酶制剂在家禽上的应用研究进展
2016-11-07喻小琼任文刘宏汪仕奎
喻小琼,任文,刘宏,汪仕奎
帝斯曼(中国)动物营养研发有限公司,河北霸州165799
综述
酶制剂在家禽上的应用研究进展
喻小琼,任文,刘宏,汪仕奎
帝斯曼(中国)动物营养研发有限公司,河北霸州165799
酶制剂作为一种新型高效饲料添加剂,添加到家禽饲料中可消除原料中的抗营养因子,提高日粮组分的消化率,改善动物生产性能及增加经济和生态效益。但受限于酶本身及受试对象等方面的影响,其产生的效果不尽一致。这些问题的存在,影响了科研人员对酶制剂的深度开发利用,制约了酶制剂在市场上的推广。本文旨在通过引用近年来酶制剂在家禽试验中的科研数据,对酶制剂在家禽生产中的应用效果进行探讨,以期为酶制剂科学研发与合理利用提供参考依据。
酶制剂;效应差异;家禽
随着畜牧业的迅猛发展,饲料原料资源短缺问题日益突出,如何充分提高饲料原料的利用率是亟待解决的难题。近年来,随着基因工程与酶工程等新型学科的交叉应用,世界范围内酶制剂的生产与研制达到了前所未有的水平。复合酶制剂的合理添加能够不同程度地提高家禽生长代谢性能,改善动物福利,提高产品品质,从而提高畜牧业经济效益。但在实际应用中,酶的应用效果受到酶本身(来源、剂量)及其作用对象(动物、底物、试验环境)等诸多因素的影响,从而限制了酶制剂功效的发挥。为此,本文对相关酶在家禽日粮中的应用状况进行综述,分析了酶制剂应用效果存在的差异及影响作用效果的因素,为深入研究酶制剂作用机理和合理应用饲用酶提供参考。
1 禽饲料酶制剂的生物学特性
1.1禽饲料酶制剂的酶学特性自然界中酶制剂来源不同,其生物表现形式如折叠方式、酶底物特异性、水解活性等理化特性等差异较大(岳晓禹等,2007)。以家禽饲料中最常用的木聚糖酶为例,此酶主要由细菌和真菌产生,但达到稳定性的最佳pH和最适反应温度有很大差别。一般来说,真菌产生酸性木聚糖酶,最佳反应pH范围为4.0~6.0,适宜温度在50℃左右;细菌和放线菌产生中性或碱性木聚糖酶,其最佳反应pH为6.0~8.0,最适反应温度为50~60℃,其耐热性要优于真菌(马文静等,2008)。
酶制剂来源不同,其催化特性有很大差异,尤其是最佳酶活的温度、pH及耐热性等不同(表1),因而在生产中应用的效果不同。
1.2影响酶活性的关键因素
1.2.1制粒温度对饲料酶活性的影响调质制粒、挤压膨化等加工过程会导致饲料热敏性酶制剂的损失。孙相俞等(2015)报道,通过比较不同的调制设备及不同的操作参数(调质时间、热能和制粒温度)等对酶回收率的影响,发现对酶稳定性起关键影响因素为制粒温度。杨海锋等(2014)研究表明,随着调质温度的升高酶活性显著下降,在60℃调质温度下,耐高温型β-葡聚糖酶活和纤维素酶活保存率分别为91.3%和90.4%;在85℃调质温度下,其酶活保存率下降至36.7%和58.2%。牛化欣等(2014)研究后熟化条件对饲用酶活性的损伤的影响表明,在温度85~95℃和0~40 min后熟化过程中,随着温度的升高和后熟化时间的延长,饲用酶的活性下降,在85、90、95℃下后熟化40 min时,蛋白酶活力分别下降了37.7%、51.0%、86.5%,淀粉酶活力分别下降了17.6%、26.0%、33.0%。

表1 禽用酶制剂酶学性质
1.2.2金属离子对饲料酶活性的影响在实际应用中,添加到饲料中的酶制剂活性往往低于包装袋规格,部分原因是由于饲料中金属离子结合酶功能基团引发酶的变性。赵玉蓉等(2004)研究表明,Na+、K+、Mg2+、Zn2+、Mn2+、Fe3+、Ca2+对纤维素酶活性起激活作用,Cu2+起轻微的抑制作用,其激活率随Mn2+离子浓度的增加而提高。而Na+、Ca2+、K+对木聚糖酶活性有激活作用,而Cu2+、Mg2+、Mn2+、Zn2+、Fe3+均对木聚糖酶起抑制作用,其抑制率随Cu2+离子浓度的增加而提高,可能是Cu2+离子结合了木聚糖酶必需基团的活性中心造成的。张红艳等(2009)在研究金属离子对饲用复合酶活性的影响表明,Mg2+对蛋白酶有一定的激活作用,Mn2+的激活作用最大,Ca2+、Zn2+、Cu2+、Fe2+对蛋白酶活有不同程度的抑制作用,其大小顺序依次为Cu2+>Fe2+>Zn2+>Ca2+。李忠玲等(2014)研究表明金属离子对于淀粉酶激活作用不明显,抑制作用大小顺序依次为Fe2+>Cu2+>Mn2+>Zn2+>Co2+。
2 家禽消化酶分泌特征及酶添加的时间规律
2.1家禽内源消化酶分泌及其活性变化规律禽类从胚胎期靠卵黄囊营养供给的低级方式到出雏后消化吸收外源养分的高级形式转变,其消化器官发育及消化酶分泌变化起着关键的作用。安永义等(1999)在研究0~3周龄Avian肉仔鸡公雏胰腺和肠道食糜消化酶发育规律发现,其消化酶活性(U/kg体重)随着出雏后体重的增大而增多,10日龄时各种消化酶活性达到最大。在胰腺中(酶活用U/g胰重表示),各种酶(除脂肪酶)在孵出几天后呈现出先下降后上升的趋势,淀粉酶和胰蛋白酶活力在17日龄时达到峰值;糜蛋白酶活力在10日龄前趋于稳定,随后逐步升高,至21日龄达到峰值;而脂肪酶总体分泌呈上升态势,在14日龄时稍微下调,于21日龄达到峰值,这说明胰腺合成酶在逐步增加,随着日龄的增长,胰腺的分泌对采食外源性饲料逐步适应。在肠道内容物中(酶活用U/g食糜表示),肠道中除了脂肪酶活性呈先上升后下降趋势,淀粉酶、胰蛋白酶和糜蛋白酶活性均呈先上升后平稳的趋势,在14~21日龄达最大值。胰腺分泌的酶,只有进入肠道才具有活性并发挥作用,因而肠道酶的变化更好地说明消化的发展过程。Sklan和Noy(2003)在研究雏鸡出雏19日龄消化酶分泌规律时也发现,肠道消化酶量随日龄的增大逐渐升高,且淀粉酶在19日龄时分泌最多,其次为胰蛋白酶,最少的为脂肪酶。
2.2家禽酶制剂添加的日龄规律Bedford和Classen(1992)报道幼禽消化器官发育不完善,消化酶分泌的不足,影响动物的生长,外源酶在机体酶分泌低谷期的添加能减缓或降低生长抑制。Olukosi等(2007)研究表明,雏鸡日粮添加酶,效果在4周龄以前比较明显,随着日龄的增大,肠道微生物区系趋于稳定,分泌酶活性逐渐增强,致使肠道对日粮非淀粉多糖(NSP)酶的添加敏感降低,对酶制剂的依赖性大大减少。
Gracia等(2009)在玉米-豆粕型日粮中添加NSP复合酶(木聚糖酶、蛋白酶和α-淀粉酶)的研究表明,酶制剂提高肉鸡出生后1~4 d的体重和饲料转化效率,对5~21 d的生长性能无显著影响。Santos等(2013)在肉鸡黑麦类日粮中分阶段(1~7 d、1~14 d、1~21 d、1~28 d)添加酶制剂,观察对其生长性能的影响,发现21 d与28 d加酶组与对照组比,肉鸡生长性能、胃肠道消化酶活及器官大小均未产生显著差异,表明此酶制剂在黑麦类饲粮的添加促进前3周龄肉鸡的生长,但对3周龄后肉鸡的促进效果甚微。
马卿山等(2013)在研究玉米-豆粕型饲粮添加脂肪酶对肉鸡生长性能及养分利用率的影响发现,肉鸡的改善作用后期要大于前期,可能是由于前期肉鸡摄食量较低,脂肪酶的作用有限。此外,张铁鹰等(2005)研究发现,肉鸡1~49日龄的空肠食糜脂肪酶活性(U/g鲜重)呈三次曲线关系,在14日龄达到峰值后,到42日龄一直降低,并且腺胃和肌胃pH从7~42日龄一直升高。空肠是脂肪吸收的主要场所,因此建议在肉鸡中后期日粮中添加酶制剂可能更有利于体重的增加,并且腺胃和肌胃pH的升高有利于外源脂肪酶活性在消化道内的稳定。
2.3家禽自身的生理限制对消化酶发挥作用的影响尽管家禽体内适宜的温度能满足酶发挥活性,但由于消化道内pH值波动较大,无疑对酶的稳定性及活力释放造成一定的限制。另外,家禽消化道短,食糜停留时间短也限制了酶的充分释放。由表2可知,外源酶在家禽体内需要经历弱酸性的嗉囔环境,到达酸性的肌胃,之后进入由弱酸性逐渐变为中性的肠道中,而大部分外源酶发挥活力的最适pH为4~6,因而限制了酶活力的充分释放。但由于来源的差异,使得有些酶能够耐受前胃的降解作用而进入肠道发挥更大的效果。Svihus(2011a)报道饲料停留在整个消化道的时间大概为3~4 h,食糜在前段消化道的时间为60~90 min,因而提供了不足的酶反应时间。且自由采食及粉料的饲喂方式不足以促进嗉囔和肌胃的正常发育,而未发育成熟的肌胃仅仅充当了转运而不是研磨的作用,这样极大减低了饲料的停留时间,降低了酶活的释放。Svihus(2011b)研究表明,饲喂粗饲料及整个谷物能有效刺激肌胃的生长,促使肌胃发挥更好的研磨功能,增加了食糜的逆向蠕动,促进肌胃容量的扩张,因而增加了停留时间,促进酶活力的发挥。
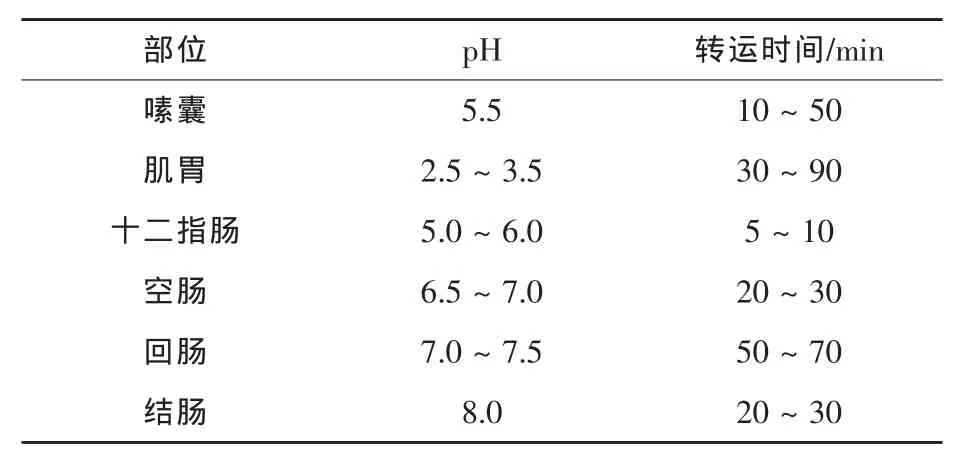
表2 肉鸡消化道不同部位食糜的平均消化转运时间及pH
3 饲料原料及配方营养水平对酶效果的影响
3.1饲料原科的多样性对酶效果的影响饲料原科作为酶制剂发生催化反应的底物,不同原料不仅其NSP含量不同,且NSP结构和物理化学特性也相差较大。Knudsen(2014)在家禽常用饲料原料纤维及非淀粉多糖含量的报道中提到,对于大多数麦类及谷物原料,黑麦、黑小麦、小麦、玉米、高粱中富含阿拉伯木聚糖,大麦和燕麦主要为β-葡聚糖;且大部分原料(除了玉米和高粱)的NSP是可溶性的。对于谷物副产品、麸皮及谷壳类,主要NSP为纤维素、木质素及不可溶的阿拉伯木聚糖。作为家禽饲料的主要抗营养NSP,β-葡聚糖与阿拉伯木聚糖均是高分子聚合物,易产生黏性,但阿拉伯木聚糖更耐受降解,导致富含阿拉伯木聚糖的原料更容易产生黏性问题。
近年来,国内外研究人员对不同饲料原料中NSP含量与组成做出大量研究(表3)。Jaworski等(2015)检测了12种饲料原料(玉米、小麦、高粱及其副产物)的NSP组成,证实谷物产品相对于其副产品中NSP含量低,淀粉含量高,更容易被家禽消化。同时发现原料干物质的消化率负相关于NSP水平(P=0.97),表明NSP的组成对谷物及副产品的能值释放有一定影响。

表3 部分常见饲料原料中NSP的含量和组成

表4 玉米-豆粕型饲粮中添加复合酶对肉鸡生长性能的影响
3.2同种谷物地域差异对酶效果的影响不同谷物的NSP含量、结构和物理化学特性差别很大,即使是同一谷物也因品种、气候、收获期及产地条件的不同差异很大(Mccracken等,2002)。同样是小麦,越是低质量的小麦,酶的改善效果越明显。Annison(1991)对澳大利亚13种小麦品种的调查表明,鸡对约25%的小麦品种的表观代谢能值(AME)低于13 MJ/kg干物质。Annison(1992)进一步研究表明,这主要是由于其中的水溶性阿拉伯木聚糖含量不同引起的,且随着NSP含量的增加,AME呈线性降低。而酶的添加可以一定程度上消除来自不同批次原料的差异。
3.3饲料原料质量对酶效果的影响不同批次的小麦,其初始AME在2200~3820 kcal/kg的能值区间变异(Wiseman,2000)。木聚糖酶的添加效果因小麦初始AME的不同而不同。Ravindran和Son(2011)统计了大量不同能值范围的小麦原料与木聚糖酶作用关系,发现对于质量较差的小麦(初始AME<2870 kcal/kg),酶添加对原料的AME有11%的提高;对于一般质量的小麦(初始AME为2870~3350 kcal/kg),酶作用能提高近5%;而对于优质的小麦(初始AME>3350 kcal/kg),仅有1%的提高,说明小麦质量越差,其对添加酶的反应越显著,酶的效果发挥空间就越大。
3.4日粮NSP水平应随肉鸡生长阶段的变化而变化肉鸡随着饲养阶段的变化,其饲粮配方中NSP含量也会随之变化。Ward(2014)研究报道,在含有3%DDGS典型的肉鸡配方中,随着生长阶段推进,能量水平增高,蛋白质含量降低,饲料配方中玉米含量增加,豆粕含量降低,日粮NSP水平随着生长阶段的变化而相应改变,相对生长前期,后期配方中的阿拉伯木聚糖增加了22%,低聚糖和果胶降低了22%~30%,纤维素及β-葡聚糖的比例相差不大。理论上,同一配比的酶组合并不能适用于肉鸡生长全期。
3.5日粮配方组成对酶效果的影响酶制剂效果的发挥主要受到原料类型及能量水平等方面影响。一般在配方日粮中使用更多的高黏度谷类杂粕性原料,或者营养水平下降至常规饲养标准的理想营养水平之下时,更有利于外源酶制剂发挥对饲料营养利用率提高的作用。
3.5.1不同日粮类型在不同的日粮组合类型中,玉米-豆粕型日粮因含有少量的抗营养物质,因而不会引发严重的消化道黏度问题(Mccracken等,2002)。由表4可知,多数试验证实酶制剂对玉米-豆粕型日粮无明显改善效果,仅有少数研究得出NSP复合酶在一定范围内改善了体增重、饲料转化效率(FCR)等生长性能指标,且发现酶在玉米-豆粕型日粮的添加相对负对照对FCR的平均改善率为2.3%,对平均体增重改善率2.0%。相反,多数在麦类杂粕型日粮的研究上表明,肉鸡日粮中添加NSP复合酶制剂能显著改善营养成分的利用率,而且饲料配方中NSP含量越高,NSP复合酶酶解效果越好,其对肉鸡生长性能、肠道发育等指标的改善程度总结见表5。
总体来看,玉米-豆粕型日粮中通过添加非特异性酶(木聚糖酶、蛋白酶、淀粉酶)对动物生产性能改善的程度比较低,可能存在如下自身局限(Slominski,2011):(1)玉米豆粕原料中水溶性NSP不会产生黏性问题;(2)淀粉的消化率高;(3)原料中大量存在的寡糖充当能量来源;(4)NSP物质,如阿拉伯木聚糖、葡聚糖、纤维素、甘露糖及果胶作为能量来源,促进了肠道健康;(5)豆粕中的糖蛋白质不能被有效利用。
3.5.2日粮能量水平日粮中的NSP在动物消化道内的酶的作用下分解成短链脂肪酸如乙酸、丙酸等,释放能量,可被肠道吸收利用。Zhou等(2009)研究结果表明,在低能量水平日粮中添加复合酶对日粮代谢能提高幅度越大,使用效果更好。Zhu等(2014)研究表明,在日粮能量降低的情况下添加NSP复合酶提高了小肠的消化酶活力及吸收能力。由于酶对底物的专一性,因此不同的日粮类型需使用相应酶制剂。曾容愚等(2006)研究表明,NSP酶制剂可显著提高小麦组肉鸡生长前期日增重(P<0.05),但对中期、后期无显著影响;而低能小麦组的变化趋势正好与此相反,前期日增重变化不显著,中后期日增重显著提高(P<0.05)。这种阶段性的差异在一定程度上反映了肉鸡饲喂阶段、营养水平等因素对酶制剂应用效果的影响。总结近几年的研究结果发现,酶制剂的添加能提高日粮平均代谢能值0.42 MJ/kg,因而在设计加酶配方时,对玉米-豆粕型可在原配方基础上降低2%~5%能量,对于小麦-豆粕型、大麦-豆粕型日粮,能量调整幅度为6%~10%。

表5 肉鸡麦类杂粕型饲粮中添加复合酶对肉鸡生长性能的影响
4 饲用酶制剂应用研究展望
饲用酶制剂不但可以促进动物对营养物质的消化吸收,提高饲料利用率,改善动物的生长性能,有效降低疾病的发生率;并且其无毒副作用,几乎无残留,大大减少了环境污染。然而在进一步产品的研发中,还应该注重以下几个方面:
一是复合酶产品的组合及识别。复合酶能产生比单酶更大的生产效益,但酶复配的功效建立在单酶的基础上,阐明单一酶是如何水解相应的底物的机理是很有必要的。例如,木聚糖酶产品在含有有效降解可溶性和不可溶性的木聚糖的前提下才能发挥最大的效益,而那些专门亲和不可溶性木聚糖促使可溶性水解产物增多,会导致食糜的黏性增加(Choct等,2004)。Tahir等(2008)评估半纤维素酶、纤维素酶及果胶酶单独或者混合物产品的水解能力,发现半纤维素酶是营养物质水解程度限速步骤,因此,仅含纤维素酶及果胶酶的NSP产品的降解作用甚微。除此之外,阐明底物与酶结合的精确定位能促进辅酶的作用发挥并且促进底物的降解(Adeola和Cowieson,2011)。
二是基因表达与潜在的生理机制。从生理角度出发,利用基因表达规律分析复合酶的添加对动物生长性能的影响是非常重要的。Jozefiak等(2010)通过在家禽日粮中添加碳水化合物酶及植酸酶研究其对生长相关基因表达的影响,结果表明,酶的添加降低胰岛素样生长因子基因的表达(P=0.083),酶的添加通过影响与代谢相关基因的表达影响了生长性能。总之,通过分子调控机理可阐释酶对生长代谢的生理作用,可为研究酶是如何发挥作用的提供一种新视角。
三是非常规饲料酶制剂的开发。由于常规玉米、豆粕原料成本上涨,市场上开始积极寻求成本低廉且营养丰富的非常规饲料原料,如大麦、高粱及DDGS,这些原料由于价格低廉、营养丰富,有明显的市场前景。但它们的使用受到植酸、NSP、生长抑制因子、单宁等的影响,从而降低原料的利用价值。总体上,当前的酶在家禽非常规原料上发挥的效用是有限的,并且有针对性的酶的开发还比较缺乏。随着对非常规饲料原料中抗营养因子的功能特性、抗营养机理和对策的深入认识,这个领域将会有更多的研究成果。
[1]安永义,周毓平,呙于明,等.0~3周肉仔鸡消化道酶发育规律的研究[J].动物营养学报,1999,1:17~24.
[2]国春艳,刁其玉,乔宇,等.酸性木聚糖酶产生菌株的筛选和酶学性质分析[J].中国农业科学,2010,7:1524~1530.
[3]李用芳,李学梅,张建新,等.木聚糖酶产生菌的选育及发酵条件研究[J].生物技术,2004,1:25~27.
[4]李忠玲,徐升运,李文孝,等.复合酶制剂羊饲料添加剂中纤维素酶和淀粉酶影响因素的研究[J].中国酿造,2014,11:85~89.
[5]马卿山,丁雪梅,白世平,等.饲粮添加脂肪酶对肉鸡生长性能及养分利用率的影响[J].动物营养学报,2013,10:2447~2458.
[6]马文静,张美云,房桂干.木聚糖酶的应用研究进展[J].江苏造纸,2008,1:20~24.
[7]牛化欣,常杰,后卫.水产饲料后熟化工艺对饲用酶活力的影响[J].饲料工业,2014,20:40~43.
[8]孙春阳,何科林,萨仁娜,等.非淀粉多糖复合酶对肉仔鸡生产性能和排泄物中含氮物含量的影响[J].动物营养学报,2014,5:1370~1378.
[9]孙相俞,万建美,Jér Me Lamoine.饲料加工技术有助于酶活性回收率[J].国外畜牧学(猪与禽),2015,2:64~65.
[10]王绍花,宗工理,刘飞,等.不同来源木聚糖酶酶学性质研究[J].食品与药品,2013,6:397~399.
[11]闻雅男,赵娜,滕雪莹,等.黑曲霉植酸酶高产菌株产酶条件的筛选[J].辽宁农业科学,2011,3:16~19.
[12]杨海锋,刘泽辉,宋卫国,等.制粒工艺对饲料中-葡聚糖酶及纤维素酶活性影响的研究[J].粮食与饲料工业,2014,3:35~37.
[13]尹兆正.高麸加酶替代玉米饲粮对肉仔鸡生长性能的影响[J].浙江农业学报,2005,4(17):191~195.
[14]岳晓禹,贺小营,牛天贵,等.木聚糖酶的研究进展[J].酿酒科技,2007,4:113~115.
[15]曾容愚,张莉莉,王恬.不同能量水平小麦日粮添加非淀粉多糖酶制剂对肉鸡生产性能的影响[J].动物营养学报,2006,4:278~282.
[16]张红艳,马齐,岳淑宁,等.常见金属离子对饲用复合酶活性的影响[J].黑龙江畜牧兽医,2009,21:62~63.
[17]张铁鹰,汪儆,李永清.0~49日龄肉仔鸡消化参数的变化规律研究[J].中国畜牧兽医,2005,1:6~10.
[18]赵必迁,张克英,丁雪梅.非淀粉多糖复合酶制剂对肉鸡生产性能及养分表观利用率的影响[J].家禽科学,2012,2:3~8.
[19]赵玉蓉,金宏,陈清华,等.金属离子对饲料酶活性的影响[J].饲料研究,2004,8:1~3.
[20]邹佳伶.酶制剂种类与能量水平对肉鸡生产性能和消化生理的影响研究:[硕士学位论文][D].四川农业大学,2012.
[21]Adeola O,Cowieson A J.BOARD-INVITED REVIEW:opportunities and challenges in using exogenous enzymes toimprovenonruminant animal production[J].Journal of Animal Science,2011,89(10):3189~3218.
[22]Amerah A M,van de Belt K,van Der Klis J D.Effect of different levels of rapeseedmealandsunflowermealandenzymecombinationonthe performance,digesta viscosity and carcass traits of broiler chickens fed wheatbased diets[J].Animal,2015,9(7):1131~1137.
[23]AnnisonGCM.Anti-nutritiveactivitiesofcerealnon-starch polysaccharides in broiler diets and strategies minimizing their effects[J]. World's Poultry Science Journal,1991,3(47):232~242.
[24]Annison G.Commercial enzyme supplementation of wheatbased diets raises ilealglycanase activities and improves apparent metabolisable energy,starch and pentosandigestibilities in broiler chickens[J].Animal Feed Science and Technology,1992,2(38):105~121.
[25]Bedford M R,Classen H L.Reduction of intestinal viscosity through manipulation of dietary rye and pentosanase concentration is effected through changes in the carbohydrate composition of the intestinal aqueous phase and results in improved growth rate and food conversion efficiency of broiler chicks[J].Journal of Nutrition,1992,122(3):560~569.
[26]Choct M.Feed non-starch polysaccharides:Chemical structures and nutritional significance[J].Feed milling international,1997,191:13~26.
[27]Choct M,Kocher A,Waters D L,et al.A comparison of three xylanases on the nutritive value of two wheats for broilerchickens[J].Br J Nutr,2004,92(1):53~61.
[28]Cowieson A J,Bedford M R,Ravindran V.Interactions between xylanase and glucanase in maize-soy-based diets for broilers[J].Br Poult Sci,2010,51(2):246~257.
[29]Cowieson A J,Ravindran V.Effect of exogenous enzymes in maize-based diets varying in nutrient density for young broilers:growth performance and digestibility of energy,minerals and amino acids[J].Br Poult Sci,2008,49(1):37~44.
[30]DotsenkoGS,SemenovaMV,SinitsynaOA,etal.Cloning,purification,and characterization of galactomannan-degrading enzymes from Myceliophthorathermophila[J].Biochemistry(Mosc),2012,77(11):1303~1311.
[31]Golovan S,Wang G,Zhang J,et al.Characterization and overproduction of the Escherichia coli appA encoded bifunctional enzyme that exhibits both phytase and acid phosphatase activities[J].Canadian Journal of Microbiology,2000,46(1):59~71.
[32]GraciaMI,LázaroR,LatorreMA,etal.Influenceofenzyme supplementation of diets and cooking–flaking of maize on digestive traits and growth performance of broilers from 1 to 21 days of age[J].Animal Feed Science and Technology,2009,150(3~4):303~315.
[33]Greiner R,Da S L,Couri S.Purification and characterisation of an extracellular phytase from Aspergillus niger11T53A9[J].Brazilian Journal of Microbiology,2009,40(4):795~807.
[34]Jaworski N W,Laerke H N,Bach K K,et al.Carbohydrate composition and in vitro digestibility of dry matter and nonstarch polysaccharides in corn,sorghum,and wheat and coproducts from these grains[J].Journal of Animal Science,2015,93(3):1103~1113.
[35]Jozefiak D,Ptak A,Kaczmarek S,et al.Multi-carbohydrase and phytase supplementation improves growth performance and liver insulin receptor sensitivity in broiler chickens fed diets containing full-fat rapeseed[J].Poult Sci,2010,89(9):1939~1946.
[36]Kaper T,van Heusden H H,van Loo B,et al.Substrate specificity engineering of beta-mannosidase and beta-glucosidase from Pyrococcus by exchangeofuniqueactivesiteresidues[J].Biochemistry,2002,41(12):4147~4155.
[37]Knudsen K E.Fiber and nonstarch polysaccharide content and variation in common crops used in broiler diets[J].Poult Sci,2014,93(9):2380~2393.
[38]Mccracken K J,Preston C M,Butler C.Effects of wheat variety and specific weight on dietary apparent metabolisable energy concentration and performance of broiler chicks[J].Br Poult Sci,2002,43(2):253~260.
[39]Momtazan.R M H Z M.A note on the effects of a combination of an enzyme complex and probiotic in the diet on performance of broiler chickens[J].Irish Journal of Agricultural and Food Research,2011,50:249~254.
[40]Olukosi O A,Cowieson A J,Adeola O.Age-related influence of a cocktail of xylanase,amylase,and protease or phytase individually or in combination in broilers[J].Poult Sci,2007,86(1):77~86.
[41]Ouhida L P J F.Enzymes(13-glucanase and arabinoxylanase)and/ orsepiolite supplementation and the nutritive value of maize-barley-wheat based diets for broiler chickens[J].British Poultry Science,2000,41:617~624.
[42]Puhl A A,Greiner R,Selinger L B.Stereospecificity of myo-inositol hexakisphosphate hydrolysis by a protein tyrosine phosphatase-like inositol polyphosphatasefromMegasphaeraelsdenii[J].ApplMicrobiolBiotechnol. 2009,82(1):95~103.
[43]Ravindran V,Son J H.Feed enzyme technology:present status and future developments[J].Recent Pat Food Nutr Agric,2011,3(2):102~109.
[44]Santos C I,Ribeiro T,Ponte P,et al.The effects of restricting enzyme supplementation in rye-based diets for broilers[J].Animal Feed Science and Technology,2013,186(3):214~217.
[45]Shi H,Huang Y,Zhang Y,et al.High-level expression of a novel thermostable and mannose-tolerant beta-mannosidase from Thermotogathermarum DSM 5069inEscherichiacoli[J].BMCBiotechnology,2013,13:83.
[46]Sklan D,Noy Y.Functional development and intestinal absorption in the young poult[J].Br Poult Sci,2003,44(4):651~658.
[47]Slominski B A.Recent advances in research on enzymes for poultry diets[J].Poult Sci,2011,90(9):2013~2023.
[48]Sutivisedsak N,Leathers T D,Bischoff K M,et al.Novel sources of betaglucanase for the enzymatic degradation of schizophyllan[J].Enzyme Microb Technol,2013,52(3):203~210.
[49]SvihusB.Effectofdigestivetractconditions,feedprocessingand ingredients on response to NSP enzymes.[M].Enzymes in Farm Animal Nutrition.2nd ed,Partridge M B A G,Wallingford,UK.:CAB International,2011.129~159.
[50]Svihus B.The gizzard:Function,influence of diet structure and effects on nutrient availability[J].World’s Poult.Sci,2011(67):207~224.
[51]TahirM,SalehF,AmjedM,etal.Aneffectivecombinationof carbohydrases that enables reduction of dietary protein in broilers:importance of hemicellulase[J].Poult Sci,2008,87(4):713~718.
[52]Tang D,Hao S,Liu G,et al.Effects of maize source and complex enzymes on performance and nutrient utilization of broilers[J].Asian-Australas J Anim Sci,2014,27(12):1755~1762.
[53]Wang Z R,Qiao S Y,Lu W Q,et al.Effects of enzyme supplementation on performance,nutrient digestibility,gastrointestinal morphology,and volatile fatty acid profiles in the hindgut of broilers fed wheat-based diets[J].Poult Sci,2005,84(6):875~881.
[54]Ward N E.Choosing enzyme solution depends on many factors[J]. Feedstuff.2014,04(86).
[55]West M A C W.Assessment of dietary Rovabio Excel in practical United States broiler diets[J].J App Poult Res,2007,16(3):313~321.
[56]Wiseman J.Correlation between physical measurements and dietary energy values of wheat for poultry and pigs[J].Animal Feed Science and Technology,2000,84(1–2):1~11.
[57]Yu B,Wu S T,Liu C C,et al.Effects of enzyme inclusion in a maize–soybean diet on broiler performance[J].Animal Feed Science and Technology,2007,134(3–4):283~294.
[58]Zahura U A,Rahman M M,Inoue A,et al.Characterization of a beta-D-mannosidase from a marine gastropod,Aplysiakurodai[J].Comp BiochemPhysiol B BiochemMol Biol,2012,162(1-3):24~33.
[59]Zhang M,Jiang Z,Li L,et al.Biochemical characterization of a recombinant thermostable-mannosidase from Thermotogamaritima with transglycosidase activity[J].JournalofMolecularCatalysisB:Enzymatic,2009,60(3–4):119~124.
[60]Zhou Y,Jiang Z,Lv D,et al.Improved energy-utilizing efficiency by enzyme preparation supplement in broiler diets with different metabolizable energy levels[J].Poult Sci,2009,88(2):316~322.
[61]Zhu H L,Hu L L,Hou Y Q,et al.The effects of enzyme supplementation on performance and digestive parameters of broilers fed corn-soybean diets[J]. Poult Sci,2014,93(7):1704~1712.
Feed exogenous enzymes,as a new class of highly efficient feed additive,could eliminate adverse effects of antinutritional factors,increase the availability of nutrient by digesting the dietary components,improve bird performance and increase the economic and ecological efficiency when added to poultry diets.But the limitation of its effect was associated with enzyme,substrate,and bird factors.These variable responses,however,has limited the further development of enzyme and its wider acceptance by the market.This review aims to discussenzyme efficacy used in poultry production by documenting recent literature data on enzyme use in poultry diets,which provides scientific referencefor further study on research and reasonable application of enzyme preparations.
feed enzyme;variable response;poultry
S816.7
A
1004-3314(2016)12-0010-06
10.15906/j.cnki.cn11-2975/s.20161203
