越高,花朵闭合进程就越快。
关键词:东北蒲公英(Taraxacum ohwianum Kitam.);花开闭;温度;授粉;适合度
中图分类号:Q945.7 文献标识码:A 文章编号:0439-8114(2016)04-0931-04
DOI:10.14088/j.cnki.issn0439-8114.2016.04.030
Preliminary Study on Effects of Temperature and Pollination on Flower Opening
and Closing of Taraxacum ohwianum
ZHANG Jian,ZHOU Cun-yu,FEI Yong-jun
(College of Horticulture and Garden, Yangtze University, Jingzhou 434025, Hubei, China)
Abstract: The inflorescence of Taraxacum ohwianum Kitam. opened and closed repeatedly for 3 to 4 days, then permanently closed. Pollination or unpollination has little impact on floral longevity; but pollination accelerated flower closure thus reduced the show time. Temperature and pollination can affect flower opening and closing. The capitula opened at about 20 ℃ and closed at 15 ℃. Florets quantity and weather condition would affect the insect species and quantity, so further affect the close process. Pollination process can greatly reduce the show time(about 4 h/d). The more pollinators and the higher temperature is, the faster the closure process of flowers.
Key words: Taraxacum ohwianum Kitam.; flower-opening and closing; temperature; pollination; fitness
花朵开闭受一系列内在和外在因素的影响[1],开放时间也受众多环境因素调节,如内在与细胞延伸有关,外在受温度、光照质量和强度以及光暗持续时间等因素影响。并且花朵闭合常受内源节律调节,以保证花朵再次开放与闭合;还可能与衰败有关。环境因子控制花朵开闭的报道并不多。Iwanami等[2]报道了半支莲(Portulaca grandiflora Hook.)花朵开放受温度升高的影响;欧洲银莲花(Anemone coronaria L.)开放也受温度升高的控制[3];而牵牛花[Pharbitis nil (L.)Choisy]在低温(20 ℃)环境里促进开放,高温(30 ℃)则抑制开放[4]。此外,相对湿度和内源节律对花朵开闭也有影响[1]。在蒲公英属(Taraxacum F. H. Wigg.)植物中,温度也影响花朵开放,但光照可大大加速这一进程[5,6]。以上试验均在室内条件下完成,缺乏田间观察支持,如昆虫对花朵开闭的影响均未包括在内。为此,试验以有性生殖的东北蒲公英(Taraxacum ohwianum Kitam.)[7]为材料,在露天通过连续观察测定,探讨了温度和光照对东北蒲公英花朵开闭的影响程度、授粉是否加速花朵闭合、对花寿命产生什么影响等方面的问题,以进一步丰富花朵开闭研究内容,为植物繁育理论研究提供基础依据。
1 材料与方法
1.1 材料
植物材料为沈阳农业大学百草园中的多年生东北蒲公英,常规田间管理,于2012年5月进行各项指标测定。
1.2 方法
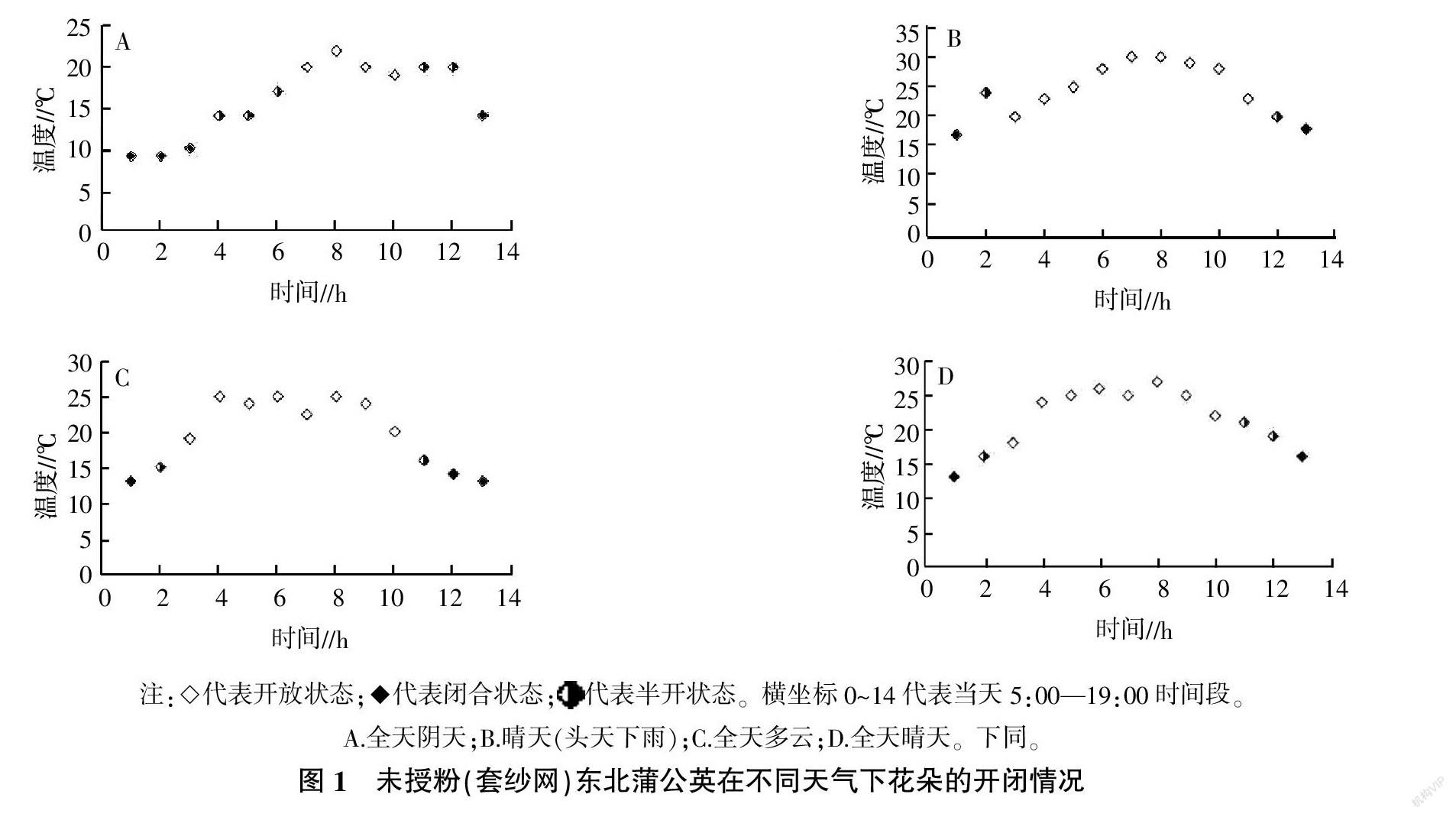
1.2.1 不同天气条件下花朵开闭观察 根据试验要求,选择4种不同天气进行自然条件和套纱网(隔绝昆虫)处理,观察花朵开闭状态。4种天气情况分别是:A.5月1日,阴天;B.5月8日,晴天,7日曾经下雨;C.5月10日,多云;D.5月15日,晴天。在试验前1 d,随机选择处于花朵开放状态的20株东北蒲公英,每株在花序上分别进行如下处理:2朵直接挂牌,2朵进行套纱网处理,隔离昆虫。试验当天全天观察,未开放时,记为未开放,有1朵开始开放,就记为半开,直至所有标记小花均开放,记为全开。闭合过程也做相同记录。每1 h观察1次。
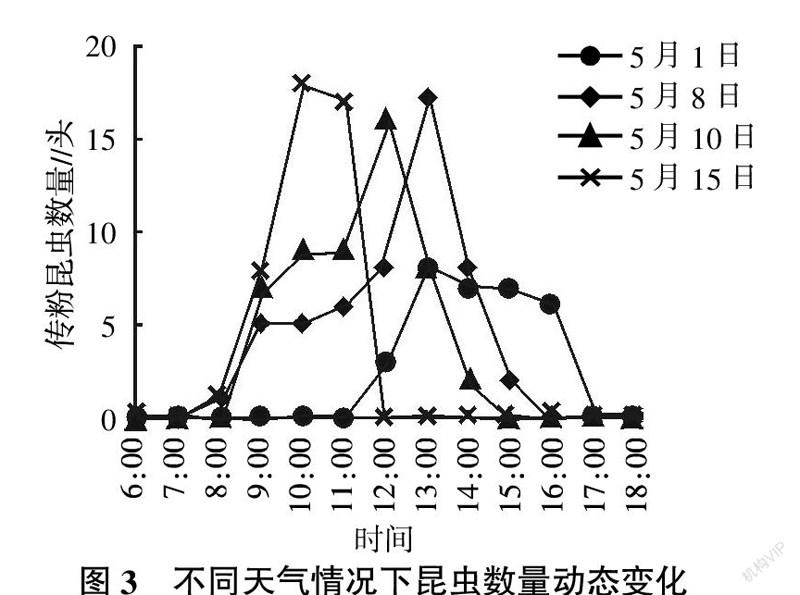
1.2.2 传粉昆虫数量动态观察 分别在4种天气的当天6:00~18:00,每隔1 h观察试验区中昆虫的数量,并进行记录,直至花序闭合停止记录。
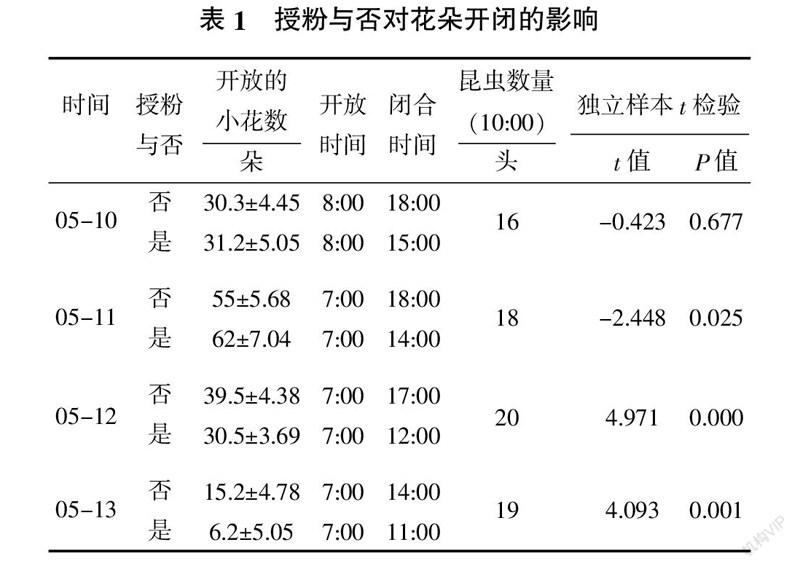
1.2.3 授粉与否对花序开放小花数的影响 选择即将开放的花序,进行自然和套纱网2种处理,每个处理20个花序,分别在5月10~13日于每天11:00统计开放的小花数,4 d定株连续观察,直至永久性闭合。
2 结果与分析
2.1 不同天气情况下授粉与否对花朵开闭的影响
2.1.1 未授粉情况下东北蒲公英花朵开闭情况 试验测定的未授粉(套纱网)东北蒲公英在不同天气下花朵的开闭情况见图1。从图1可见,天气情况对花朵开闭影响较大。在沈阳市的早春(4月底至5月初)天气蒲公英处于始花期,在阴天条件下(5月1日),气温偏低,经过3 h的半开状态,花序才全部展开,受温度影响,展现4 h后,花就开始闭合,2 h后,闭合完毕。在晴天(5月8日)花序经过1 h就全部展开,开放9 h后,陆续闭合。开放和闭合过程较5月1日均缩短。在全天多云和全天晴天(5月10日和5月15日)也具有类似现象。表明温度和光照对开闭这一进程均有影响。15 ℃左右花朵开始开放,20 ℃左右花处于完全开放状态。从开放到闭合全程大约9~11 h,在合适条件下,完全开放状态的花展示时间为8~9 h。其中光照对花朵开放有促进作用。
2.1.2 自然条件下东北蒲公英花朵开闭情况 在自然条件下东北蒲公英花朵开闭情况见图2。从图2可见,自然条件下,花朵开闭受昆虫数量的影响较大。在5月1日,昆虫数量少,其开闭过程与套纱网处理(图1-A)一致;晴天(5月8日)后盛花期来临,传粉昆虫数量急剧上升,花展示时间明显缩短;开花后期(5月10日和5月15日),气温明显升高,花朵闭合进程更快(3 h)。说明受温度和花朵开放状态的影响,传粉昆虫数量增加,传粉过程加速了花朵闭合,使花展示时间缩短4~5 h,从而可以把花展示的能量用于其他小花开放以及种子发育上,从而提高繁殖适合度。
2.1.3 不同天气情况下昆虫活动动态 试验测定的不同天气情况下昆虫数量动态变化情况见图3。从图3可见,昆虫活动受天气影响较大。开花初期(5月1日),气温较低,开花数量少,昆虫数量也偏低,且活动时间也仅在比较适宜的温度条件下。由于前一天降雨原因,5月8日温度偏低,昆虫活动高峰延迟。5月10日为多云天气,昆虫活动高峰较8日提前一个小时。5月15日天气晴朗,温度偏高,昆虫活动高峰出现在10:00—11:00,12:00时,花序全部闭合。从昆虫访花动态可知,开花阶段以及温度对花朵闭合进程影响较大,开花末期以及较高温度均可促进花朵闭合,反之亦然。
2.2 授粉对花朵开闭的影响
通过4 d定株连续观察,发现在第一天,授粉与否对小花开放数影响不大,授粉的闭合时间比未授粉的提高3 h,这是因为小花连续开放导致的结果;对此进行独立样本t检验,结果显示开花数量无显著性差异。第二天,自然条件下开放的小花数较套纱网处理有显著性提高(t=-2.448,P=0.025),并且花朵闭合时间也缩短4 h。第三天,套纱网处理开放的小花数较自然条件下多,并达到极显著差异(t=4.971,P=0.000),授粉的比未授粉的开放时间缩短了5 h。第四天,套纱网处理的还继续开放,但开花数明显降低,且花朵展示时间大大缩短,表明未授粉花序可适当延长花朵展示时间。而自然条件处理的花序,部分已永久性闭合,只有小部分还在开放,但小花数极显著降低(t=4.093,P=0.001)。通过表1发现,昆虫授粉加速了花朵闭合,在花朵展示时间上,授粉的花比未授粉的花缩短了15 h。
3 讨论
试验结果表明,蒲公英存在2种类型的花朵闭合现象,这就是暂时性闭合和永久性闭合。温度可影响花朵开放状态,而授粉可加速花朵闭合,对花朵展示时间影响较大;试验的观察结果与Tanaka等[5,6]报道的一致。蒲公英2种类型的花朵闭合均可促进有性生殖完成全过程,2种不同的闭合类型在多变的早春环境可提高其生殖适合度;当环境条件不适合时(低温、降雨、传粉昆虫有无),花序闭合,以避免花粉活性和柱头活性降低;而在条件适合时重新开放[8]。花朵开闭不仅受光照和温度影响,还受传粉者的影响[9]。通过一系列试验(另文报道)表明菊科的花,包括黄鹌菜[Youngia japonica(L.)DC.],在授粉3 h后就开始闭合。相反,未授粉的花保持开放增加好几个小时,直至下午晚些时候才开始闭合。
授粉之后,花会发生一系列改变,包括颜色、气味、花蜜产出、位置、萎蔫以及脱落[9]。授粉导致花朵闭合可减少因开放所耗费的能量,包括蒸腾作用的水分散失、花呼吸、花蜜产生、在逆境条件下花的保持以及保持花冠的状态和颜色等。此外,授粉导致花朵闭合可提高整株花序的展示水平,进而吸引更多传粉昆虫[10],这可以直接对未授粉花进行授粉[11],提高授粉效率[12]。Webb等[13]的研究表明,永久性闭合可保护子房免受捕食者取食,促进花粉管生长和加速受精胚珠发育。所以,花序永久性闭合可提高雄性和雌性适合度,尤其是在开花后期,这对完成受精过程有利。
目前对花朵寿命的研究成果表明,植物花朵的雄性阶段和雌性阶段表现出一种可塑性,比如麻花秦艽(Gentiana straminea Maxim.)[14]和马齿苋属(Portulaca L.)杂种[15]。基于对被子植物花寿命调查研究[16]认为,野外条件下花的寿命测定是最有可能代表这个植物种的真正寿命。东北蒲公英花寿命在田间和套纱网2种条件下差别不大,均为3~4 d,表明蒲公英花序寿命为一个定值,这与Primack[17]的描述较为一致。但花朵展示时间有明显差别;自然条件下较套纱网可缩短15 h,表明授粉可大大加速花朵闭合,这对于种子质量的影响需要进一步的研究。
参考文献:
[1] WOUTER G, VAN DOORN, UULKE VAN M. Flower opening and closure: A review[J]. Journal of Experimental Botany, 2003, 389: 1801-1812.
[2] IWANAMI Y, TSUJI T. The opening and closing movements of the flower of Portulaca grandifloraI[J]. Botanical Magazine Tokyo, 1962, 75: 443-448.
[3] HOROVITZ A, GALIL J, ZOHARY D. Biological flora of Israel. 6. Anemone coronaria L[J]. Israel Journal of Botany, 1975, 24: 26-41.
[4] KAIHARA S, TAKIMOTO A. Effects of light and temperature on flower-opening of Pharbitis nil[J]. Plant and Cell Physiologym, 1981, 22: 215-221.
[5] TANAKA O, WADA H, YOKOYAMA T, et al. Environmental factors controlling capitulum opening and closing of dandelion, Taraxacum albidum[J]. Plant and Cell Physiology,1987, 28: 727-730.
[6] TANAKA O, TANAKA Y, WADA H. Photonastic and thermonastic opening of capitulum in dandelion,Taraxacum officinale and Taraxacum japonicum[J]. Botanical Magazine Tokyo, 1988, 101: 103-110.
[7] 张 建,邢艳萍,赵 鑫,等.东北蒲公英花部综合征与繁育系统研究[J].中国农学通报,2013,29(33):214-219.
[8] BYNUM M R, SMITH W K. Floral movement in response to thunderstorms improve reproductive effort in the alpine species Gentiana algida (Gentianaceae) [J]. Amer J Bot, 2001, 88: 1088-1095.
[9] GORI D F. Post-pollination phenomena and adaptive floral changes[A]. JONES C E, LITTLE R J. Handbook of Experimental Pollination Biology[M]. Scientific and Academic Editions, New York, 1983.
[10] JONES C E, CRUZAN M B. Floral morphological changes and reproduct ive success in deer weed (Lotus scoparius Fabaceae) [J]. Amer J Bot, 1999, 86: 273-277.
[11] CASPER B B, LA PINE T P. Changes in corolla color and other floral characteristics in Cryptantha humilis (Boraginaceae) : Cues to discourage pollinators[J]. Evolution, 1984, 38: 128-141.
[12] WEISS M R. Floral color change as cues for pollinators[J]. Nature,1991,354:227-229.
[13] WEBB C J, LITTLETON J. Flower longevity and protandry in two species of Gentiana (Gentianaceae)[J]. Ann Missouri Bot Gard,1987,74:51-57.
[14] HE Y P, DUAN Y W, LIU J Q, et al. Floral closure in response to temperature and pollination in Gentiana straminea Maxim. (Gentianaceae), an alpine perennial in the Qinghai-Tibetan Plateau[J]. Pl Syst Evol, 2006,256: 17-33.
[15] KAZUO ICHIMURA, KENICHI SUTO. Environmental factors controlling flower opening and closing in a Portulaca Hybrid[J]. Annals of Botany, 1998, 82: 67-70.
[16] ASHMAN T L, SCHOEN D J. Floral longevity: Fitness consequences and resource costs[A]. LLOYD D G,BARRETT S C H. Floral Biology[M]. Chapman and Hall, New York, 1996.
[17] PRIMACK R B. Longevity of individual flowers[J]. Annual Rev Ecol Syst,1985,16:15-37.
