生态型造林与传统造林的杉木林邻体干扰效应比较
2016-11-16许忠坤徐清乾郭文清顾扬传杨建华
许忠坤, 徐清乾, 郭文清, 顾扬传, 杨建华
(1.湖南省林业科学院, 湖南 长沙 410004; 2.会同县林业科学研究所, 湖南 会同 418300)
生态型造林与传统造林的杉木林邻体干扰效应比较
许忠坤1, 徐清乾1, 郭文清1, 顾扬传2, 杨建华2
(1.湖南省林业科学院, 湖南 长沙 410004; 2.会同县林业科学研究所, 湖南 会同 418300)
为了比较杉木生态型造林和传统造林邻体对杉木基株干扰效应的差异,阐明生态型造林的优越性,避免造林地滋生大量五节芒,在不影响杉木生长的前提下解决传统造林中出现的炼山和抚育造成的水土流失、地力衰退、生物多样性减少和人工成本高等问题,为今后造林工作提供技术支撑。本研究选用生态型造林与传统造林两种类型造林8年生生长数据,采用邻体干扰指数模型的方法,研究两种造林类型邻体对杉木的干扰效应。结果表明:生态型造林地中有灌木植物22种、草本植物4种;传统造林地中五节芒占草本植物总数量的91.8%,仅有灌木植物7种、其它草本植物1种,前者林地灌木层和草本层的丰富度指数和香农维纳指数均大于后者。测得两者初始个体高第1、2年平均生长速度分别为86.8 cm/a、31.9 cm/a和94.2 cm/a、51.9 cm/a,灌木植物第1、2年的平均生长速度分别为48.7 cm/a和53.3 cm/a,草本植物的平均生长速度为55.7 cm/a;两者8年生杉木平均高和平均胸径分别为6.4 m、5.8 m和7.9 cm、8.7 cm。生态型造林地中灌木层干扰效应强于传统造林,主要干扰物种有4种,平均干扰指数为2.40;后者林地中草本层干扰效应强于前者,主要干扰物种有1种,平均干扰指数为0.42。生态型造林初始个体高较大,第1、2年平均生长速度大于灌、草植物,无缓苗期,造林后快速生长,对造林初期生长速度促进作用明显;传统造林初始个体高较小,有缓苗期,第1、2年平均生长速度小于灌、草植物,林地必需进行抚育。分别拟合两者总干扰指数与杉木基株大小、基株生长速度唐守正树高曲线,知其均呈显著负相关关系,前者抵抗邻体干扰能力强于后者,个体大小受邻体的影响较小,邻体对基株生长的整体干扰作用要小于后者。生态型造林不抚育促使杉木个体高生长较快,传统造林由于频繁抚育杉木个体胸径生长较快。生态型造林炼山为五节芒提供了良好的生长环境,前者无炼山抚育,生物多样性丰富,抑制五节芒生长。
杉木; 炼山; 干扰指数; 邻体; 基株; 生物多样性; 生态型造林; 传统造林
杉木(Cunninghamialanceolata)传统造林中炼山是人工造林在采伐迹地或宜林地上用火烧低成本来清理林地的一种营林措施,该方法使造林地干净,便于开展杉木造林施工活动,清理林地初期易于管理;但是此方法易引起水土流失、破坏生物多样性等[1],在亚热带地区滋生大量的五节芒(Miscanthusfloridulus),炼山后林地裸露为五节芒的生长提供了最佳的生长环境,而林地抚育后五节芒则会持续大量生长,直到林分达到郁闭状态,对目的树种生长极为不利,人工林幼林抚育的目的是抑制其它物种与目的树种竞争,为目的树种创造良好的生长环境[2],抚育的同时带来水土流失、地力衰退、破坏生物多样性,且耗费大量人工,抚育成本高等问题。由于植物的相对不移动性和表型可塑性,使得邻体及其行为对其个体的形态和生产力产生了强烈的负相互作用影响,即所谓的干扰效应[3],这种从个体水平出发来研究邻体竞争效应的数学模型称为邻体干扰指数模型[4]。由于灌木和草本植物与杉木幼龄之间以及杉木幼龄种内之间存在着生长竞争的关系,故本文从维护林地肥力、减少水土流失、降低抚育成本、保护生物多样性、提高杉木幼林的竞争力目的出发,采用生态型造林与传统造林比较个体竞争力,并采用前人研究的邻体干扰指数模型来研究杉木人工幼林、灌木以及草本之间的竞争关系与干扰效应,比较生态型造林和传统造林邻体对杉木干扰效应的差异,进而指导该地区杉木人工林的造林工作。
1 研究区概况
试验地位于杉木中心产区湖南省会同县林科所,其地理坐标为109°45′E、26°50′N,亚热带地区气候湿润,年均温度16.5 ℃,极端最高气温和极端最低气温分别为36.4 ℃和-4.4 ℃,年均降水量为1200~1400 mm,年均相对湿度在80%以上,年均日照1445.4 h,全年无霜期304天,海拔350~380 m,坡度10°~15°,土壤为红壤[5],前作为杉木次生林。
2 研究方法
2.1试验设计
生态型造林与传统造林在同一造林地进行,试验地立地指数为14,造林密度2 m×2 m,穴垦整地规格为50 cm×50 cm×40 cm,面积3000 m2。生态型造林比传统造林推迟1年,其营林措施只是伐倒林地上的植被,不炼山,采用同一批1年生裸根苗在容器中培育1年后造林(苗龄2年),在整个幼林期只伐除对杉木有干扰的藤本植物,不抚育;传统造林炼山清理林地,采用同一批1年生裸根苗比生态型造林提前1年,其营林措施是连续抚育4年,每年刀抚2次。两种类型造林8年生生长数据见表1。

表1 杉木人工林林分特征Tab1 ThestandsfeatureofChinesefirplantation造林方式营林措施苗龄(a)树龄(a)造林初始个体平均高(cm)基株平均胸径(cm)基株平均树高(m)传统造林炼山、刀抚4年8次183078758生态型造林不炼山、只伐除藤本植物278107964
2.2数据分析
2.2.1 基株选择和计算 本研究调查了杉木196株、灌木303株和草本植物5156株,为研究杉木与其周围植株干扰效应,以一株杉木为圆心,3 m为半径,圆内所有乔木、灌木和草本被定义为邻体,该株杉木为基株[3],选择28株杉木作为基株(生态型杉木林与传统杉木林各14株),测定基株的树高、胸径,并以造林后树高连年生长量来作为基株的生长速度。根据所测的邻体大小、基株大小和邻体到基株距离,计算邻体干扰指数,采用ForStat软件求出生长速度和邻体干扰指数间的回归方程。
2.2.2 邻体干扰指数模型 以Harper和Mack为首的植物生态学家提出了邻体效应(neighborhood effect),他们摒弃了传统的采用密度效应来探讨竞争的方法[6]。Weiner通过建立一年生植物(Polygonumminimun和P.Casadense)的邻体干扰模型,确定邻体干扰指数是邻体的数目、邻体到基株的距离和邻体中物种数量的函数;张大勇[7]、石培礼[3]等通过改进模型比较发现把Weiner模型中距离负二次幂式改成了负一次幂式,并在公式中加入了基株的个体大小S(通常用生物量表示[8]),最后拟合效果更好。指数模型为:
(1)
因此,本研究采用邻体到基株距离的负一次幂式来解释杉木人工林的邻体干扰效应。
2.2.3 生物量计算
(1) 乔木层生物量计算
杉木生物量计算模型参照李燕、张建国等《杉木生物量生物量估算模型的选择》中最优模型[9]。杉木全树生物量模型公式为:
W=0.093 0(D2H)0.803 0
(2)

(2)灌木层生物量计算
直接收获法和样方法是当前大多学者研究灌木层生物量多采用的方法,这些方法需消耗太多时间和精力,并且严重破坏生态系统,最重要的是存在着样本量和均匀性都很难满足统计学要求这一很大的不确定性。陶冶、张元明等[10]采用模型来估算生物量的方法解决了这一问题。宋于洋和胡晓静[11]、赵成义、宋郁东等[12]都把基径平方乘株高(D2H)作为生物量估测的形态指标,分别研究了梭梭(Haloxylonammodendron)单株地上生物量(aboveground biomass ,AGB)和其构件生物量。根据陶冶等[10]的研究结果可知地上生物量与地下生物量(belowground biomass,BGB)的关系公式为BGB=0.8737×AGB0.939 4,本研究选择宋于洋和胡晓静研究的地上生物量计算模型来作为计算灌木层生物量的方程。计算模型如下:
AGB=0.057 754(D2H)0.849 941
(3)
BGB=0.873 7(AGB)0.939 4
(4)

(3) 草本层生物量计算
林下草本层生物量的测定采用“单株收获法”,在基株3 m范围内按不同草本种类称取鲜重(草本种类极少),采集样品带回实验室,在80~85 ℃恒温下烘干至恒重并称重,求出干鲜重之比,最后将鲜重换算成干重,最后根据草本种类的株数估算各草本的生物量。
2.2.4 林下生物多样性 在样方内分灌木层、草本层和藤本植物,对其植物种类进行仔细调查记录,对灌木层、草本层和藤本植物按种类测定其高度、株数、盖度等信息,对林下生物多样性动态特征(物种丰富度指数、香农维纳多样性指数、均匀度指数)进行调查与分析。生物多样性动态特征值计算方法:
物种丰富度指数=S/ln(A)
(5)

香农维纳多样性指数(Shannon-Wiener多样性指数)

(6)
式中:P——物种在全部个体中的比例;
均匀度指数:
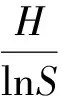
(7)

3 结果与分析
3.1杉木基株生长指标及邻体干扰指数比较
根基调查数据分别计算杉木基株生长速度、个体大小以及各干扰指数的均值(图1),可以看出,生态型造林基株个体大小平均值(11.90 kg/m2)和传统造林基株个体大小平均值(12.23 kg/m2)相差不大,比较两者基株平均生长速度发现前者(0.93 m/a)要大于后者(0.78 m/a),说明杉木基株高生长前者要快于后者;对于总干扰指数平均值却是生态型造林(4.13)明显大于传统造林(2.50);比较两者林地中各部分干扰指数平均值发现乔木层和草本层差别不大,传后者均大于前者,而灌木层差别明显(2.40>0.05),后者可忽略不计,说明前者邻体对基株的干扰效应主要来自乔木层和灌木层,后者邻体对基株的干扰效应则主要来自乔木层和草本层。
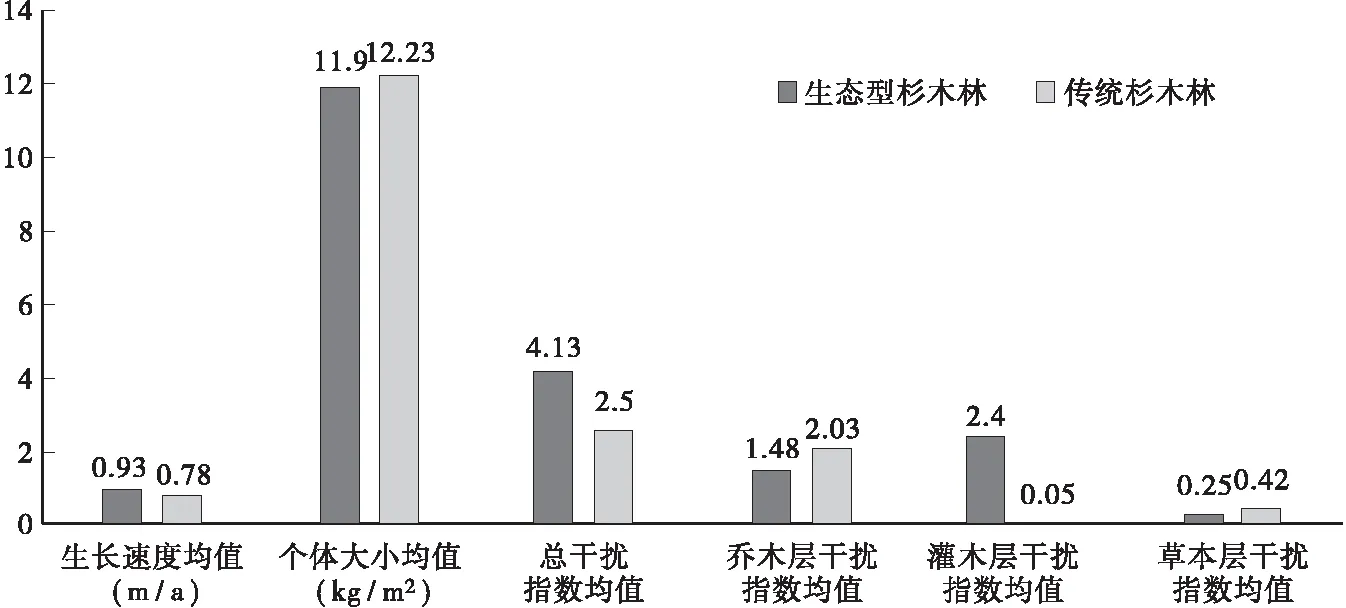
图1 杉木生态型造林和传统造林各指标均值Fig.1 The mean of each index between the ecological afforestation and traditional afforestation of Chinese fir
3.2杉木基株个体大小、生长速度与邻体干扰指数间相互关系比较
采用ForStat软件分别拟合两者杉木邻体总干扰指数与基株个体大小、生长速度的回归关系,并进行显著性检验,选择相关系数较高的拟合曲线,比较两者杉木邻体总干扰指数与基株个体大小、生长速度回归关系间的差异。
3.2.1 杉木基株个体大小和邻体干扰指数回归关系比较 拟合邻体总干扰指数与杉木基株个体大小间的回归曲线,可知生态型造林(S1)和传统造林(S2)中杉木基株个体大小和干扰指数的唐守正树高曲线[13-15]分别为:
S1=1.27+58.89/(2.0+I总),
S2=3.19+44.19/(2.0+I总)
由回归方程可知,杉木个体大小与干扰指数存在着显著的负相关,其在抵抗邻体干扰上发挥着重要的作用,即基株个体越大,受到邻体的干扰程度就越小,生长也就越快。比较两个回归曲线(图2)可知,曲线S1的回归系数b1(58.89)>曲线S2的回归系数b2(44.19)、c1(2.0)=c2(2.0),随着杉木基株个体的增大,生态型造林受邻体干扰的趋势快速下降,传统造林受邻体干扰趋势下降缓慢,说明前者在抵抗邻体干扰能力强于后者。

图2 杉木生态型造林和传统造林中个体大小与干扰指数回归曲线Fig.2 The regression curve between individual size and disturbance index in ecological afforestation and traditional afforestation of Chinese fir
3.2.2 杉木基株生长速度和邻体干扰指数回归关系比较 模拟杉木基株生长速度与邻体干扰指数间的回归曲线,可得出生态型造林(Y1)和传统造林(Y2)中杉木的生长速度和邻体干扰指数的唐守正[13]树高曲线分别为:
Y1=0.49+2.48/(2.0+I总),
Y2=0.67+0.40/(0.52+I总)
石培礼等[3]研究表明邻体的数量、大小和距离基株的距离均对基株的生长产生影响,主要表现在邻体越多,个体越大,离基株越近,基株受邻体干扰程度越大,生长速度相应的就越慢。研究杉木基株的生长速度和邻体干扰指数的回归关系,可知生长速度和干扰指数之间呈负相关。比较两个回归曲线(图3),曲线Y1回归系数b1(2.48)>曲线Y2回归系数b2(0.40)、b1/c1(1.24)>b2/c2(0.77),可知随着基株生长速度的增快,生态型造林受邻体干扰指数的影响下降明显,传统造林受影响趋势下降平缓。因此表明生态型造林邻体对基株生长的整体干扰作用要小于传统造林,表明前者林地中杉木基株生长速度快于后者。

图3 杉木生态型造林和传统造林中基株生长速度与干扰指数回归曲线Fig.3 The regression curve between growth rate and disturbance index in ecological afforestation and traditional afforestation of Chinese fir
3.3杉木基株第1、2年生长速度和初始个体高的回归关系比较
采用ForStat软件拟合杉木第1、2年生长速度和杉木初始个体高的回归曲线,并进行显著性检验,选择相关系数较高的拟合曲线,可得出杉初始个体高和第1年、第2年生长速度的唐守正树高曲线分别为:
Y1=122-2883/(2.0+H1),
Y2=121.3-2219.3/(2.0+H2)

从回归方程可知,杉木第1、2年生长速度和初始个体高之间呈显著正相关,表明初始个体越高生长速度越快,在竞争中生存下来的可能越大。图4、图5左边部分分别代表传统造林初始个体高对第1、2年生长速度的促进趋势,图4、图5右边部分分别代表生态型造林初始个体高对第1、2年生长速度的促进趋势,生态型造林地中杉木生长速度在造林初期明显大于传统造林,传统造林第1年生长速度较小,第2年开始快速生长,这也符合杉木的生长特性,即传统造林是采用1年生裸根苗造林,有缓苗期,造林后第1年生长缓慢,第2~3年开始进入快速生长期,初始个体高对生长速度的促进作用不明显;生态型造林是2年生容器苗造林,初始个体较高,无缓苗期,杉木苗上山造林后即进入迅速生长阶段,初始个体高对生长速度促进作用明显。测得两种类型造林地杉木第1、2年平均生长速度分别为86.8 cm/a、31.9 cm/a和94.2 cm/a、51.9 cm/a,灌木植物第1、2年的生长速度分别为48.7 cm/a和53.3 cm/a,草本植物的平均生长速度为55.7 cm/a,前者第1、2年平均生长速度大于灌木植物和草本植物,后者小于灌木植物和草本植物,为了使杉木生长不受影响传统造林需不断抚育,生态型造林则不需抚育。综合以上可知,生态型造林初始个体高对杉木第1、2年生长速度的促进作用强于传统造林。

图4 杉木第1年生长速度与初始个体高回归曲线Fig.4 The regression curve between the first year of growth rate and initial individual high of Chinese fir
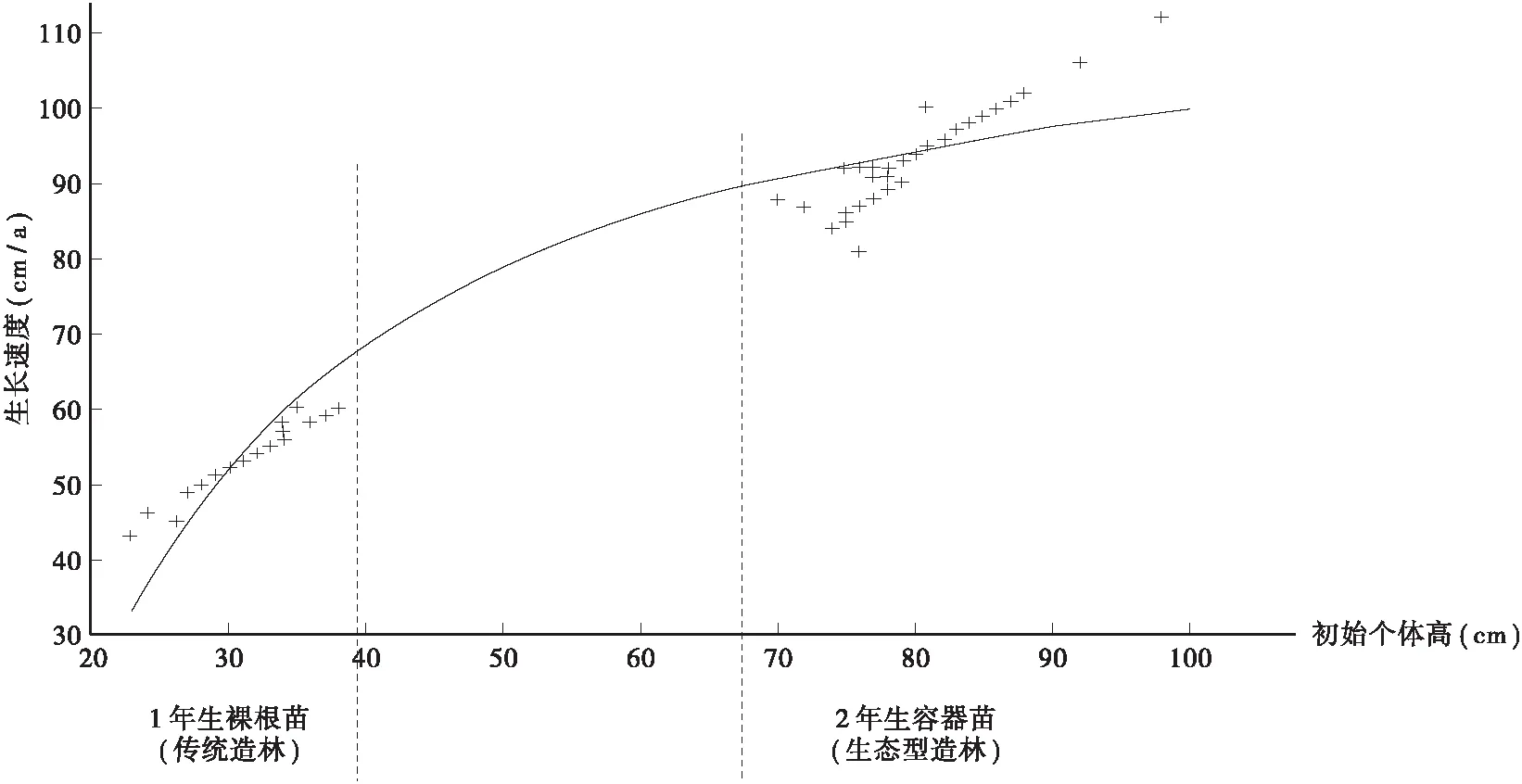
图5 杉木第2年生长速度与初始个体高回归曲线Fig.5 The regression curve between the second year of growth rate and initial individual high of Chinese fir
3.4灌木层、草本层对杉木基株的邻体干扰效应比较
3.4.1 灌木层对杉木基株邻体干扰效应比较 通过计算灌木层各个物种对基株的干扰指数的大小,找出各个基株灌木层的主要干扰物种,确定灌木层对杉木基株干扰效应的主要物种。最后比较生态型造林和传统造林中灌木层对杉木基株干扰效应的强弱,如表2所示。
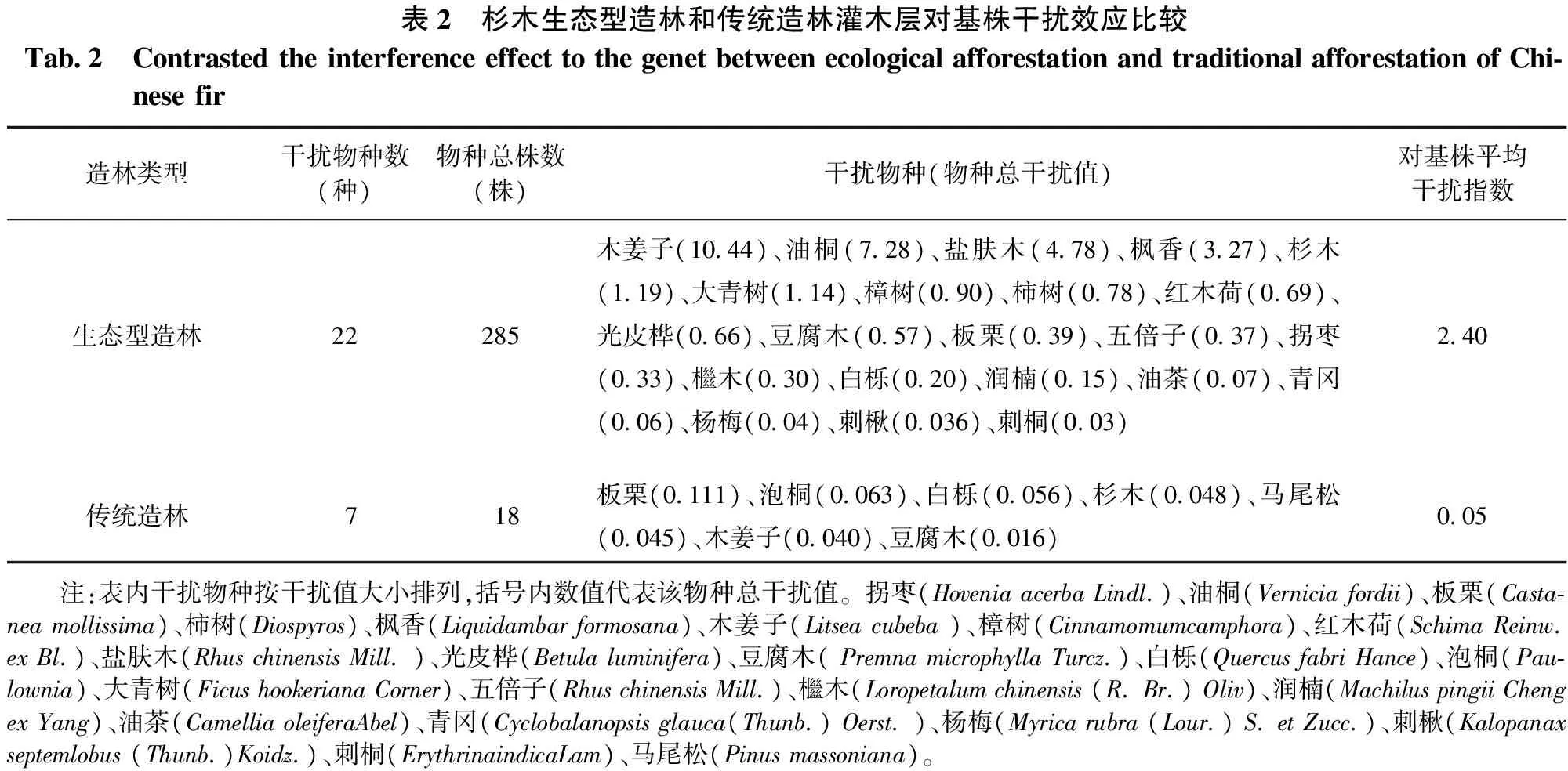
表2 杉木生态型造林和传统造林灌木层对基株干扰效应比较Tab2 ContrastedtheinterferenceeffecttothegenetbetweenecologicalafforestationandtraditionalafforestationofChi⁃nesefir造林类型干扰物种数(种)物种总株数(株)干扰物种(物种总干扰值)对基株平均干扰指数生态型造林22285木姜子(1044)、油桐(728)、盐肤木(478)、枫香(327)、杉木(119)、大青树(114)、樟树(090)、柿树(078)、红木荷(069)、光皮桦(066)、豆腐木(057)、板栗(039)、五倍子(037)、拐枣(033)、檵木(030)、白栎(020)、润楠(015)、油茶(007)、青冈(006)、杨梅(004)、刺楸(0036)、刺桐(003)240传统造林718板栗(0111)、泡桐(0063)、白栎(0056)、杉木(0048)、马尾松(0045)、木姜子(0040)、豆腐木(0016)005 注:表内干扰物种按干扰值大小排列,括号内数值代表该物种总干扰值。拐枣(HoveniaacerbaLindl)、油桐(Verniciafordii)、板栗(Casta⁃neamollissima)、柿树(Diospyros)、枫香(Liquidambarformosana)、木姜子(Litseacubeba)、樟树(Cinnamomumcamphora)、红木荷(SchimaReinwexBl)、盐肤木(RhuschinensisMill)、光皮桦(Betulaluminifera)、豆腐木(PremnamicrophyllaTurcz)、白栎(QuercusfabriHance)、泡桐(Pau⁃lownia)、大青树(FicushookerianaCorner)、五倍子(RhuschinensisMill)、檵木(Loropetalumchinensis(RBr)Oliv)、润楠(MachiluspingiiChengexYang)、油茶(CamelliaoleiferaAbel)、青冈(Cyclobalanopsisglauca(Thunb)Oerst)、杨梅(Myricarubra(Lour)SetZucc)、刺楸(Kalopanaxseptemlobus(Thunb)Koidz)、刺桐(ErythrinaindicaLam)、马尾松(Pinusmassoniana)。
由表2可以发现,生态型造林林地中干扰物种共22种285株,传统造林干扰物种仅有7种18株,可见前者林地中灌木植物在种类和数量上均比后者丰富; 其中前者中干扰强度较大的物种有木姜子、油桐、盐肤木、枫香、杉木和大青树等阳性树种,灌木植物平均干扰指数为2.40;而后者中灌木植物干扰指数较小,平均干扰指数仅为0.05。
综合以上分析可知,生态型造林和传统造林中灌木层对基株的干扰效应强弱不同,前者林地中灌木层干扰效应较强,后者灌木层干扰效应极弱,可以忽略不计。
3.4.2 草本层对杉木基株邻体干扰效应比较 通过计算草本层各个物种对基株的干扰指数的大小,确定草本层对杉木基株的主要干扰物种。最后比较生态型造林和传统造林林地中草本层对杉木基株干扰效应的强弱,如表3所示。
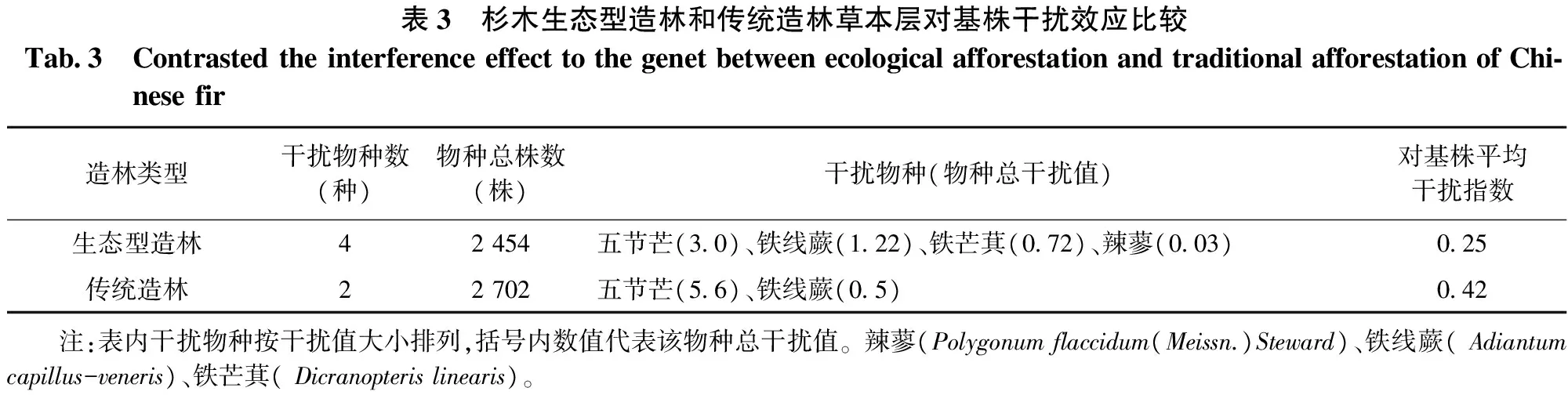
表3 杉木生态型造林和传统造林草本层对基株干扰效应比较Tab3 ContrastedtheinterferenceeffecttothegenetbetweenecologicalafforestationandtraditionalafforestationofChi⁃nesefir造林类型干扰物种数(种)物种总株数(株)干扰物种(物种总干扰值)对基株平均干扰指数生态型造林42454五节芒(30)、铁线蕨(122)、铁芒萁(072)、辣蓼(003)025传统造林22702五节芒(56)、铁线蕨(05)042 注:表内干扰物种按干扰值大小排列,括号内数值代表该物种总干扰值。辣蓼(Polygonumflaccidum(Meissn.)Steward)、铁线蕨(Adiantumcapillus-veneris)、铁芒萁(Dicranopterislinearis)。
由表3可知,生态型造林林地中草本植物种类多于传统造林,但总株数少于传统造林,其中五节芒数量占生态型造林草本植物数量的58%,占传统造林草本植物的91.8%;比较草本植物对杉木人工林的干扰指数,发现两者林中干扰强度较大的均为五节芒,分别为3.0和5.6,其它草本植物干扰强度相对较弱;草本植物对基株平均干扰指数后者大于前者(0.42>0.25)。
综合3.4.1和3.4.2分析其原因发现五节芒为风媒介进行种子传播的植物,生长发育需要开阔的空间,传统造林由于采用了炼山造林并且每年定期抚育两次,为五节芒的生长提供了有利的生长环境,使得五节芒的生长高度直逼杉木高度,同时也抑制了灌木和其它草本植物的生长,故传统造林地灌木植物数量稀疏,草本植物种类较少,五节芒对基株的干扰效应起到了极其重要的作用,在一定程度上减缓了杉木基株的生长速度;生态型造林由于未炼山未抚育,生物多样性保留较为丰富,五节芒种子无法落地进行生长,同时林地内保留的大量灌木植物的生长也相对的抑制了林地内少量残存五节芒的快速生长,故灌木植物种类和数量较多,五节芒较少,邻体对基株的干扰效应主要体现在灌木植物,而草本植物的干扰极弱。
3.5林下生物多样性比较
杉木林下植物群落组成成分在群落中所占空间不同,对群落层次结构、生态功能、动态变化等方面作用有差异。本文主要从草本和灌木植物对杉木生态型造林地和传统造林地的影响进行生物多样性分析比较。
3.5.1 草本层生物多样性比较 采用公式(5)~(7)分别计算杉木生态型造林和传统造林地中草本植物生物多样性指标,丰富度指数和香农维纳指数反映了草本植物的种类丰富程度,均匀度指数则反映了草本植物的分布情况,比较两者间的草本生物多样性差异,发现两者林地中草本层的主要干扰物种均为五节芒,可见草本层中五节芒对杉木基株的干扰作用起到了关键作用;具体结果如图6所示。
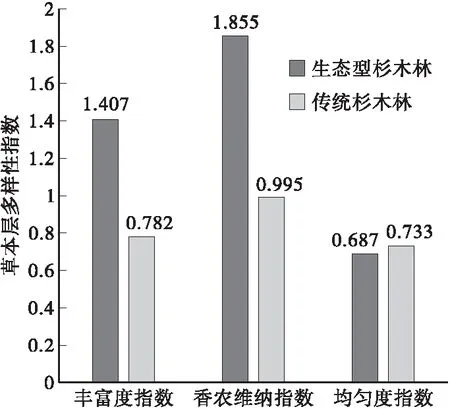
图6 两种杉木林下草本植物多样性变化Fig.6 Contrasted the changes in biodiversity of two types Chinese fir herbaceous layer
由图6可以看出,两种类型丰富度指数分别为1.407和0.782、香农维纳指数分别为1.855和0.995,生态型造林均大于传统造林,可知前者草本植物种类丰富,相比均匀度指数知前者(0.687)小于后者林(0.733),故在传统造林地中草本植物的均匀度比较明显,分析原因知传统造林中草本植物种类较少且数量很多。
3.5.2 灌木层生物多样性比较 计算两者林下灌木层生物多样性指标并比较两者间的灌木植物生物多样性差异,发现生态型造林地中灌木植物种类有22种,明显多于传统造林(7种),主要干扰物种为木姜子、盐肤木和油桐,后者林地中灌木层由于受到草本植物长芒草的抑制,灌木植物稀疏,对基株的干扰作用较弱;计算结果如图7所示。
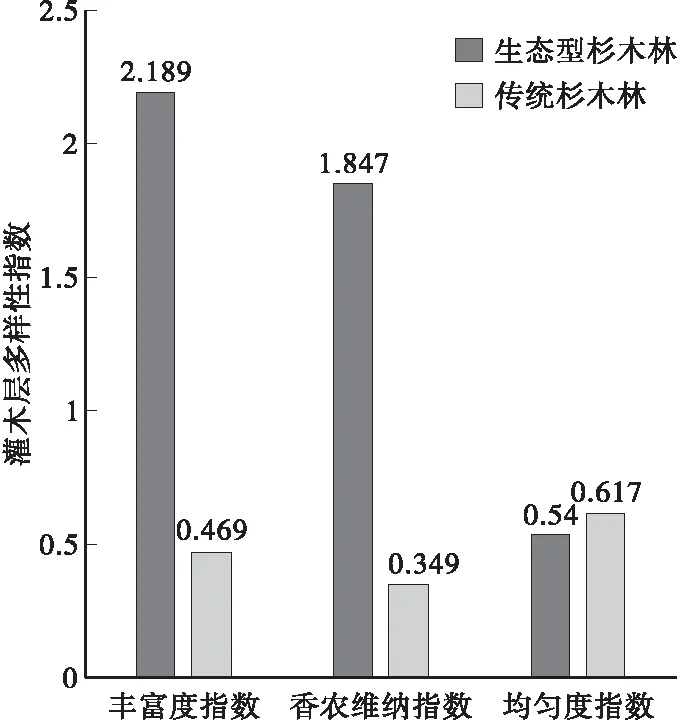
图7 两种杉木林下灌木植物多样性变化Fig.7 Contrasted the changes in biodiversity of two types Chinese fir shrub layer
由图7可以看出,两者林地中丰富度指数分别为2.189和0.469、香农维纳指数分别为1.847和0.349,生态型造林均明显大于传统造林,可知前者林地中灌木植物种类丰富,在后者林地中灌木植物种类较少,数量较小,故后者均匀度指数(0.617)大于前者(0.540)。
4 结论与讨论
(1) 杉木初始个体大小与造林后第1、2年树高生长速度呈正相关关系,测得生态型造林第1、2年平均生长速度分别为86.8 cm/a、 94.2 cm/a,传统造林为31.9 cm/a 、51.9 cm/a ,灌木植物第1、2年的平均生长速度分别为48.7 cm/a、53.3 cm/a,草本植物的平均生长速度为55.7 cm/a,生态型造林第1、2年平均生长速度大于灌木植物和草本植物,传统造林小于灌木植物和草本植物,造林初期为使杉木生长不受影响后者需不断抚育,生态型造林则不需抚育;生态型造林是2年生容器苗造林,初始个体高较大,无缓苗期,杉木苗上山后即进入迅速生长阶段,初始个体高对生长速度促进作用明显;传统造林杉木基株是采用1年生裸根苗造林,初始个体高较小,有缓苗期,前期生长缓慢,抗其它物种干扰能力弱,需要进行抚育才能正常生长。杉木基株个体大小与邻体总干扰指数呈显著负相关关系,随着基株个体的增大,生态型造林受邻体干扰的趋势快速下降,传统造林受邻体干扰趋势下降缓慢,前者抵抗邻体总干扰能力强于后者;基株生长速度与邻体总干扰指数呈负相关关系,随着基株生长速度的增快,前者受邻体总干扰指数的影响下降明显,后者受影响趋势下降平缓,前者邻体对基株生长的整体干扰作用要小于后者。
(2) 生态型造林地中生物多样性丰富,有灌木植物22种、草本植物4种,传统造林地中由于经过了炼山,为五节芒创造了适生条件,五节芒数量占草本植物总数量的91.8%,抑制了其它物种的生长,仅有灌木植物7种、其它草本植物1种;前者灌木层和草本层的丰富度指数和香农维纳指数分别为2.189、1.407和1.847、1.855,均大于后者(0.469、0.782和0.349、0.955)。
(3) 灌木植物对两者林地中基株的干扰效应强弱不同,生态型造林地内物种丰富多样,灌木种类齐全,数量较多,基株受灌木干扰效应较强,主要干扰物种有木姜子、油桐、盐肤木、枫香、杉木和大青树等阳性树种,灌木植物平均干扰指数为2.40;传统造林地内灌木植物对邻体的平均干扰指数仅为0.05,基株受灌木的干扰效应可忽略不计。
(4) 草本植物对两者林地中基株干扰效应强弱差异明显,生态型造林草本植物虽种类较多,但数量少,基株所受干扰效应较弱,主要来自五节芒和铁线蕨,草本植物对基株平均干扰指数为0.25;传统造林草本植物种类少数量多,五节芒数量占总数的91.8%,基株所受干扰效应主要来自五节芒,其它可忽略不计,对基株平均干扰指数为0.42。传统造林炼山后地面裸露,给五节芒种子提供了良好生长环境,使原来没有杂草的造林地中大量生长五节芒,要让杉木苗成活、良好生长,必需进行除草抚育,抚育后又使林地裸露,更有利于五节芒生长,反复抚育,形成恶性循环,这样恶性循环要持续到林分郁闭。生态型造林造林前不炼山,只将砍伐的剩余物,均匀铺在地面上,草种落在剩余物上面,不能与土壤直接接触,不能发芽,相比之下,杂草较少,林地上只有灌木萌芽条等植被。可见生态型造林可以抑制五节芒的生长,传统造林则为五节芒创造了有利的适生条件。
(5) 生态型造林和传统造林基株平均高分别为6.4 m和5.8 m,平均胸径分别为7.9 cm和8.7 cm,前者由于不抚育,促使杉木个体高生长相对较快,后者由于炼山和频繁抚育,使杉木个体胸径生长相对较快。
(6) 生态型造林生物多样性丰富,能够减少水土流失、维护土壤肥力、减少人工成本,解决了传统造林中五节芒大量生长的弊端,初始个体高较大,抗周围邻体干扰能力强,造林初期生长速度快。
[1] 龙昌松.炼山造林的利与弊[J].园林生态,2014,15:73-74.
[2] 李春明,杜纪山,张会儒.抚育间伐对森林生长的影响及其模型研究[J].林业科学研究,2003,16(5):636-641.
[3] 石培礼.马尾松香樟人工混交林中的邻体干扰效应[J].西南师范大学学报:自然科学版,1993,18(1):69-74.
[4] 蓝斌,洪伟,吴承祯,等.马尾松幼龄林干扰指数的研究[J].江西农业大学学报,1995,17(3):263-267.
[5] 肖复明,范少辉,汪思龙,等.湖南会同毛竹林土壤碳循环特征[J].林业科学,2009,45(6):11-15.
[6] Weiner J. A neighborhood model of annual-plant interference[J]. Ecol,1982,63(5): 1237-1241.
[7] 张大勇,赵松岭,张鹏云,等.青杄林恢复演替过程中的邻体竞争效应及邻体干扰指数的改进模型[J].生态学报,1989,9(1):52-58.
[8] 张强.植物光合-呼吸速率与个体大小以及氮元素含量间异速生长规律研究[D].兰州:兰州大学,2011.

[11] 宋于洋,胡晓静.古尔班通古特沙漠不同生态类型梭梭地上生物量估算模型[J].西北林学院学报,2011,26(2):31-37.
[12] 赵成义,宋郁东,王玉潮,等.几种荒漠植物地上生物量估算的初步研究[J].应用生态学报,2004,15(1):49-52.
[13] 唐守正,郎奎建,李海奎.统计和生物数学模型计算(ForStat教程)[M].北京:科学出版社,2009.
[14] 唐守正.长白山区树高曲线的配制[J].林业勘查设计,1980(3):26-27.
[15] 赵俊卉,亢新刚,刘燕.长白山主要针叶树种最优树高曲线研究[J].北京林业大学学报,2009,31(4):13-18.
ComparedtheinterferenceeffectfromtheneighborhoodofChineseFirbetweentheecologicalafforestationandtraditionalafforestation
XU Zhongkun1, XU Qingqian1, GUO Wenqing1,GU Yangchuan2, YANG Jianhua2
(1.Hunan Academy of Forestry, Changsha 410004, China;2.Forestry Research Institute of Huitong Country, Huitong 418300, China)
In order to compare the difference interference effect from the neighborhood of Chinese fir between the ecological afforestation and traditional afforestation, clarified the superiority of the ecological afforestation, avoided the massive growth ofMiscanthusfloridulusin the afforestation, solved the problems caused by controlled burning and upbringing under the premise of without affecting the growth of Chinese fir, for example the soil and water erosion, fertility waning, loss of biodiversity and artificial high cost etc, provide technical support for the future afforestation work. The researchSelected two kinds of plantation types of the ecological afforestation and the traditional afforestation, adopted the growth data of 8-year-old plantation and using the method of neighborhood interference index model, researched the interference effects of Chinese fir plantation from the neighborhood in the two kinds of plantation types. The results showed that:there were 22 kinds of shrubs and 4 kinds of herbs in the ecological afforestation woodland,M.floridulusaccounted for 91.8% of the total number of herbs in the traditional afforestation woodland, and there were 7 kinds of shrubs and 1 kinds of other herbs; the Richness index and Shannon-Wiener index of shrub layer and herbaceous layer in the former woodland were more than the latter woodland. Measured the initial individual height of the average growth rate in the first and second years were 86.8 cm/a, 31.9 cm/a and 94.2 cm/a, 51.9 cm/a in the former and latter respectively, the average growth rate of the first and second years were 48.7 cm/a and 53.3 cm/a in the shrubs respectively, the average growth rate were 55.7 cm/a in the herbs; the average height of the genet and the average DBH of 8-year-old Chinese fir were 6.4 m, 5.8 m and 7.9 m, 8.7 m respectively in the ecological afforestation and traditional afforestation. The interference effect of shrub layer was stronger in the ecological afforestation woodland than the traditional afforestation woodland, there were 4 kinds of the key interfering species, the average interference index of the shrub layer was 2.40; The interference effect of herbaceous layer was stronger in the latter woodland than in the former woodland, the key interfering species was 1 kind, the average interference index of the herb was 0.42. The initial individual height were higher in the ecological afforestation, the average growth rate of the first and second years were more than the shrubs and herbs, without the buffer period, rapid growth after planting, obvious improved on the growth rate in early planting period; The initial individual height were smaller in the traditional afforestation, presence the buffer period, the average growth rate of the first and second years were less than the shrubs and herbs, the woodland must be tending. Simulated the Tang Shouzheng height-diameter model between the individual size, the relationship of the curves were obviously negative correlation, the growth rate of the genet and the total interference index, the results showed that to the former woodland, the abilities to resist interference of Chinese fir were stronger, the individual size got less influences by the neighborhood, the interference effect from the neighborhood was weaker. The height growth of Chinese fir was faster because of no tending in the ecological afforestation; the growth was faster because of frequent tending in the traditional afforestation. After controlled burning the latter woodland was created the suitable conditions for the growth ofM.floridulus, and the ecological afforestation was no tending and controlled burning, the biodiversity was abundant, could inhibit the growth ofM.floridulus.
Chinese fir; controlled burning; interference index; neighborhood; genet; biodiversity; ecological afforestation; traditional afforestation
2016-06-15
“十二五”农村领域国家科技计划,杉木速生丰产林定向培育技术研究(2015BAD09B0101)。
许忠坤(1955 — ),男,湖南省慈利县人,研究员,主要从事林木栽培与育种研究工作。
S 791.27; S 72
A
1003 — 5710(2016)04 — 0028 — 09
10.3969/j.issn. 1003 — 5710.2016.04.007
(文字编校:杨 骏)
