拉萨河流域外来鱼类的分布、群落结构及其与环境的关系
2016-11-12范丽卿刘海平
范丽卿 刘海平 林 进 普 穷
(1. 西藏大学农牧学院,林芝 860000; 2. 西藏林芝地区第二高级中学,林芝 860000)
拉萨河流域外来鱼类的分布、群落结构及其与环境的关系
范丽卿1刘海平1林进2普穷1
(1. 西藏大学农牧学院,林芝 860000; 2. 西藏林芝地区第二高级中学,林芝 860000)
为揭示当前拉萨河流域外来鱼类的现状及其主要影响因素,于2010、2012及2014年对该流域的鱼类群落结构及水体环境进行了调查。共捕获4目6科17属25种鱼类,包括13种外来鱼类和12种土著鱼类。在42个采样点中的19个发现了外来鱼类,包括12处的湿地、6处的河流及1处水库采样点。外来鱼类基本都分布在下游,除了鲫(Carassius auratus)分布到了上游,鲤(Cyprinus carpio)、鳙(Aristichthys nobilis)和泥鳅(Misgurnus anguillicaudatus)分布到了中游。分布最广、数量最多的土著鱼类是拉萨裸裂尻鱼(Schizopygopsis younghusbandi younghusbandi)、东方高原鳅(Triplophysa orientalis)和西藏高原鳅(Triplophysa tibetana),最常见的外来鱼类为泥鳅、鲫和麦穗鱼(Pseudorasbora parva)。典范对应分析表明,该流域的鱼类群落同时受局域和区域环境因素的作用,二者对鱼类群落变异的总解释量为33.9%。海拔、圆石的比例和沙的比例与鱼类群落结构的关系最为紧密。今后应当开展外来鱼类的监控和清除工作,减少人为活动对河流生态系统的干扰,以促进该地区外来鱼类的防控工作及土著鱼类的保护。
外来鱼类;群落结构;拉萨河流域;入侵
外来物种的生物入侵是当今国际社会广泛关注的全球变化现象之一[1—3]。鱼类是被最频繁引入的外来水生动物之一[3]。由于水产养殖、休闲渔业、观赏鱼贸易的发展和宗教文化等原因[4—6],目前世界范围的水生生态系统都受到了外来鱼类的影响,并且大部分外来鱼类对被引进的生态系统是有害的[5]。
外来鱼类的入侵和鱼类的群落结构常常受到诸多环境因素的影响,如河段[6,7]、海拔[8,9]、水文状况[6]、底质[9]、水温[10—12]、水深[13,14]、植被[8,11]、总氮和总磷[12]、电导率[11]、盐度和溶解氧[10]等。研究表明,在环境恶劣的水体中,外来鱼类的入侵受环境的影响更大[15]。
近十年来,陆续有文章报道了在西藏的天然水体中捕获到外来鱼类,如麦穗鱼(Pseudorasbora parva)、棒花鱼(Abbottina rivularis)、鲫(Carassius auratus)、鲤(Cyprinus carpio)、鲢(Hypophthalmichthys molitrix)、鳙(Aristichthys nobilis)、草鱼(Ctenopharyngodon idellus)、泥鳅(Misgurnus anguillicaudatus)、大鳞副泥鳅(Paramisgurnus dabryanus)、鲇(Silurus asotus)和黄黝鱼(Micropercops cinctus)共9种外来鱼类分布于雅鲁藏布江中游[16,17],麦穗鱼、鲫、鲤、泥鳅、大鳞副泥鳅和黄黝鱼被捕获于尼洋河[18,19]。拉萨河流域的水体中也分布着鲫、银鲫(C. gibelio)、鲤、草鱼、麦穗鱼、泥鳅、大鳞副泥鳅、鲇和黄黝鱼等种类众多的外来鱼类[20—22]。
拉萨河流域地处青藏高原高海拔的恶劣环境中,具有独特的鱼类区系[23—26],掌握外来鱼类在整个流域的分布、鱼类群落结构及其与环境关系的资料,是该区域生态环境保护和河流管理工作的基础。然而,目前尚无关于整个拉萨河流域外来鱼类入侵情况的报道。现有的关于外来鱼类的研究只是调查了河流下游局部河汊[20]或局限在单个湿地中[21,22],且仅仅是获取了鱼类物种和数量的信息。为此,本研究的目的在于探讨引入外来鱼类后,拉萨河流域鱼类的群落结构与环境的关系,为该流域的生物多样性保护工作提供参考。
1 材料与方法
1.1研究区域
拉萨河是雅鲁藏布江最大的支流,全长551 km,河道平均坡降为2.9‰。拉萨河流域(N29°20′-31°15′,E90°05′-93°20′)面积32471 km2,海拔范围3580—7162 m,平均海拔5400 m,流域内地形起伏,大部分为山地,在山地斜坡间夹有盆地或河谷平原[27]。该流域属典型的高原温带半干旱季风气候区,具有辐射强,平均气温低,干湿季明显,年降水量小的特点,平均气温约为-1.7—9.7℃,年均降雨量约为460 mm[28]。本流域是西藏人口、工业、农牧业相对集中的区域,也是西藏政治、经济、文化、交通的中心。
拉萨河常年基流不断,主要支流有麦地藏布、桑曲、拉曲、学绒藏布、墨竹曲和堆龙曲等,整个水系呈羽毛状(图1),年径流总量为90.82×108m3,多年平均流量为288 m3/s。每年6月至9月为洪水期,11月至次年3月为枯水期[27]。
根据河谷形态和地质情况,拉萨河大致分为上、中、下游三段。上游河段从河源到桑曲汇入口,长256 km,海拔由5200降至4240 m,平均坡降3.2‰。中游河段从桑曲汇入口到直孔,长约138 km,海拔降至3880 m,平均坡降2.6‰。下游河段从直孔到雅鲁藏布江汇入口,长约157 km,平均比降约1.9‰[27]。目前,中游主河道中建有旁多水电站,中游和下游之间的主河道于2007年建成了直孔电站。流域内分布着丰富而多样的高原湿地,共有湿地面积2093 km2,占流域总面积的6.4%[28]。
1.2采样方法
2010年、2012年和2014年的春夏季,对拉萨河流域内的42个采样点进行了多次采样(图1),包括23个溪流、河流或河汊,2个水库,17个湿地,包括库塘湿地、草甸沼泽湿地、草沼泽湿地和芦苇泥炭沼泽湿地等。源头区约50 km范围由于没有路可到达,没有进行调查。上游和中游部分河段交通不便且天气寒冷,设置的采样点较少,但采样点基本覆盖了整个流域,位于上、中、下游的采样点数量分别为13、10和19个。根据陈锋和陈毅峰的调查[20]以及对渔民的访问得知,外来鱼类主要分布在流域的静水或缓流水体中。因此,本文的河流和水库采样点主要位于流速较低的河汊或近岸的水中,而非湍急河流的河道中心和水库中心。
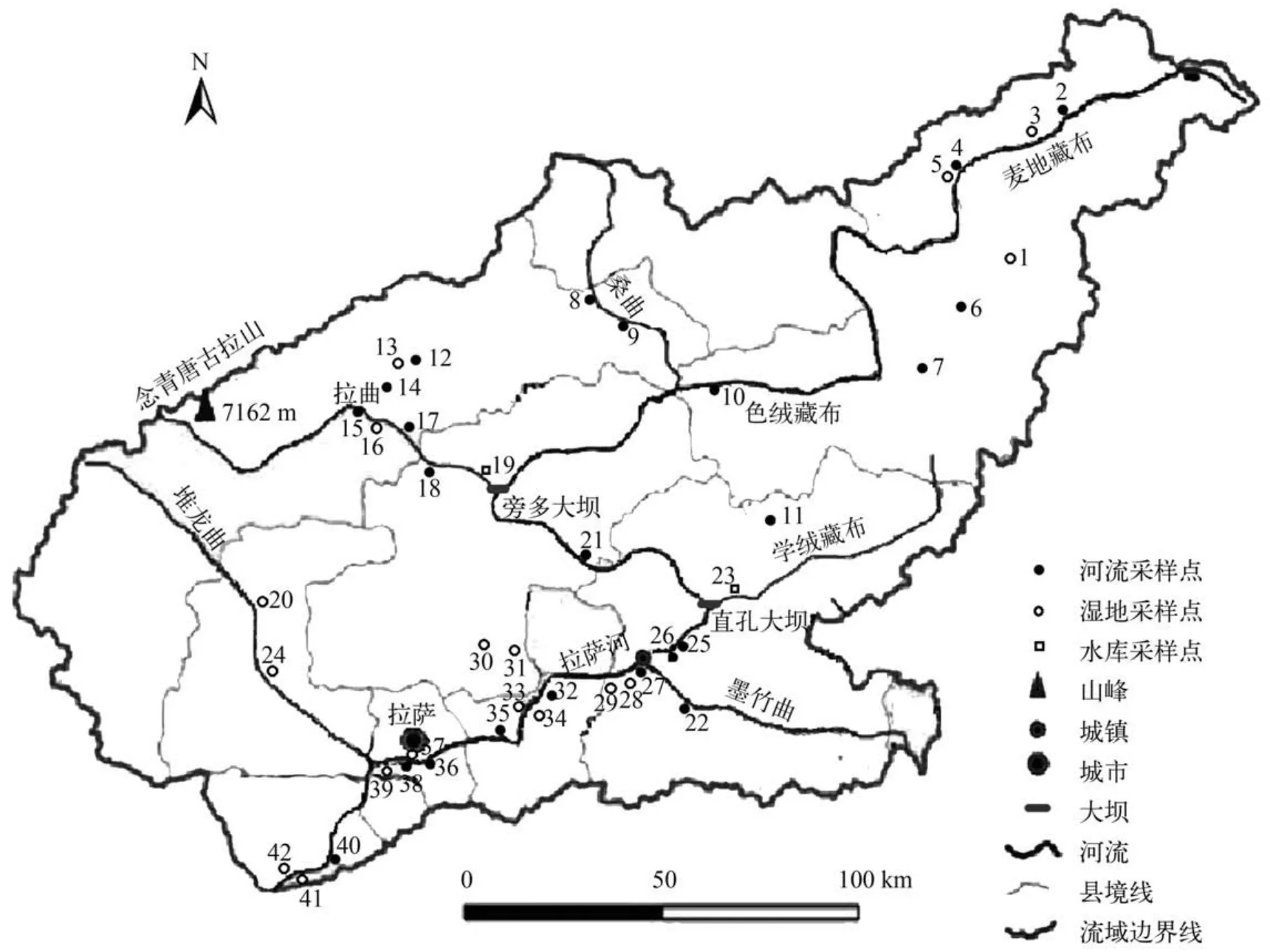
图1 研究区域及采样点示意图(图片改自Zhang等[28])Fig. 1 Map of the Lhasa River basin and sampling sites(modified from Zhang,et al.[28])
使用三层刺网(外层网目4—10 cm、内层2—6 cm,高1—1.5 m,长15 m)、定置地笼(网目10 mm,高35 cm,长5 m)结合抄网(网目5 mm)进行鱼类采样。采样时,刺网和地笼在水中放置24h后收起,大部分地点每次连续采样2 d。此外,在每个采样点使用抄网进行20—40 min的采样。其中15个采样点由于水较浅、水草茂密,仅使用地笼结合抄网的方式进行采样。
1.3环境因子的测定
测量并记录每个采样点的环境参数:经度、纬度、海拔高度(m)、所属水体的水体类型(1. 河流,包括溪流及河叉; 2. 水库; 3. 湿地)及水流类型(1. 静止; 2. 缓流; 3. 急流[8])、平均水深(0.1 cm)、植被盖度(分为5个等级:0—20%、21%—40%、41%—60%、61%—80%、81%—100%,分别赋值为1—5)、底质组成(根据Wentworth粒级划分标准,分为土、泥、沙、小砂砾、鹅卵石、圆石,分别估算它们的比例[8])。使用WGZ-1B 浊度计测量浊度(0.1 NTU),SmartTester cx-401便携式水质分析仪测量水温(0.1℃)、pH(0.1)、溶解氧(0.1 mg/L)和电导率(0.1 s/cm)。参考国家标准规定的过硫酸钾一紫外分光光度法(GBl1894-1989)和钼酸铵分光光度法(GB 11893-89)测定总氮(0.0001 mg/L)和总磷(0.0001 mg/L)。
1.4数据分析
应用CANOCO 4.5软件[29]的典范对应分析(Canonical correspondence analysis,CCA)解析环境对鱼类群落结构的影响。在分析时,采用向前引入法(The forward selection procedure),基于Monte Carlo permutation检验(n = 9999)。
只要某种鱼类被3种网具中的任何一种捕获到,则认为该物种分布于该采样点中。但由于不同采样点之间抄网的采样强度不一致,因此未将其结果用于计算鱼类群落结构。将地笼和刺网捕获的鱼类数量折算成每米网具每24h渔获量(Catch per unit fishing effort,单位捕捞努力量渔获量CPUE),多次调查的样点,计算其多次调查的平均单位捕捞努力量渔获量。把刺网和地笼调查的单位捕捞努力量渔获量相加,计算鱼类在渔获物中的比例,剔除占总渔获物比例不足0.1%的物种,共对15个种类进行了CCA分析。
在分析前,将物种数据进行lg(X+1)转换,将环境因子进行Z标准化。将经度、纬度、海拔水体类型和水流类型归为区域环境因子,其余环境因子归为局域环境因子[30,31]。使用CCA分析筛选出显著(P<0.05)的区域环境因子和局域环境因子,并对其进行以下计算:两类因子一起进入CCA分析,计算出总解释量[E+S]; 局域因子为解释变量,区域因子为斜变量,计算局域因子独立解释量[E|S]; 以局域因子为斜变量,计算区域因子独立解释量[S|E]; 区域因子和局域因子共同解释部分[E∩S]为[E+S]-[E|S]-[S|E],未解释部分为1-[E+S][31]。
2 结果
2.1鱼类分布及群落结构
本次调查共捕获到率属4目6科17属25种的鱼类,其中13种为外来鱼类(表1)。在近一半的采样点(19个)中捕获到外来鱼类,包括6个河流或河汊,2个库塘湿地,1个沼泽草甸湿地,6个草湿地,3个芦苇泥炭沼泽湿地和直孔大坝上的水库。7个采样点中外来鱼类的种数超过了鱼类总种数的一半(图2),其中一处湿地外来鱼类多达11种。下游受外来鱼类影响较大,20个样点中有16个分布有外来鱼类,而中游和上游仅有2个和1个采样点发现了外来鱼类(图2)。除了直孔水库,在其余分布有外来鱼类的采样点中,均捕获到了外来鱼类的幼鱼。除了银鲫和鳙外,捕获了其余外来鱼类的幼鱼,表明它们已经在这些水体中成功建立了繁殖种群。
在所有鱼类中,土著的拉萨裸裂尻鱼(Schizo pygopsis younghusbandi younghusbandi)、东方高原鳅(Triplophysa orientalis)和西藏高原鳅(T. tibetana)是全流域最常见的鱼类,分布最广(表1),它们在数量上分别占所有渔获物的23.1%、17.3%和16.3%,在各自分布的水体中占有的平均比例分别为24.6%(0.03%—91.2%)、21.5%(0.9%—100.0%)和26.3%(0.3%— 76.4%)。
流域下游外来鱼类的种类和数量最多,其次为中游,上游的外来鱼类最少(图2)。分布最广的是泥鳅、鲫和麦穗鱼(表1),它们也是数量最丰富的外来鱼类,分别占所有渔获物数量的4.1%、7.3%和10.6%。泥鳅的数量占其分布水体中渔获物的8.2%(0.8%—73.0%),在4个采样点中的数量超过了渔获物的20.0%。鲫是分布海拔最高的外来鱼类,在8个样点的数量超过了10.0%,是其分布水体中主要的优势种(平均占其分布水体中渔获物数量的15.5%; 0.1%—42.4%)。麦穗鱼通常是其分布的水体中最常见的鱼类(平均占其分布水体中渔获物数量的25.8%; 0.3%—73.8%),其数量在下游7个采样点中都超过渔获物的20.0%。
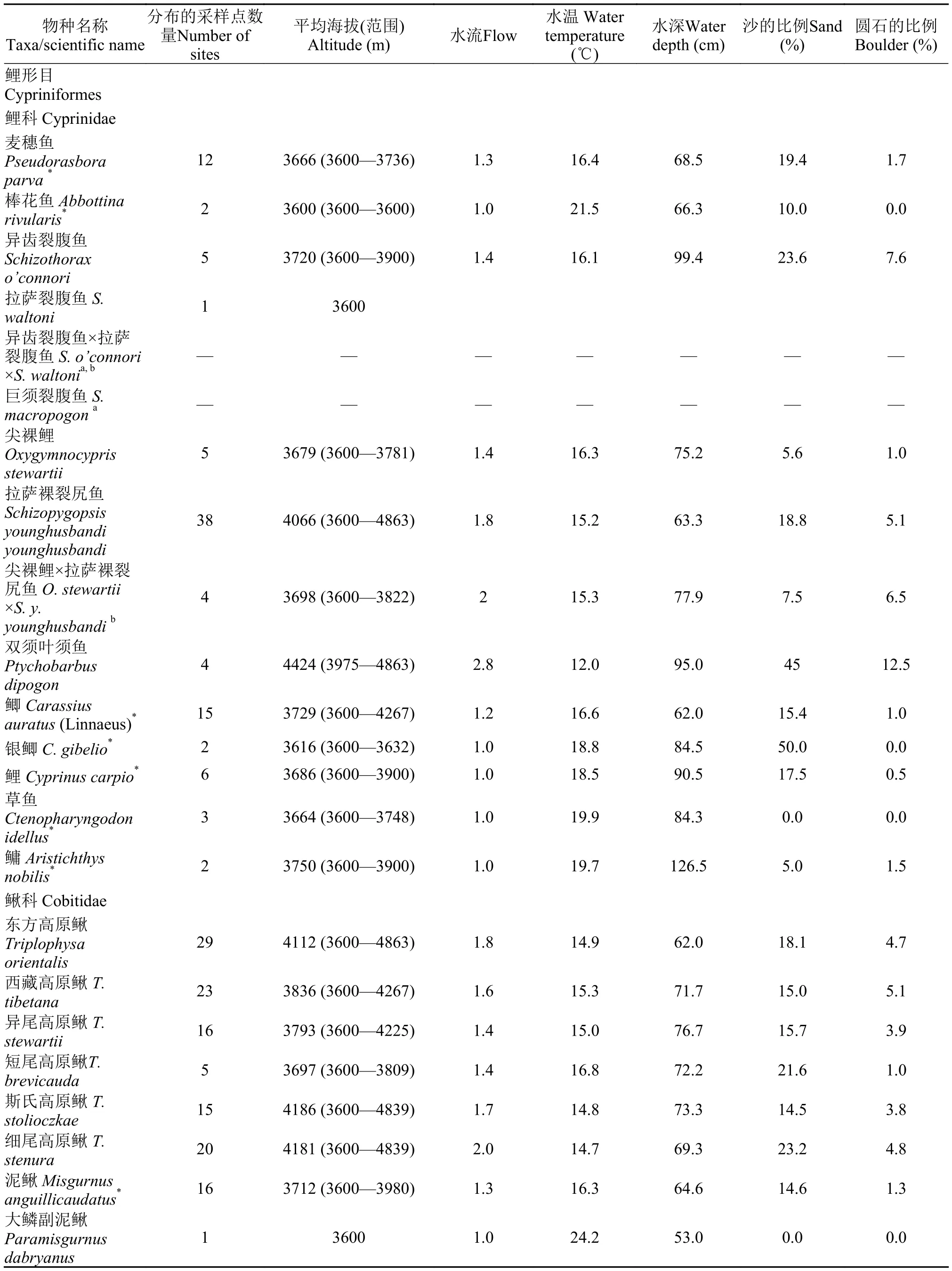
表1 拉萨河流域的鱼类及其分布水体的主要环境特征Tab. 1 Fish species and environmental parameters of sampling sites
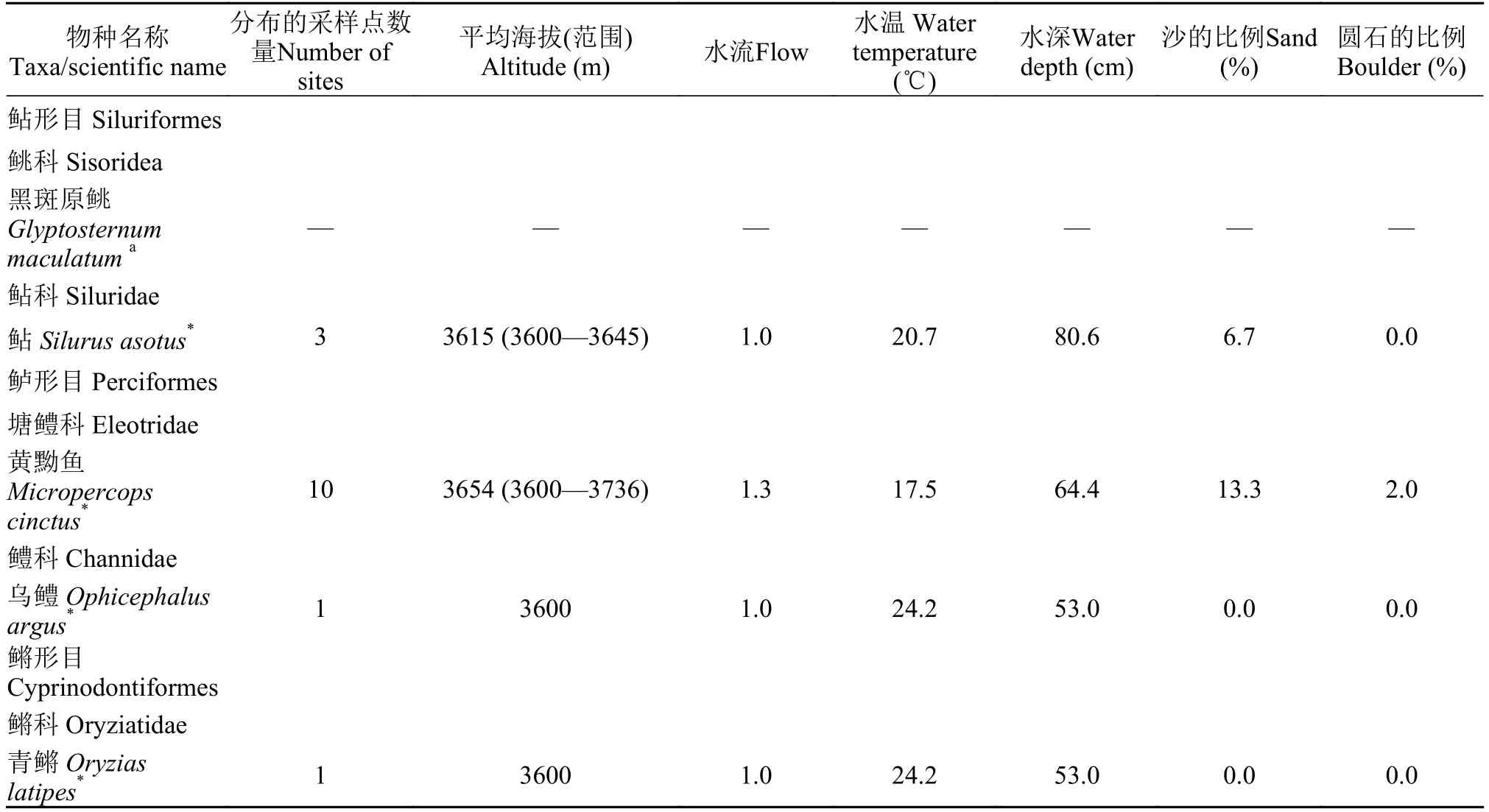
续表
本次调查只在一或两处采样点中捕获到的鱼类有棒花鱼、拉萨裂腹鱼(Schizothorax waltoni)、乌鳢(Ophicephalus argus)、银鲫、鳙和青鳉(Oryzias latipes)。其中棒花鱼和青鳉在一处芦苇泥炭沼泽湿地中有较大的数量。
2.2环境对鱼类群落的影响
拉萨河流域外来鱼类与土著鱼类分布的水体具有不同的环境特征(表1)。外来鱼类麦穗鱼、棒花鱼、鲫、泥鳅和黄黝鱼生活于相似的环境,它们常常共存于这些水体中。在裂腹鱼中,拉萨裸裂尻鱼适应整个流域的各种环境,其他裂腹鱼主要分布在水温较低的河流中。高原鳅广泛分布于流域中的各种水体,其中异尾高原鳅(T. stewartii)和短尾高原鳅(T. brevicauda)在它们分布的大部分水体中与外来鱼类共存。
CCA分析的结果表明区域环境因子中,海拔和水流类型与鱼类群落的关系最为显著(图3a)。第一轴和第二轴的特征值分别为0.518和0.176,分别解释了14.7%和5.0%的物种数据变异及54.7%和18.5%的物种-环境关系变异,前三轴共解释了22.7%的物种数据变异和84.5%的物种-环境关系变异,基本反映了鱼类群落与环境因子的关系。CCA第一排序轴与海拔(相关系数:0.7588)最相关,第二轴与水流类型相关性(0.4674)较高。
局域环境因子中,圆石的比例、水温、沙的比例和水深与鱼类的分布显著相关(图3b)。CCA第一轴和第二轴的特征值分别为0.517和0.359,分别解释了14.7%和10.1%的物种数据变异及34.8%和24.2%的物种-环境关系变异,前三轴共解释了31.0%的物种数据变异和73.5%的物种-环境关系变异。第一轴与水温相关性较高(0.5222),第二轴与圆石的比例最相关(0.7052),第三轴与水深有较大的相关性(0.5608)。
区域环境因子和局域环境因子对鱼类群落变异的总解释量为33.9%,其中局域环境因子独立解释量为16.9%,区域环境因子独立解释量为10.9%,区域和局域环境因子共同解释量为6.16%。海拔对拉萨河鱼类群落结构的影响最大,贡献了总解释量的35.0%,其次是圆石的比例和沙的比例,分别贡献了总解释量的24.2%和15.0%(图3c、表2)。
外来鱼类与土著鱼类群落在排序图中能较好的区分开来(图3)。外来鱼类在海拔较低的下游静水环境中数量较多,主要分布在水温较高、底质中较少有圆石和沙的水体中。土著鱼类拉萨裸裂尻鱼和东方高原鳅受环境影响较小,除了在下游受外来鱼类入侵严重的水体中数量较少,在整个流域的数量都比较丰富。西藏高原鳅、异尾高原鳅和异齿裂腹鱼主要分布在中下游。双须叶须鱼(Ptyc hobarbus dipogon)、细尾高原鳅(T. stenura)和斯氏高原鳅(T. stolioczkae)在上游湍急、寒冷、多沙或圆石的河流中有较高的比例。
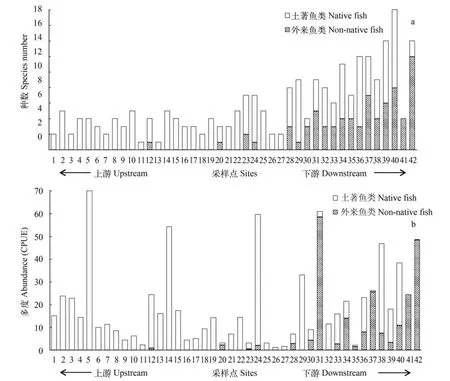
图2 拉萨河流域42个采样点外来鱼类和土著鱼类的物种数(a)和多度(b)Fig. 2 Species number(a) and abundance(b) of native and non-native fishes at sampling sites
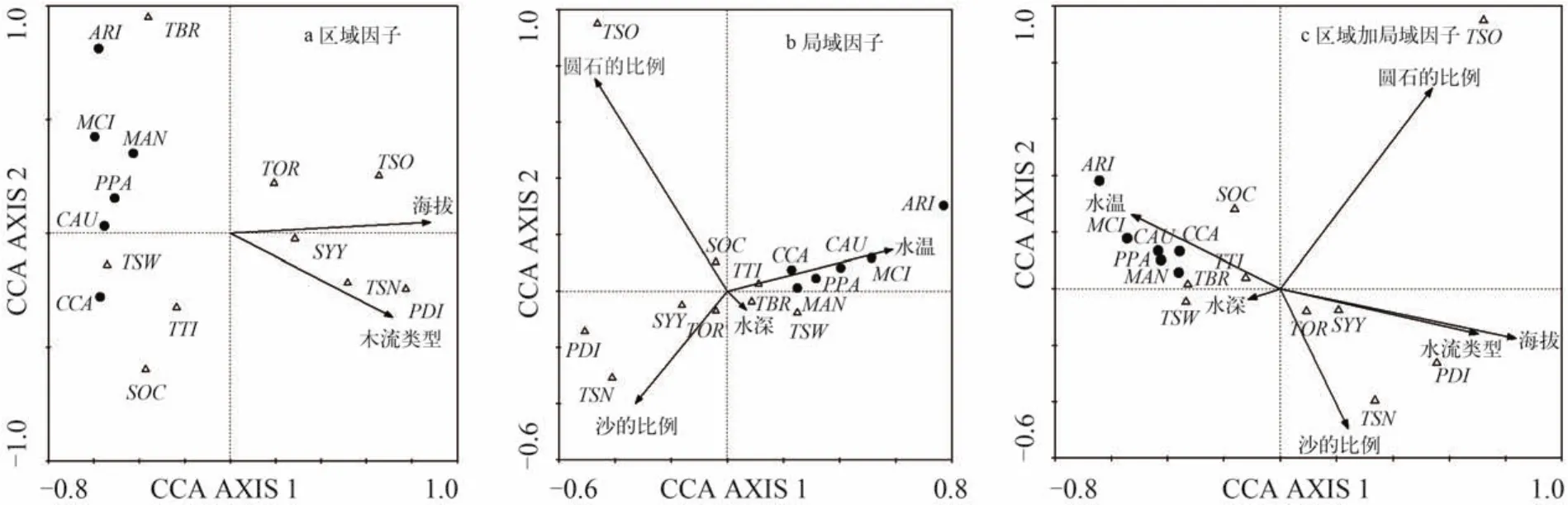
图3 拉萨河流域鱼类群落与环境因子的典范对应分析排序Fig. 3 The relationship between abundance of fishes in the Lhasa River basin and significant environmental parameters by CCA diagram analysis
3 讨论
3.1拉萨河流域鱼类的组成
本次调查捕获到了拉萨河流域15种土著鱼类中的12种,未捕捉到巨须裂腹鱼(S. macropogon)、拉萨裂腹鱼和异齿裂腹鱼的一个自然杂交种(S. waltoni × S. o'connori)和黑斑原鮡(Glyptosternum maculatum)。并且,除了拉萨裸裂尻鱼外,捕获的裂腹鱼数量很少。这主要是因为本研究缺乏对河道中心的调查,结果低估了裂腹鱼类的数量。另外一个原因是由于裂腹鱼和黑斑原鮡为当地主要的经济鱼类,从20世纪90年代开始受到较大强度的捕捞[32],种群数量下降较快。黑斑原鮡肉味鲜美,价格昂贵(市场价格大于1200 元/kg),受到极大的捕捞压力,目前在拉萨河已基本绝迹。此外,拉萨裂腹鱼和巨须裂腹鱼主要分布在雅鲁藏布江中游,在拉萨河河段的数量比较少[20]。双须叶须鱼、异齿裂腹鱼(S. o'connori)原为拉萨河裂腹鱼中的优势种[20],近年来,由于过度捕捞,资源破坏也较严重。尖裸鲤(Oxygymnocypris stewartii)为西藏自治区一级重点保护水生野生动物,被列入《中国濒危动物红皮书(鱼类)》[33],其在中下游存在一定种群,在上游数量极少[20],但近年来,由于外来鱼类和捕捞的影响,其数量也进一步下降,在市场上并不常见。拉萨裸裂尻鱼是流域的绝对优势种,仍是市场上常见的经济鱼类,但日益小型化。
高原鳅对高原环境的适应能力较强[23—25],在该流域十分常见,数量也比较丰富。但它们受到日益增加的捕捞压力和外来鱼类入侵的威胁,在河流下游的部分水体中也已经或正走向绝迹。以前,由于个体小,渔民并不捕捞它们。但近几年来,由于人们开始食用这些小型鱼类,拉萨市场上每天都有一定数量的高原鳅在被贩卖。
本次调查新捕获的外来鱼类有鳙、乌鳢和青鳉,其中鳙在雅鲁藏布江中游也有分布[16]。麦穗鱼、鲫和泥鳅是拉萨河流域最主要的外来鱼类,这与对其下游河叉[20]、拉鲁湿地[21]及茶巴朗湿地[22]的研究相似,这些外来鱼类大多对土著鱼类具有极大的威胁[22],尤其是鲫和麦穗鱼,在下游多个水体中数量众多,已造成较大影响。而这些湿地中土著鱼类的种类和数量很少(图2),这一方面可能是外来鱼类的入侵造成的,另一方面可能是人为捕捞、生境破坏等因素造成其种群数量下降,具体原因则有待进一步研究。

表2 CCA分析筛选出的区域(R)和局域环境因子(L)对鱼类群落变异的解释量Tab. 2 Variance explained by significant regional(R) and local(L) environmental parameters using CCA analysis
3.2环境对鱼类群落的影响
拉萨河流域的鱼类群落同时受区域和局域环境因子的影响(表2),这与前人的研究一致[30]。虽然研究区地理范围不大,但区域环境因子独立解释了32.2%的总解释量,表明局域的鱼类群落不仅受到局域环境的影响,同时也受到较大程度的区域环境,尤其是海拔的调节。这主要是由于部分种类,如麦穗鱼和黄黝鱼等只分布在低海拔河段,棒花鱼只分布在下游2处水体中,以及短尾高原鳅只在下游被捕获到。与陈锋和陈毅峰[20]的调查结果一致,外来鱼类主要分布于拉萨河流域下游(图2、图3),这与这些外来鱼类的引入途径有关,因为该地区的外来鱼类大多来于宗教放生及鱼塘逃逸,而放生活动及鱼塘主要分布在流域下游(未发表数据)。与中上游相比,河流下游通常受到较强的人类干扰,有利于外来鱼类的入侵[34],例如Cosumnes河的外来鱼类主要分布在农业开发程度较高和城市化更强的低地河段[8]。
诸多研究表明,底质、水温和水深是影响鱼类群落结构的重要局域环境因素,如Humber河河口的鱼类群落结构与水温有关[10]; Kootenai河上、中、下游河段的鱼类群落都受到底质组成和水深的影响[35]。拉萨河的外来鱼类群落与这些环境因子的关系不同于土著鱼类,这与这些外来鱼类的生活习性有关。它们多原产于东亚低海拔温带和亚热带水系[36],引入到拉萨河后,大多生活在底质多泥的浅水温暖水体中。少沙、少石而多泥的水体,多为静止的水域,通常水温和植被盖度更高,能够为鱼类提供更丰富的食物、栖息地及避难所,更适宜它们生活。
本次调查的环境因子只解释了33.9%的鱼类群落变异。经度和纬度等区域因子对鱼类的群落结构影响较小,这可能与研究区经度和纬度跨度较小有关。此外,各个采样点之间在植被盖度和各种水体理化因子,如溶解氧、pH和浊度等方面的差异并不大,从而降低了环境变量的解释量。今后,若能获取更多的环境及人为活动参数,如气温、降水量、坡降、土地利用情况等数据,将能更好地解释鱼类群落变异。
本次调查的结果表明,多种外来鱼类能够很好地适应拉萨河流域的环境,海拔3600 m以上的高原水体对这些外来鱼类而言是可入侵的,例如4267 m海拔的上游河流中便生活着鲫及其幼鱼。高海拔、急流及大坝原本可以阻隔外来鱼类向上游的进一步扩散,但由于人为活动的介入,外来鱼类如今已经扩散至中游的直孔水库及上游的河流中。虽然在大部分中上游并未捕获到外来鱼类,但持续存在的人类干扰,势必进一步促进外来鱼类在拉萨河中上游河段的引入和入侵成功。
地笼及刺网对鱼类种类和大小的选择性较强,如地笼对一些中上层活动鱼类的捕获效率较低,刺网的网目大小对所捕获种类的大小具有很大的影响,导致所获取的鱼类群落结构存在一定的局限。但在本次调查中,由于部分采样点植被丰富、电鱼的效率很低,因此仍然选择网具调查的方式,并通过增加网具、调查时间和调查次数来尽量弥补网具采样的不足。此外,缺乏深水区的采样,使得本文估算出的土著鱼类分布范围小于实际情况,得到的鱼类群落结构与环境关系的信息不够全面。土著鱼类,尤其是异齿裂腹鱼、拉萨裂腹鱼、尖裸鲤和双须叶须鱼等裂腹鱼类,大多生活在河流中,其分布的水深应远大于本次调查的结果,水温则应更小。
3.3对鱼类保护的启发
当前,拉萨河流域已经被全面禁渔,土著鱼类受到了比较好的保护。然而,外来鱼类已经占据流域下游的大量水体,由于人为活动的影响,今后外来鱼类的种类和数量很可能进一步增加,给土著鱼类及水生生态系统的保护和管理工作带来严峻的考验。对外来入侵鱼类进行治理、恢复流域内的鱼类群落将是一项十分艰难的工作,当务之急是在拉萨河流域下游水体开展外来鱼类清除工作[3],积极开展有关外来鱼类入侵危害的宣传教育、对民众的宗教放生活动进行合理引导,以及对天然水体的外来鱼类进行长期监测,以减少外来鱼类在该流域的进一步引入、避免外来鱼类进一步扩散。
致谢:
本研究野外工作得到西藏大学农牧学院郭其强老师、研究生孙包鹏、珍珠等的帮助以及拉萨市农业局的大力支持与帮助,中国科学院水生生物研究所何德奎老师、西藏大学农牧学院张惠娟老师对标本鉴定工作提供帮助,特此致谢。
[1]Simberloff D,Martin J L,Genovesi P,et al. Impacts of biological invasions:what's what and the way forward [J]. Trends in Ecology and Evolution,2013,28(1):58—66
[2]Vitousek P M,D'Antonio C M,Loope L L,et al. Introduced species:A significant component of human-caused global change [J]. New Zealand Journal of Ecology,1997,21(1):1—16
[3]Gozlan R E,Britton J R,Cowx I,et al. Current knowledge on non-native freshwater fish introductions [J]. Journal of Fish Biology,2010,76(4):751—786
[4]Gozlan R E. Introduction of non-native freshwater fish:is it all bad [J]? Fish and Fisheries,2008,9(1):106—115
[5]Vitule J R S,Freire C A,Simberloff D. Introduction of non-native freshwater fish can certainly be bad [J]. Fish and Fisheries,2009,10(1):98—108
[6]Moyle P B,Light T. Fish Invasions in California:do abiotic factors determine success [J]? Ecology,1996,77(6):1666—1670
[7]Adams S B,Frissell C A,Rieman B E. Geography of invasion in mountain streams:Consequences of head-water lake fish introductions [J]. Ecosystems,2001,4(4):296—307
[8]Moyle P B,Crain P K,Whitener K,et al. Alien fishes in natural streams:Fish distribution,Assemblage structure,and conservation in the Cosumnes River,California,USA[J]. Environmental Biology of Fishes,2003,68(2):143—162
[9]Grossman G D,Freeman M C. Microhabitat use in a stream fish assemblage [J]. Journal of Zoology,1987,212(1):151—176
[10]Marshallf S,Elliottf M. Environmental influences on the fish assemblage of the Humber Estuary,U.K [J]. Estuarine,Coastal and Shelf Science,1998,46(2):175—184
[11]Marchetti M P,Moyle P B. Effects of flow regime on fish assemblages in a regulated California stream [J]. Ecological Applications,2001,11(2):530—539
[12]Meador M R,Brown L R,Short T. Relations between introduced fish and environmental conditions at large geographic scales [J]. Ecological Indicators,2003,3(2):81—92
[13]Bell J D. Effects of depth and marine reserve fishing restrictions on the structure of a rocky reef fish assemblage in the North-Western Mediterranean sea [J]. Journal of Applied Ecology,1983,20(2):357—369
[14]Brown L R. Fish communities and their associations with environmental variables,lower San Joaquin River drainage,California [J]. Environmental Biology of Fishes,2000,57(3):251—269
[15]Ross S T. Mechanisms structuring stream fish assemblages:are there lessons from introduced species [J]?Environmental Biology of Fishes,1991,30(4):359—368
[16]Yang H Y,Huang D M,Xie S,et al. Status quo of fishery resources in the middle reach of Brahmaputra River[J]. Journal of Hydroecology,2010,3(6):120—126 [杨汉运,黄道明,谢山,等. 雅鲁藏布江中游渔业资源现状研究. 水生态学杂志,2010,3(6):120—126]
[17]Ma B S. Study on the biology and population dynamics of Schizothorax o'connori [D]. PH.D thesis. Huazhong Agricultural University. 2011 [马宝珊. 异齿裂腹鱼个体生物学和种群动态研究. 博士学位论文. 华中农业大学. 2011]
[18]Shen B H,Guo L. Investigation of fish resources in the Nyang River [J]. Hebei Fisheries,2008,5:51—54,60[沈红保,郭丽. 西藏尼洋河鱼类组成调查与分析. 河北渔业,2008,5:51—54,60]
[19]Zhou J,Lai J S,Du J,et al. Investigation and protection countermeasures of fish resources in Nyingchi prefecture[J]. South west China Journal of Agricultural Sciences,2010,23(3):938—942 [周剑,赖见生,杜军,等. 林芝地区鱼类资源调查及保护对策. 西南农业学报,2010,23(3):938—942]
[20]Chen F,Chen Y F. Investigation and protection strategies of fishes of Lhasa River [J]. Acta Hydrobiologica Sinica,2010,34(2):278—285 [陈锋,陈毅峰. 拉萨河鱼类调查及保护. 水生生物学报,2010,34(2):278—285]
[21]Fan L Q,Tu Y L,Li J C,et al. Fish assemblage at Lhalu Wetland:was the native fish still present [J]? Resources Science,2011,33(9):1742—1749 [范丽卿,土艳丽,李建川,等. 拉萨市拉鲁湿地鱼类现状与保护. 资源科学,2011,33(9):1742—1749]
[22]Ding H P,Qin J H,Lin S Q,et al. Exotic fishes in Chabalang wetland of Lhasa [J]. Journal of Hydroecology,2014,35(2):49—55 [丁慧萍,覃剑晖,林少卿,等.拉萨市茶巴朗湿地的外来鱼类. 水生态学杂志,2014,35(2):49—55]
[23]Zhu S Q. Synopsis of Freshwater Fishes of China [M]. Nanjing:Jiangsu Science and Technology Publishing House. 1989,6—8 [朱松泉. 中国条鳅志. 南京:江苏科学技术出版. 1989,6—8]
[24]Wu Y F,Wu C Z. The Fishes of the Qinghai-Tibet Plateau [M]. Chengdu:Sichuan Publishing House of Science and Technology. 1991,89—100 [武云飞,吴翠珍. 青藏高原鱼类. 成都:四川科学技术出版社. 1991,89—100]
[25]Bureau of Aquatic Products. Fishes and Fish Resources in Tibet,China [M]. Beijing:China Agriculture Press. 1995,10—23 [西藏自治区水产局. 西藏鱼类及其资源. 北京:中国农业出版社. 1995,10—23]
[26]Chen Y F,Cao W X. Schinzothoracinae [A]. In:Yue P Q,et al.(Eds.),Fauna Sinaica Osteichthyes Cypriniformes III [C]. Beijing:Science Press. 2000,273—390 [陈毅峰,曹文宣. 裂腹鱼亚科. 见:乐佩琦等,中国动物志:硬骨鱼纲鲤形目(下卷). 北京:科学出版社. 2000,273—390]
[27]The Team of the Comprehensive Scientific Expedition to the Qinghai-Xizang Plateau,Chinese Academy of Sciences. Rivers and Lakes in Tibet [M]. Beijing:Science Press. 1984,108—142 [中国科学院青藏高原综合考察队. 西藏河流与湖泊. 北京:科学出版社. 1984,108—142]
[28]Zhang Y L,Wang C L,Bai W Q,et al. Alpine wetlands in the Lhasa River Basin [J]. Journal of Geographical Sciences,2010,20(3):375—388
[29]ter Braak C J F,Smilauer P. CANOCO Reference Manual and CanoDraw for Windows User's Guide:Software for Canonical Community Ordination. Version 4.5. Microcomputer Power,Ithaca,New York. 2002
[30]Hoeinghaus D J,Winemiller K O,Birnbaum J S. Local and regional determinants of stream fish assemblage structure:inferences based on taxonomic vs. functional groups [J]. Journal of Biogeography,2007,34:324—338
[31]Cai Y J,Jiang J H,Zhang L,et al. Structure of macrozoobenthos in lakes along the Yangtze River and relationships with environmental characteristics [J]. Acta Ecologica Sinica,2013,33(16):4985—4999 [蔡永久,姜加虎,张路,等. 长江中下游湖群大型底栖动物群落结构及影响因素. 生态学报,2013,33(16):4985—4999]
[32]Zhang C G. Fish resources in Tibet [J]. China's Tibet,1997,5:53—55 [张春光. 西藏鱼类资源. 中国西藏,1997,5:53—55]
[33]Yue P Q,Chen Y Y. China Red Data Book of Endangered Animals(Pisces) [M]. Beijing:Science Press. 1998,158—159 [乐佩琦,陈宜瑜. 中国濒危动物红皮书(鱼类). 北京:科学出版社. 1998,158—159]
[34]Leprieur F,Beauchard O,Blanchet S,et al. Fish invasions in the world's river systems:when natural processes are blurred by human activities [J]. PLoS Biology,2008,6(2):e28. doi:10.1371/journal.pbio.0060028
[35]Smith C D,Quist M C,Hardy R S. Fish assemblage structure and habitat association in a large western river system [J]. River Research and Applications,2015,doi:10.1002/rra.2877
[36]Chen Q T,Zhen B S. Systematic Synopsis of Chinese Fishes [M]. Beijing:Science Press. 1987,126,161,166,179,180,182,199,210,223,430,457 [成庆泰,郑葆珊.中国鱼类系统检索(上册). 北京:科学出版社. 1987,126,161,166,179,180,182,199,210,223,430,457]
NON-NATIVE FISHES:DISTRIBUTION AND ASSEMBLAGE STRUCTURE IN THE LHASA RIVER BASIN,TIBET,CHINA
FAN Li-Qing1,LIU Hai-Ping1,LIN Jin2and PU Qiong1
( 1. Agriculture and Animal Husbandry College of Tibet University,Linzhi 860000,China; 2. The Second High School of Linzhi,Linzhi 860000,China)
The Lhasa River is the biggest tributary of the Yarlung Zangbo River,Tibet,China. Non-native fishes were found recently in the basin,but the distribution of non-native fishes and fish assemblage in the whole basin were poorly known. We examined fish assemblage and its relationship with the environmental factors in the basin in 2010,2012 and 2014. 25 fish species were collected,and 13 of them were non-native fishes. Non-native fishes were distributed at 19 of 42 sampling sites,including 12 wetland sites,6 river sites and 1 reservoirs above dam. Most non-native fishes occurred in the downstream with the exception of Carassius auratus in the upstream and Cyprinus carpio,Aristichthys nobilis as well as Misgurnus anguillicaudatus in the middlestream of the river. The most widely distributed and abundant native fishes were Schizopygopsis younghusbandi younghusbandi,Triplophysa orientalis and T. tibetana,and the most widely distributed and abundant non-native fishes were M. anguillicaudatus,C. auratus and Pseudorasbora parva. Canonical correspondence analysis indicated that the fish assemblage structure was impacted by both local environmental and regional environment factors,and the non-native fishes and native fishes had different relationships with the environment factors(elevation and substrate).
Non-native fishes; Fish assemblage structure; The Lhasa River Basin; Invade
Q958
A
1000-3207(2016)05-0958-10
10.7541/2016.124
2015-04-21;
2016-01-21
国家自然科学基金(31160140); 西藏自治区科技重大专项(2015XZ01G72)资助 [Supported by the National Natural Science Foundation of China(31160140); Science and Technology Major Project of Xizang Province(2015XZ01G72)]
范丽卿(1982—),女,福建大田人; 硕士; 主要从事鱼类生态学研究。E-mail:flqpig@163.com
