中国水仙锌指蛋白NtPLATZ1的克隆与表达分析
2016-11-11林江波王伟英李海明吴水金李跃森戴艺民
林江波,王伟英,李海明,吴水金,邹 晖,李跃森,戴艺民
(福建省农业科学院 甘蔗研究所,福建 漳州363005)
中国水仙锌指蛋白NtPLATZ1的克隆与表达分析
林江波,王伟英,李海明,吴水金,邹晖,李跃森,戴艺民
(福建省农业科学院 甘蔗研究所,福建 漳州363005)
【目的】 克隆中国水仙植物AT富集序列锌结合蛋白基因(NtPLATZ1),为研究该基因的生物学功能奠定基础。【方法】 利用RACE技术克隆了NtPLATZ1 cDNA全长,利用生物信息学方法分析其核苷酸及氨基酸序列,将NtPLATZ1与原核表达载体pGEX-4T-3连接,转化BL21并进行诱导表达,通过半定量PCR分析用多效唑处理后该基因在中国水仙不同生长时期叶片中的表达情况。【结果】NtPLATZ1的cDNA全长1 024 bp,包含1个642 bp的完整阅读框架,编码213个氨基酸;编码蛋白具有PLATZ superfamily蛋白保守区,与大豆(Glycinemax, AII19314.1)PLATZ蛋白序列的同源性最高,为81%。原核表达结果表明,NtPLATZ1在大肠杆菌中能有效表达。半定量RT-PCR分析表明,喷雾多效唑后,NtPLATZ1基因在中国水仙不同生长时期叶片中的表达量先上升后下降,抽葶期叶片的表达量最高;喷雾清水后NtPLATZ1基因表达量先下降后又有所回升,抽葶期和始花期的表达量低于长叶期和盛花期。【结论】 成功克隆了中国水仙NtPLATZ1基因,多效唑处理能明显诱导该基因的表达。
中国水仙;PLATZ;基因表达;锌指蛋白
锌指蛋白(zinc finger protein)是一类具有“手指状”结构域的转录因子,通过结合Zn2+来稳定一种很短的可自我折叠成“手指”的蛋白结构[1]。根据半胱氨酸(C)和组氨酸(H)残基数目、位置的不同,锌指结构域的转录因子可分为 C2H2、C2HC、C2C2、C3H、C3HC4(RING finger)和 C2HC5(LIM finger)等亚类[2]。根据锌指蛋白的结构特征或功能差异,又可以分为WRKY型蛋白、GATA型蛋白、PHD型蛋白、DOF型蛋白和TFⅢA型蛋白等[3-4]。锌指蛋白在转录和翻译水平上对基因表达进行调控,对植物种子发育[3]、花发育[5]、侧根和分枝发育[6]以及环境胁迫的应答反应[7-8]起重要作用。
PLATZ是一类新型的锌指蛋白,其具有2个锌指结构域:一个是C-X2-H-X11-C-X2-C-X(4~5)-C-X2-C-X(3~7)-H-X2-H,其半胱氨酸和组氨酸残基(CHC4H2)的数量与RING finger(C3HC4)和LIM finger(C2HC5)相同,但属于不同类型的锌指结构域;另一个是 C-X2-C-X(10~11)-C-X3-C,与GATA finger(C2C2)相似,但不同于GATA finger(C-X2-C-X(17~18)-C-X2-C);PLATZ结合于DNA序列的AT富集区域起转录抑制作用,是锌依赖的DNA结合蛋白[9]。有关中国水仙PLATZ基因的研究尚未见相关报道。
中国水仙(Narcissustazettavar.chinensis)为中国的十大名花之一,香味独特。因室内光线较弱,中国水仙在室内水养时,容易引起徒长。施用适宜浓度的多效唑能使植株矮化,株形紧凑,提高其观赏价值[10]。笔者前期通过筛选多效唑诱导中国水仙的SSH文库,获得1个cDNA片段,该片段编码的氨基酸与已知植物的PLATZ有较高的同源性。因此,本研究克隆了中国水仙PLATZ1基因cDNA序列全长,探讨该基因的生物学信息及其在多效唑处理后不同生长阶段水仙植株中的表达情况,以期为进一步研究该基因的生物学功能奠定基础。
1 材料与方法
1.1材料
中国水仙的处理参照林江波等[11]的方法,水养16 d后,每天用150 mg/L的多效唑喷雾1次,连续喷雾10次,另设喷雾清水为对照。取长叶期、抽葶期、始花期和盛花期叶片,置于-70 ℃冰箱中保存备用。
大肠杆菌(Esherichiacoli)菌株BL21(DE3)、HB101和载体pGEX-4T-3,由福建省农业科学院甘蔗研究所实验室保存;pMD19-T连接试剂盒、ExTaqDNA Polymerase、限制性内切酶、PrimeScriptTMReverse Transcriptase、RNAiso Plus和5′-Full RACE Kit,购自宝生物公司;UNIQ-10柱式DNA胶回收试剂盒,购自上海生工。
1.2总RNA的提取与cDNA的克隆
参照RNAiso Plus试剂盒说明书提取中国水仙叶片总RNA,按照PrimeScriptTMReverse Transcriptase试剂说明书进行逆转录,合成cDNA第1链。
用于5′RACE的引物序列为:5PLATZ1.5′-CTC-CCCCATCATAGAAGCACAGA-3′,5PLATZ2.5′-GACGGGCTGAAGCTCTGTATCACATTGCTC-TCGG-3′;用于3′RACE的引物序列为:3PLATZ1.5′-GCTCTCTTTGCCTGGCTTATC-3′,3′adaptor:5′-CTGATCTAGAGGTACCGGATCCTTTTTTTTTTTTTTTTT-3′。
5′ RACE按照5′-Full RACE Kit说明操作,3′RACE的PCR扩增采用常规的反应程序。PCR产物用12 g/L琼脂糖凝胶电泳,目的片段经切胶回收后克隆并测序,测序结果的拼接由软件DNAMAN V6.0完成。
1.3中国水仙PLATZ1蛋白序列分析
中国水仙PLATZ1蛋白序列的相似性和氨基酸保守结构域预测采用NCBI的BLAST X和BLAST P搜索引擎完成,用ProtParam在线分析蛋白质的理化性质,采用在线工具SSPro(http://scratch.proteomics.ics.uci.edu/)预测蛋白质的二级结构,以NetPhos 2.0 Server(http://www.cbs.dtu.dk/services/NetPhos/)在线软件预测氨基酸的磷酸化位点[12],采用MEGA 6.0软件NJ法进行分子进化树[13]分析。
1.4原核表达载体的构建及表达
根据中国水仙PLATZ1序列的拼接结果设计1对引物(PLATZ1:5′-CGCGGATCCAGGGAGG-AGGTGAGGACAAC-3′;PLATZ2:5′-CGCCTCG-AGCTCCCCCATCATAGAAGCACA-3′),在引物上、下游下划线处分别引入BamHⅠ和XhoⅠ酶切位点,用于扩增中国水仙PLATZ1完整开放阅读框。PCR产物与载体pGEX-4T-3经酶切、回收、连接和转化后,挑取单克隆扩繁,经PCR验证后送上海生工测序。用鉴定为阳性的重组原核表达质粒转化表达宿主菌E.coliBL21(DE3),37 ℃下以200 r/min振荡培养至OD600为0.4~0.6。加入终浓度1 mmol/L IPTG,于37 ℃、150 r/min振荡培养3 h,取1 mL经诱导的菌液于Eppendorf管中,连同诱导前样品于13 000 r/min离心1 min,弃上清,加入50 μL 50 mmol/L的Tris-HCl缓冲液(pH8.0)悬浮沉淀;加等体积2×凝胶上样缓冲液混匀,100 ℃水浴变性处理10 min;蛋白纯化参照Clontech的glutathione resin use manual操作进行;将诱导前、诱导菌样和纯化蛋白各取30 μL经SDS-PAGE检测,考马斯亮蓝R-250染色检测蛋白的表达情况。
1.5NtPLATZ1基因的表达特性分析
以中国水仙不同生长时期的叶片 cDNA为模板,actin为内参[14],PCR检测NtPLATZ1的表达特性。NtPLATZ1引物为PLATZ1和PLATZ2,产物长度747 bp,PCR扩增条件为:94 ℃预变性5 min;94 ℃变性30 s,60 ℃退火30 s,72 ℃延伸50 s,20个循环;actin的PCR扩增程序为:94 ℃预变性5 min;94 ℃变性30 s,50 ℃退火30 s,72 ℃延伸40 s,25个循环。PCR扩增均重复3次。
2 结果与分析
2.1中国水仙PLATZ1基因的5′RACE和3′RACE
以随机引物反转录的水仙叶片cDNA为模板,用引物3PLATZ1和3′adaptor进行PCR;参照5′-Full RACE Kit的方法合成cDNA第1链;再用引物5PLATZ1、5PLATZ2和试剂盒提供的5′RACE Outer Primer、5′RACE Inner Primer进行两轮PCR;3′RACE在850 bp左右有一条亮带(图1-A),5′RACE在630 bp左右有一条亮带(图1-B)。
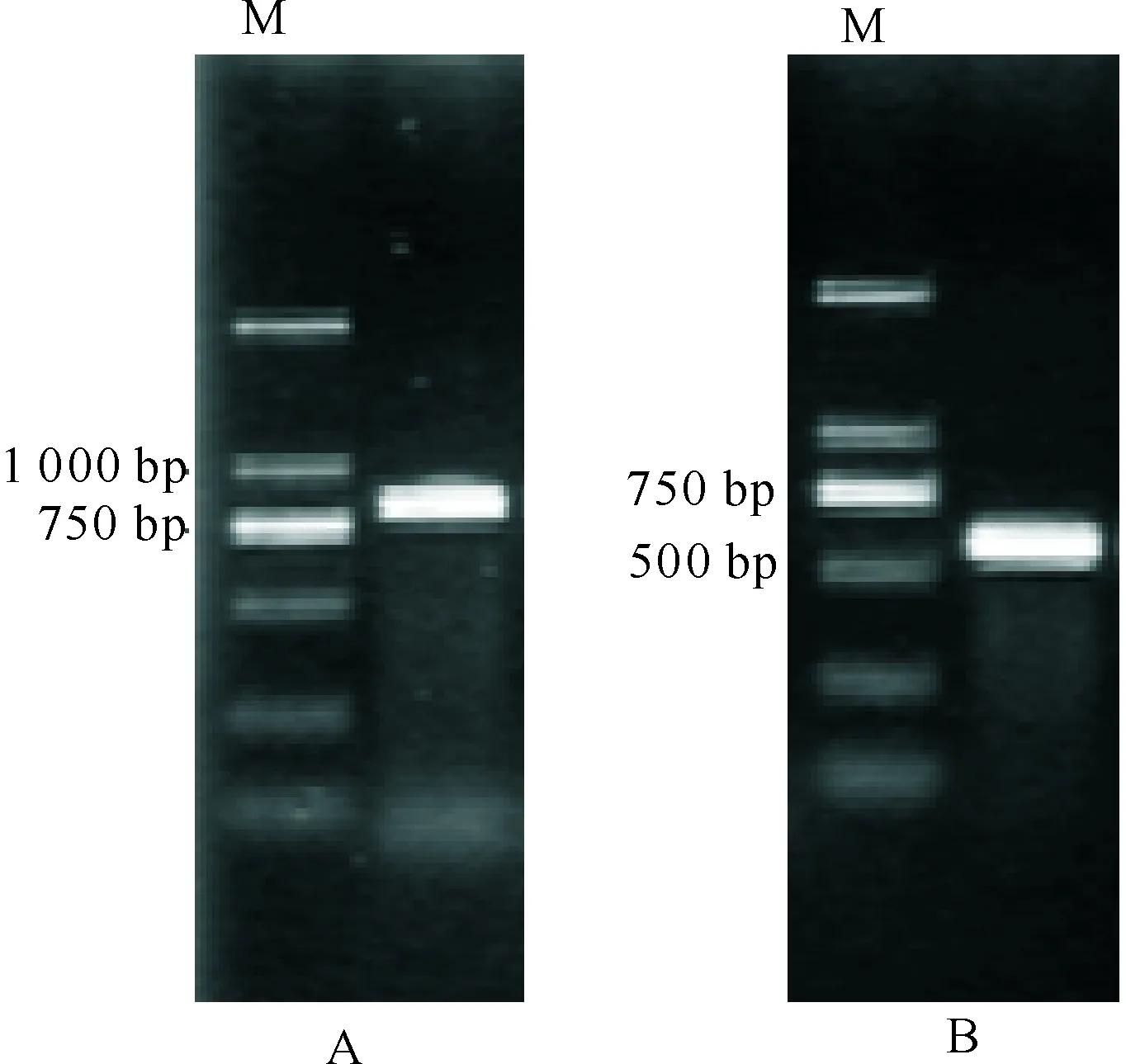
图 1 中国水仙PLATZ1基因的 PCR电泳结果M.分子质量标记(100~2 000 bp);A.3′RACE;B.5′RACEFig.1 Electrophoresis products for PLATZ1 fromNarcissus tazetta var.chinensis by PCRM.DNA Marker (100-2 000 bp);A.3′RACE;B.5′RACE
用DNAMAN V6.0对NtPLATZ1基因3′RACE和5′RACE的测序结果进行序列拼接,并进行开放阅读框分析,结果(图2)表明,所获得的cDNA全长为1 024 bp,包含1个642 bp的完整开放阅读框,编码213个氨基酸,5′末端非翻译区48 bp,3′末端非翻译区334 bp。

图 2中国水仙PLATZ1基因及其推导的氨基酸序列
Fig.2Complete cDNA sequence ofPLATZ1 fromNarcissustazettavar.chinensis
2.2中国水仙PLATZ1基因开放阅读框的克隆
以随机引物反转录的中国水仙PLATZ1基因cDNA为模板,用引物PLATZ1和PLATZ2进行PCR扩增,产物经测序后用DNAMAN V6.0比对,结果与拼接得到的中国水仙PLATZ1开放阅读框完全一致,获得了带BamHⅠ和XhoⅠ酶切位点的中国水仙PLATZ1基因。
2.3中国水仙PLATZ1基因序列的同源性和系统进化分析
使用BLAST X对中国水仙PLATZ1基因推导的氨基酸序列进行同源性检索,结果表明,其所编码的氨基酸序列与大豆(Glycinemax,AII19314.1)、可可(Theobromacacao,XP_007039480.1)、陆地棉(Gossypiumhirsutum,AFH57272.1)的PLATZ氨基酸序列的相似度分别为81%,75%和75%;通过NCBI的BLAST P在线搜索蛋白保守区,相匹配的有1个(图3),为PLATZ superfamily,E值为1.77×10-45。因此,推断该基因为中国水仙的PLATZ基因,命名为NtPLATZ1。

图 3 中国水仙PLATZ1蛋白的保守结构域Fig.3 Putative conserve domains for PLATZ1 protein from Narcissus tazetta var.chinensis
采用MEGA 6.0软件将NtPLATZ1推导的氨基酸序列与其他12个具有PLATZ结构域的物种进行NJ聚类分析,结果(图4)表明,单子叶植物和双子叶植物的PLATZ分别处于两个独立进化的分支中,单子叶植物又分为两类,其中禾本科的玉米与日本稻聚为一类,中国水仙单独聚为一类。

图 4 中国水仙和其他植物PLATZ的分子进化树Fig.4 Molecular phylogenetic tree of NtPLATZ1 and PLATZ from other plants
2.4NtPLATZ1序列的生物信息学分析
ProtParam在线分析结果表明,NtPLATZ1蛋白的分子质量为23.817 ku,理论等电点为9.12,具有19个带负电荷的氨基酸残基(Asp+Glu),28个带正电荷的氨基酸残基(Arg+Lys)。不稳定系数54.11,属于不稳定蛋白;脂肪系数70.94,总平均疏水性为-0.463。
采用SSPro在线工具进行预测发现,在NtPLATZ1蛋白质的二级结构(图5)中,含有146个形成无规则卷曲的氨基酸残基,占68.55%;24个形成α螺旋的氨基酸残基,占11.27%;43个形成β折叠的氨基酸残基,占20.18%。
以NetPhos 2.0 Server在线软件预测NtPLATZ1氨基酸的磷酸化位点,结果表明该蛋白中有19个丝氨酸(serine,S)残基参与磷酸化,酪氨酸(Tyrosine,Y)和苏氨酸(Threonine,T)各有2个参与磷酸化。

图 5 NtPLATZ1蛋白的二级结构预测 大写字母为氨基酸序列;小写字母为二级结构组成:c.无规则卷曲,h.α螺旋,e.β折叠Fig.5 Predicted secondary structure of NtPLATZ1 Capital letters show amino acid sequence;small letters show structure of amino acid chain,c.Random coil,h.α-helix,e.β-extended
2.5NtPLATZ1蛋白的原核表达
将重组原核表达质粒pGEX-NtPLATZ1转化入大肠杆菌(E.coli)BL21(DE3),用1 mmol/L IPTG诱导3 h后,经SDS-PAGE电泳及考马斯亮蓝染色发现,NtPLATZ1蛋白的分子质量为23.817 ku,融合蛋白分子质量为49.817 ku。试验结果(图6)表明,与携带质粒pGEX-4T-3的菌体相比,携带重组质粒pGEX-NtPLATZ1的E.coil菌株中出现1条约49.817 ku的条带,蛋白经纯化后同样获得1条约49.817 ku的条带,说明NtPLATZ1能在大肠杆菌中有效表达。
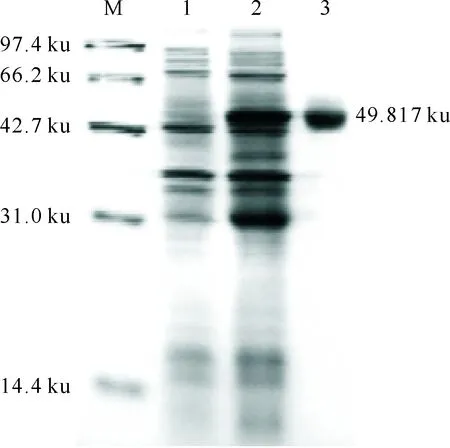
图 6 NtPLATZ1蛋白在大肠肝菌中表达的 SDS-PAGE电泳结果M. 分子质量标记;1.诱导前;2.诱导后;3.纯化Fig.6 SDS-PAGE of NtPLATZ1 expressed in Esherichia coliM.Protein Marker;1.Before induction; 2.After induction;3.Purification
2.6NtPLATZ1基因的表达特性
用半定量RT-PCR法,分析经多效唑处理后,中国水仙不同生长时期叶片中NtPLATZ1基因的表达情况,结果(图7)显示,NtPLATZ1在中国水仙抽葶期和始花期叶片中的表达量较喷雾清水(对照)高,长叶期与盛花期表达量基本一致;从整个生长期来看,喷雾多效唑后,NtPLATZ1的表达量先上升后下降,抽葶期的表达量达到最高,后开始下降;喷雾清水后NtPLATZ1的表达量先下降后又有所回升,抽葶期和始花期的表达量低于长叶期和盛花期。actin基因在各个时期的表达量基本一致。

图 7 经多效唑处理后中国水仙不同生长时期 叶片中NtPLATZ1基因的表达分析 1~4.喷雾多效唑,5~8.喷雾清水;1,5.长叶期; 2,6.抽葶期;3,7.始花期;4,8.盛花期Fig.7 Expression analysis of NtPLATZ1 gene at different growth stages in response to paclobutrazol 1-4.Sprayed with paclobutrazol,5-8.Sprayed with water; 1,5.Leaf stage;2,6.Scape stage;3,7.Early flowering stage;4,8.Full-bloom stage
3 讨 论
本研究从多效唑处理的中国水仙叶片中克隆到了NtPLATZ1基因,其推导的氨基酸序列具有一个PLATZ superfamily,E值为1.77×10-45。所编码的氨基酸与大豆、可可、陆地棉PLATZ氨基酸序列的相似度分别为81%,75%和75%,推断该基因为中国水仙的PLATZ基因。
光敏色素参与了植物种子的萌发、叶的扩展、光周期和开花的诱导等生理功能,并调控相关基因的表达。豌豆的GTPase基因pra2主要在上胚轴表达,但光照会抑制其表达[15]。PLATZ1能与pra2基因中12 bp的顺式作用元件DE1上游的A/T-rich DNA序列结合,下调pra2基因的表达;PLATZ1基因在豌豆的根尖及顶芽中的表达量较高,而在幼叶、成熟叶及茎中的表达量都很低,推测其可能与组织的细胞分化有关[9]。抽葶期和始花期是中国水仙室内水养叶片徒长最快的阶段,喷雾多效唑后,能克服弱光引起的叶片徒长。半定量RT-PCR分析结果表明,从长叶期到盛花期的中国水仙叶片中都能检测到NtPLATZ1的表达,在抽葶期和始花期,喷雾清水时NtPLATZ1表达量较低,喷雾多效唑则能明显上调NtPLATZ1的表达。因此,PLATZ可能参与了光信号转导过程中基因表达的调控。
PLATZ1与A/T-rich DNA序列的结合是非特异的。豌豆petE基因的增强子含有A/T-rich DNA序列[16],蛋白HMG-1和HMG-I/Y与增强子结合可提升petE基因的表达[17],PLATZ1能与petE基因增强子的A/T rich DNA序列结合,减弱该增强子的功能,下调petE基因的表达[9]。而对于中国水仙NtPLATZ1能调控哪些基因的表达及其机制,均有待进一步研究。
[1]Benjamin L.Gene(Ⅸ) [M].Boston,Toronto,London,Singapore:Jones and Bartlett Publishers,2007:654-656.
[2]黄骥,王建飞,张红生.植物C2H2型锌指蛋白的结构与功能 [J].遗传,2004,26(3):414-418.
Huang J,Wang J F,Zhang H S.Structure and function of plant C2H2 zinc finger protein [J].Hereditas,2004,26(3):414-418.
[3]黄骥,张红生.TFIIIA 型锌指蛋白及在提高植物耐逆性中的作用 [J].遗传,2007,29(8):915-922.
Huang J, Zhang H S.The plant TFIIIA-type zinc finger proteins and their roles in abiotic stress tolerance [J].Hereditas,2007,29(8):915-922.
[4]宋冰,洪洋,王丕武,等.植物C2H2型锌指蛋白的研究进展 [J].基因组学与应用生物学,2010,29(5):1133-1141.
Song B,Hong Y,Wang P W,et al.Advances on plant C2H2-type zinc finger protein [J].Genom Appl Biol,2010,29(5):1133-1141.
[5]Nakagawa H,Jiang C J,Sakakibara H,et al.Over expression of a petunia Zinc-finger gene alters cytokinin metabolism and plant forms [J].Plant J,2005,41(4):512-523.
[6]Dathan N,Zaccaro L,Esposito S,et al.TheArabidopsisSUPERMAN protein is able to specifically bind DNA through its single Cys2-His2zinc-finger motif [J].Nucleic Acids Res,2002,30(22):4945-4951.
[7]陈晨,彭辉,高文瑞,等.鹰嘴豆锌指蛋白基因ZF1的克隆及表达分析 [J].作物学报, 2009,35(12):2180-2186.
Chen C,Peng H,Gao W R,et al.Cloning and expression of zinc finger protein geneZF1 in Chickpea (CicerarietinumL.) [J].Acta Agron Sin,2009,35(12):2180-2186.
[8]郭丽香,高世庆,唐益苗,等.小麦TaCRF2基因的克隆及其在烟草中的初步功能验证 [J].作物学报,2011,37(8):1389-1397.
Guo L X,Gao S Q,Tang Y M,et al.TaCRF2 gene isolation fromTriticumaestivumand primary function validation in tobacco (NicotianatabacumL.) [J].Acta Agron Sin,2011,37(8):1389-1397.
[9]Yukio N,Hirofumi F,Takehito I,et al.A novel class of plant-specific zinc-dependent DNA-binding protein that binds to A/T-rich DNA sequecne [J].Nucleic Acids Res, 2001,29(20):4097-4105.
[10]王伟英,邹晖,戴艺民,等.多效唑喷雾对中国水仙的矮化效应 [J].热带作物学报,2011,32(9):1661-1664.
Wang W Y,Zou H,Dai Y M,et al.The dwarfing effect onNarcissustazettavar.chinensisby spraying with paclobutrazol [J].Chin J Trop Crops,2011,32(9):1661-1664.
[11]林江波,王伟英,邹晖,等.中国水仙叶绿素ab结合蛋白基因Ntcab1的克隆与序列分析 [J].福建农业学报,2013,28(5):463-467.
Lin J B,Wang W Y,Zou H,et al.Cloning and sequence analysis on gene of chlorophyll a/b-binding protein fromNarcissustazettavar.chinensis[J].Fujian J Agric Sci,2013,28(5):463-467.
[12]Blom N,Gammeltoft S,Brunak S.Sequence-and structure-bas-ed prediction of eukaryotic protein phosphorylation sites [J].J Mol Bio,1999,294(5):1351-1362.
[13]Tamura K,Stecher G,Peterson D,et al.MEGA6:Molecular evolutionary genetics analysis Version 6.0 [J].Mol Biol Evol,2013,30:2725-2729.
[14]林江波,王伟英,邹晖,等.中国水仙肌动蛋白基因的克隆与表达分析 [J].福建农业学报,2012,27(9):931-935.
Lin J B,Wang W Y,Zou H,et al.Cloning and expression analysis ofactingene fromNarcissustazettavar.chinensis[J].Fujian J Agric Sci,2012,27(9):931-935.
[15]Nagano Y,Okada Y,Narita H,et al.Location of light-repressible,small GTP-binding protein of the YPT/rab family in the growing zone of etiolated pea stems [J].Proc Natl Acad Sci USA,1995,92:6314-6318.
[16]Sandhu J S,Wdbster C I,Gray J C.A/T-rich sequences act as quantitative enhancers of gene expression in transgenic tobacco and potato plant [J].Plant Mol Biol,1998,37(5):885-896.
[17]Webster C I,Packman L C,Pwee K H,et al.High mobility group proteins HMG-1 and HMG-I/Y bind to a positive regulatory region of the pea plastocyanin gene promoter [J].Plant J,1997,11(4):703-715.
Cloning and expression analysis ofNtPLATZ1 gene fromNarcissustazettavar.chinensis
LIN Jiangbo,WANG Weiying,LI Haiming,WU Shuijin,ZOU Hui,LI Yuesen,DAI Yimin
(SugarcaneResearchInstitute,FujianAcademyofAgriculturalSciences,Zhangzhou,Fujian363005,China)
【Objective】 This study aimed to clone plant AT-rich sequence and zinc-binding protein gene fromNarcissustazettavar.chinensis(NtPLATZ1) for studying its biological function.【Method】 RACE and RT-PCR technique were used to clone NtPLATZ1,the bioinformatics analysis was conducted to analyze its sequence,and semi-quantitative RT-PCR was used to detect its expression.NtPLATZ1 was inserted into pGEX-4T-3,the prokaryotic expression vector was transformed into BL21,and protein expression was induced.【Result】 The full length ofNtPLATZ1 cDNA was 1 024 bp with an 642 bp ORF,which encoded a protein of 213 amino acids with a conservative domain of PLATZ superfamily.NtPLATZ1 shared 81% identity with PLATZ ofGlycinemax(AII19314.1).NtPLATZ1 was predicted an unstable protein.Prokaryotic expression showed that recombinant pGEX-NtPLATZ1 had high expression inE.coliBL21.Semi-quantitative RT-PCR revealed thatNtPLATZ1 expression was enhanced at scape stage and decreased hereafter after it was sprayed with paclobutrazol,while it was decreased at scape and early flowering stages without paclobutrazol. 【Conclusion】NtPLATZ1 was clone and its expression was enhanced significantly by paclobutrazol.
Narcissustazettavar.chinensis;PLATZ;gene expression;zinc finger protein
时间:2016-09-0709:03DOI:10.13207/j.cnki.jnwafu.2016.10.023
2015-03-27
福建省自然科学基金项目(2014J01098)
林江波(1976-),男,福建永春人,副研究员,硕士,主要从事观赏植物生物技术研究。E-mail:linjiangbo1976@qq.com
戴艺民(1969-),男,福建长泰人,研究员,博士,主要从事观赏植物生物技术研究。E-mail:dymttcn@163.com
Q78;S682.2+1
A
1671-9387(2016)10-0165-06
网络出版地址:http://www.cnki.net/kcms/detail/61.1390.S.20160907.0903.046.html
