荒漠草原中间锦鸡儿林土壤养分效应对群落组分和多样性的影响
2016-11-04安超平宋乃平随金明
安超平,王 兴,宋乃平,*,周 娟,随金明
(1 宁夏大学 西北土地退化与生态恢复国家重点实验室培育基地,西北退化生态系统恢复与重建教育部重点实验室,银川 750021;2 宁夏大学 农学院,银川 750021)
荒漠草原中间锦鸡儿林土壤养分效应对群落组分和多样性的影响
安超平1,王兴2,宋乃平1,2*,周娟1,随金明1
(1 宁夏大学 西北土地退化与生态恢复国家重点实验室培育基地,西北退化生态系统恢复与重建教育部重点实验室,银川 750021;2 宁夏大学 农学院,银川 750021)
为系统认识荒漠草原中间锦鸡儿人工林草本植被特征及演变趋势对土壤养分的响应,采用空间代替时间的方法,以天然草地为对照,构建3个中间锦鸡儿林龄(16 a、25 a、37 a)研究序列,调查不同中间锦鸡儿林灌丛内外土壤植被并分析二者之间的关系。结果表明:(1)随林龄增加,不同功能群植物重要值呈不同变化趋势,其中禾本科植物显著增加,豆科植物显著降低,杂草类植物先减小后增加。(2)α多样性分析显示,荒漠草原中间锦鸡儿人工林16 a灌丛外物种丰富度(S)、香浓维纳指数(H)最高(P<0.01);不同林龄均匀度指数(J)、优势度指数(P)未表现出明显的变化规律。(3)β多样性分析显示,Cody指数(βc)和Morisita-Horn(CMH) 相似指数均表现出16 a与25 a、37 a之间的物种差异较大、25 a与37 a之间物种差异相对较小、37 a中间锦鸡儿林与天然草地物种组成相似,βws指数在25 a中间锦鸡儿林最大。(4)人工中间锦鸡儿林存在显著的土壤碳氮“肥岛效应”,其中土壤有机碳是不同林龄植物群落组成差异的显著性影响因子,土壤速效氮为不同林龄灌丛外植物多样性差异的显著性影响因子,但灌丛内植物多样性对土壤养分的响应并不显著。研究认为,中间锦鸡儿林土壤养分演替正效应提高了植物多样性,增强了植物物种的更替程度,但建植后期(37 a)植物群落存在向天然草地演替趋势。
荒漠草原;中间锦鸡儿;土壤养分;植物群落;土壤养分效应
荒漠草原区域自然生态环境脆弱,受气候变化和人类活动干扰影响,该区域草地退化和植被可持续恢复形势极其严峻[1],严重制约了该区域农牧业和社会经济可持续发展。为遏制草原退化,近40年来在荒漠草原持续引入中间锦鸡儿,建植了大面积的人工林。以宁夏盐池荒漠草原为例,中间锦鸡儿林大规模建设始于20世纪70至80年代,2010年保有面积达到1.50×105hm2,占该县林木面积的60%。中间锦鸡儿(CaraganaintermediaKuang et H. C. Fu)具有固沙阻尘,固定氮素,保持水土等功能[2-3],是优质的灌木饲料植物资源,具有较高的生态经济价值[4],在退化草地恢复过程中发挥了巨大生态效应。中间锦鸡儿林重建促使退化草地土壤性质、植被结构发生一定程度的演变,土壤植被的空间格局显著变化[5-6]。因此,荒漠草原区人工中间锦鸡儿林土壤与植被相互关系的研究,对认识该区域人工中间锦鸡儿林的演替趋势以及稳定性,准确评价人工中间锦鸡儿林生态效益和制定合理的管理经营措施,均具有重要理论和实践意义。
土壤养分是影响群落组分和生物多样性的重要因子,但是二者之间的关系较为复杂。退化草地人工中间锦鸡儿林重建往往加速了灌丛肥岛效应不同程度的发生[2, 7],土壤肥力得到改善。土壤基质的改善和恢复一般可以促使生物群落恢复[8],但是不同功能群对于土壤养分的响应不同[9]。已有的研究表明,荒漠草原人工中间锦鸡儿林地表植被高度、盖度等数量特征和多样性随种植年限的增加呈现先增加后减小的趋势[5, 10],中林龄阶段显著恢复[6],土壤养分与植被变化间并不存在一一对应的关系。同时受外界复杂多变环境因素的影响,人工中间锦鸡儿林土壤养分的变化是否可以决定,或者说以何种方式影响植被的演变特征和方向,尚需深入的研究。
目前宁夏荒漠草原人工中间锦鸡儿林相关研究主要集中在不同种植密度中间锦鸡儿林土壤化学计量特征[11],不同种植年限土壤养分效应[12],土壤植被演变特征[5],植被动态结构[6, 10]等,对植物群落组成及多样性变化与土壤养分效应关系的研究较少,特别是在成熟林阶段,中间锦鸡儿与群落植被多样性和土壤之间的关系复杂,亟待研究。鉴于此,本研究选择3个林龄阶段类型为研究对象,通过调查每种林龄类型土壤养分和地表草本植被特征,分析地表植被群落组分及多样性与土壤养分变化规律及二者之间的关系,以期从更深层次揭示荒漠草原恢复演替过程中植物群落演变规律及其驱动因素。
1 材料和方法
1.1研究区概况
研究区位于宁夏盐池县(37°04′~38°10′N,106°30′~107°41′E),该区地处宁夏回族自治区东部,北与毛乌素沙地相连,南接黄土高原.气候属典型的中温带大陆性气候,年均气温8.1 ℃,无霜期120 d,年降水量250~350 mm,主要集中在7~9月,占全年降水量的60%以上,且年际变率大,年平均蒸发量2 710 mm。土壤类型以灰钙土为主,其次是黑垆土和风沙土,此外还有少量的盐土和白浆土等,表层土壤质地多为轻壤土、沙壤土和沙土,结构松散,肥力较低。该区主要植被类型有灌丛、草原、草甸、沙地植被和荒漠植被,其中人工灌丛、草原、沙地植被分布较广,当地荒漠草原典型建群种本氏针茅(Stipabunganea)呈散落的斑块化小面积分布,退化严重。主要草本植物有猪毛蒿(ArtemisiascopariaWaldst.)、白草(Pennisetumcentrasiaticum)、冰草[Agropyroncristatum(Linn.) Gaertn.]、草木樨状黄芪(Astragalusmelilotoides)、山苦荬(Ixerisdenticulata)、胡枝子(Lespedezabicolor)、甘草(GlycyrrhizauralensisFisch)、苦豆子(SophoraalopecuroidesL.)等。
1.2样品采集
2013年8月,选择沙质土壤的人工中间锦鸡儿林地,林龄分别为16 a、25 a和37 a,面积分别为53、40和87 hm2。中间锦鸡儿林平均密度20~45 丛/100 m-2,带距6~8 m的范围;灌丛生长情况如表1所示,其中37 a中间锦鸡儿林灌丛老化明显,土壤生物结皮发育完整,处于较为典型的退化阶段。在不同林龄(37 a、25 a、16 a)中间锦鸡儿灌丛覆盖区即灌丛内和中间锦鸡儿灌丛行间即灌丛外样地中分别随机设置5个1 m×1 m的植被调查样方,同时在天然草地随机地设置5个样方作为对照。调查指标包括:生物量、物种、高度、多度、盖度等。将各调查样方中的草本植物齐地面刈割,按样方标号分装,带回实验室称重。灌丛内、外,按照120°旋转三个方位各取 0~20 cm土壤,将采集土样去除植物根系和石块等,充分混匀后以四分法取适量作为一个样,装入塑封袋密封保存。土样带回实验室,烘干后分别研磨过筛(孔径1 mm)用于有机碳、活性碳及氮、磷含量测定。不同林龄中间锦鸡儿灌丛形态特征统计如表1。
1.3测定项目与方法
土壤样品风干后过1 mm筛,采用土壤农化分析常规方法。其中,土壤有机碳(SOC)采用重铬酸钾法容量法(外加热法)测定,土壤活性炭采用KMnO4氧化-比色法测定,土壤速效氮(EN)采用NaOH碱解扩散法测定,土壤速效磷(EP)采用NaHCO3浸提―钼锑抗比色法测定。

表1 不同林龄中间锦鸡儿灌丛形态特征(n = 15)Table 1 Morphological characters of C. intermediashrublands at different ages
植被调查采用样方法,调查每个样方中所有植物密度、盖度、高度和生物量等,同一处理的样方调查数据汇总合并,计算物种重要值、物种丰富度S、Shannon-Wiener指数(香浓维纳指数)、Pielou指数(均匀度指数)、Simpson指数(优势度指数)。
α多样性各指数计算公式如下:
种的重要值Pi=(相对盖度+相对密度+相对频度)/3
物种丰富度S=出现在样方内的物种数
Shannon-Wiener 多样性指数
S为样方中的物种数
Pielou均匀度指J=H/lnS
β多样性指数计算公式[13]如下:
Whittaker指数βws=S/ma-1
Cody 指数βc=[g(H)+I(H)]/2
经Wolda改进的Morisita-Horn指数CMH
公式中,S为不同样地出现的总物种数,ma为各样方的平均物种数;g(H)是沿生境梯度H而增加的物种数,I(H)是沿生境梯度H而失去的物种数;api和bpi为a和b样地中的第i种植物的相对重要值,aS和bS为a和b样地出现的物种总数。
1.4数据处理与析
利用SPSS17.0进行方差分析。采用CANOCO 4.5对植物多样性和环境因子进行排序。在选择排序模型前,首先对本研究数据进行DCA分析,查看排序轴的特征值,如果特征值中的最大值超过4选择单峰模型(CCA)较为合适;如果小于3选择线性模型(RDA)比较合适;如果介于3~4之间两种模型均可选择。对数据预处理后,我们选择RDA模型进行排序。然而RDA排序结果仅反映环境要素与植物群落间存在的相关关系,这种相关关系是所有环境因子综合作用的结果,不能判断出主导因子以及各主导因子与植物群落间独立的关系。因此,应用RDA分析中的前向选择来筛选环境因子中对植物群落特征变量影响最强的某一个或几个因子。前向选择分析过程中,每一个环境因子作为环境解释变量,被逐一引入模型分析,并通过蒙特卡罗检验(Monte Carlo permutation test)评价每个因子对植物的响应解释的显著性(P<0.05)。
2 结果与分析
2.1不同林龄植物群落组分变化
不同林龄中间锦鸡儿林的植物群落结构和组成发生明显的变化(表2和表3)。其中,16 a中间锦鸡儿林共有物种17种,分属于16个属、7个科;25 a中间锦鸡儿林共有物种15种,分属于17属、4科;37 a中间锦鸡儿林共有物种13种,分属14个属、6个科;天然草地共有物种13种,分属于12个属、7个科。从功能群组成来看,16 a中间锦鸡儿林灌丛内植物群落豆科植物占绝对优势,总体重要值为46.8%;16 a中间锦鸡儿林灌丛外和天然草地植物群落杂草类占绝对优势,总体重要值分别为56.59%和58.10%;25 a和37 a中间锦鸡儿林植物群落禾本科占绝对优势,总体重要值均大于63.40%(表3)。禾本科植物重要值随着林龄的增加显著增加(P<0.01),豆科植物重要值随林龄增加先增加后减小,在16 a灌丛内具有最大值,杂草类植物重要值随着林龄增加先减小后增加,在25 a中间锦鸡儿林具有最小值。
2.2不同林龄物种α和β多样性
不同林龄植被α多样性结果表明(表4):物种丰富度(S)、香浓维纳指数(H)在16 a灌丛外最大且达到显著水平(P<0.01),天然草地、25 a中间锦鸡儿林、37 a中间锦鸡儿林之间差异不显著。均匀度指数(J)、优势度指数(P)未表现出明显的变化规律。相同林龄灌丛内外植物多样性在16 a和37 a中间锦鸡儿林达到显著性差异水平(P<0.01)。

表2 不同种植年限中间锦鸡儿林灌丛内外群落物种组成和重要值Table 2 Species composition and important values (%) of the community ofC. intermedia shrublands at different ages (year)
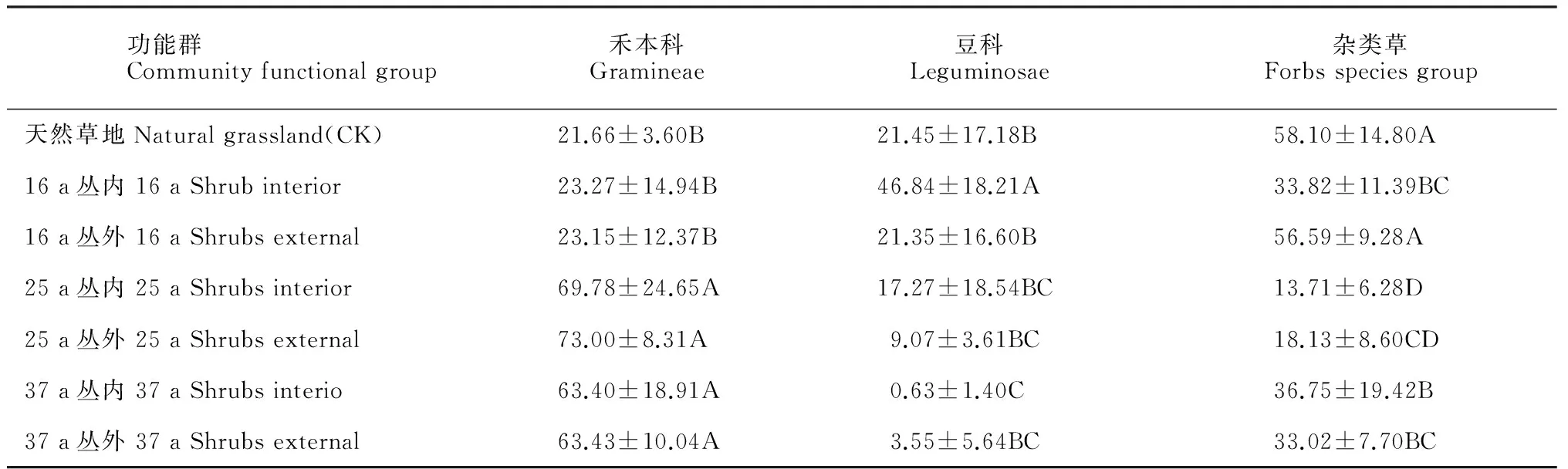
表3 不同种植年限中间锦鸡儿林灌丛内外群落功能群Table 3 Community functional group of C. intermedia shrublands at different ages (year)
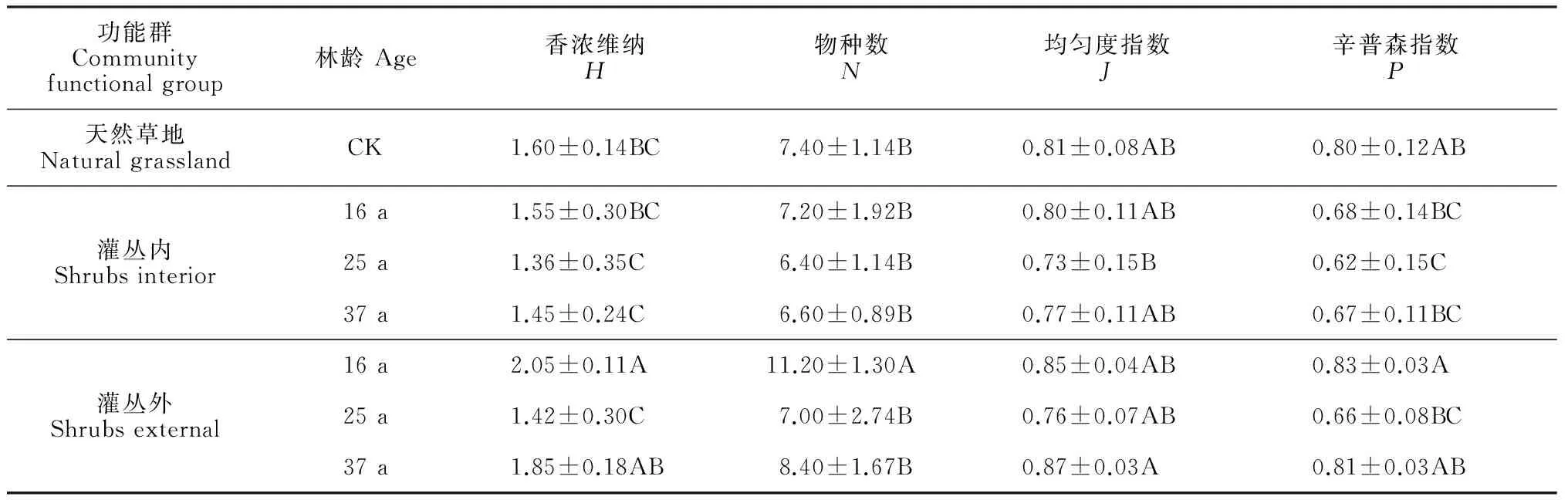
表4 不同林龄植物α多样性指数Table 4 Plant biodiversity index of Caragana intermedia shrublands at different ages (year)
不同中间锦鸡儿林Morisita-Horn指数(CMH)和Cody指数(βc)表明(表5):天然草地(CK)分别与16 a、25 a和37 a中间锦鸡儿林(灌丛内外)对比:CK与不同林龄灌丛内之间的βc值均为5,与灌丛外的βc值为3.00~4.50之间,CMH值在CK与37 a间的最大(0.54和0.52);在CK与25 a间最小(0.25和0.37)。3个林龄(灌丛内外)之间对比:16 a、25 a和37 a中间锦鸡儿林(灌丛内外)间的βc值在4.0~5之间。CMH值在37 a与25 a 间最大(0.86和0.88);在16 a与37 a间次之(0.74和0.78);在16 a与25 a间最小(0.42和0.30)。同一中间锦鸡儿林灌丛内外对比:37 a灌丛内外间CMH值最大(2.73),βc值最小(2.50);16 a中间锦鸡儿林灌丛内外间CMH值最小(0.38),βc值最大(4.00)。
Whittaker指数(βws)在1.17~2.08之间。不同林龄灌丛内外的大小分别为25 a>16 a>CK>37 a,25 a>37 a>16 a>CK。25 a中间锦鸡儿林βws指数最大(2.17和2.37),样方间物种组成差异较大,物种更替程度最大;灌丛内37a中间锦鸡儿林物种更替程度最小,样方间物种组成相似性较大;灌丛外天然草地(CK)和16 a中间锦鸡儿林物种更替程度小,样方间物种组成相似性较大。Whittaker指数(βws)在同一林龄灌丛内外比较:16 a灌丛内较高,而25 a和37 a灌丛外较高。说明人工中间锦鸡儿林建植前期灌木的覆盖可以提高林下物种的更替程度,随着林龄增加灌木对物种更替正效应由林下转移到林间。
2.3不同林龄土壤养分
不同林龄土壤养分结果表明(表6):土壤有机碳和活性碳:与林龄呈正相关,但是这种增加趋势并非简单的线性趋势。灌丛内高于灌丛外,在建植后期(25 a和37 a)灌丛内外差异显著。土壤速效磷:在不同林龄间以及灌丛内外差异均不显著。土壤速效氮:灌丛外与土壤有机碳和活性炭变化规律相同。灌丛内25 a>37 a>CK>16 a,25 a显著高于其他林地,37 a、CK和16 a三者间差异不显著。同一林龄速效氮含量在灌丛内较高,仅在25 a和37 a差异显著。

表5 不同林龄之间群落Cody指数(βc)和Morisita-Horn(CMH) 相似指数Table 5 Cody index and Morisita-Horn similarity index of C. intermedia shrublands at different ages (year)

表6 不同林龄中间锦鸡儿林土壤养分含量Table 6 Soil nutrient content of C. intermedia shrublands at different ages (year)
2.4不同林龄物种多样性与土壤养分关系
植物群落功能群、植物多样性与土壤养分(有机碳SOC、活性炭TN、速效氮AN、速效磷AP)排序结果(表7;图2,A)表明,不同林龄灌丛内植物多样性与土壤养分排序模型未达到显著水平(P>0.05),说明不同林龄灌丛内土壤养分的差异不能完全解释植物多样性随林龄的变化。植物群落功能群及不同林龄灌丛外植物多样性与土壤养分排序模型均达到显著水平(P=0.002;P=0.016),说明土壤养分很好地解释不同林龄间植物群落功能群及多样差异(表7;图1,图2,B)。不同林龄间(灌丛内外)土壤养分对植物群落功能群总解释量分别为58.6%和73.5%,对植物多样性的总解释量分别为31.3%和46.9%,RDA第一排序轴几乎包含了所有的生态信息,因此对第一排序轴展开深入分析,各个因子与排序轴第一轴的相关系数大小以及各因子箭头方向与排序轴第一轴之间的夹角大小表明(表8):群落功能群排序(灌丛内外)第一排序轴主要代表了土壤有机碳、活性炭、速效氮变化。群落多样性排序第一排序轴分别主要代表了(灌丛外)土壤速效氮、活性炭和(灌丛内)有机碳、活性碳的变化,植物群落α多样性指数与土壤养分负相关,β多样性反之。
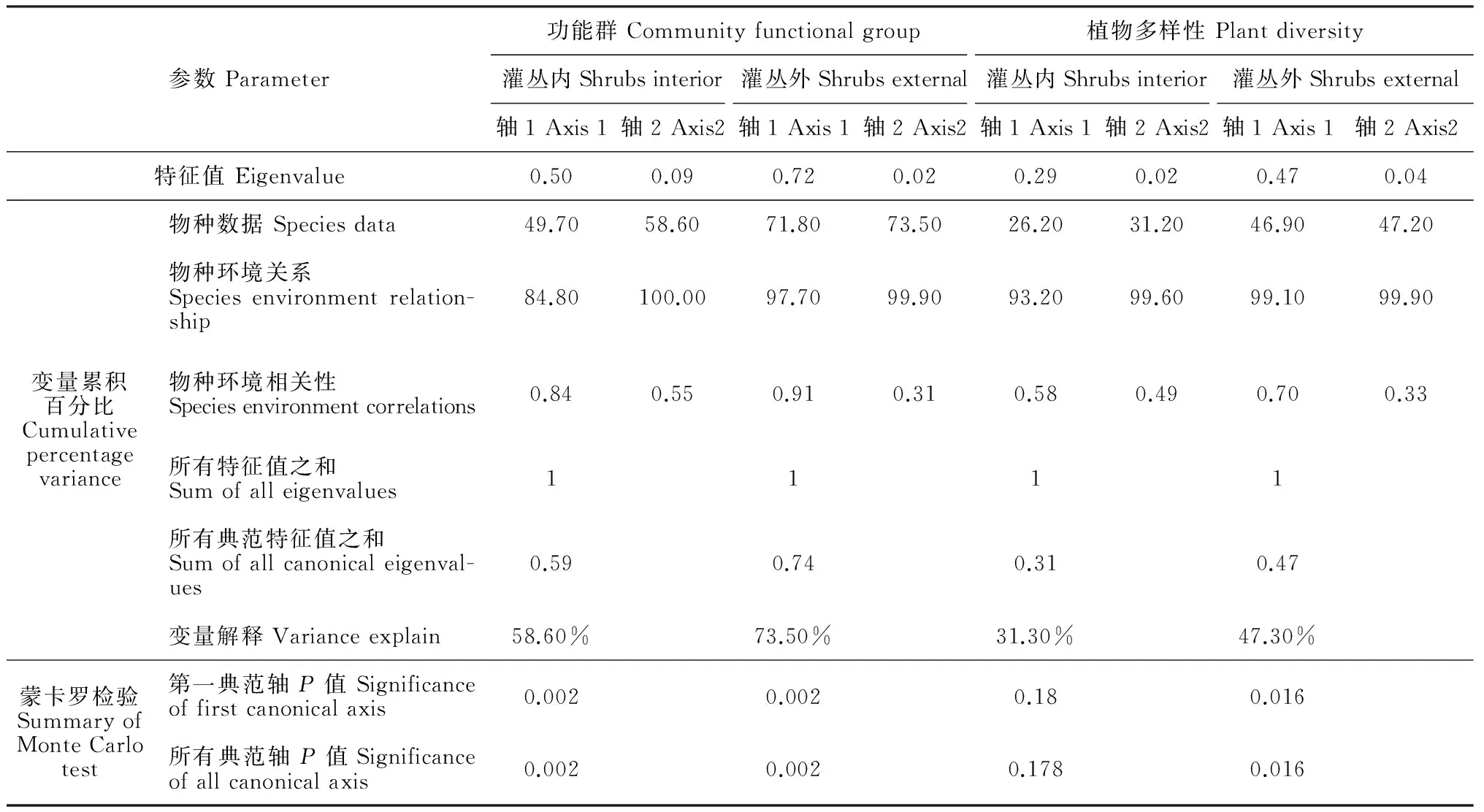
表7 植物功能群及多样性与环境因子的RDA分析结果Table 7 Redundancy analysis results of community functional group and biodiversity and environmental variables

A.灌丛内;B.灌从外,下同图1 环境因子与植物功能群RDA排序图A. Shrubs interior; B. Shrubs external. The same as belowFig.1 RDA ordination of environment factors and community functional group

图2 环境因子与植物多样性RDA排序图Fig.2 RDA ordination of environment factors and biodiversity表8 植物功能群及多样性与环境因子的相关系数Table 8 Correlation coefficients between environment characteristics and community functional group and biodiversity

参数Parameter功能群Communityfunctionalgroup植物多样性Plantdiversity灌丛内Shrubsinterior灌丛外Shrubsexternal灌丛内Shrubsinterior灌丛外Shrubsexternal轴1Axis1轴2Axis2轴1Axis1轴2Axis2轴1Axis1轴2Axis2轴1Axis1轴2Axis2土壤有机碳Soilorganiccarbon0.8990.0730.9500.0160.641-0.3490.3730.303土壤活性碳Soilactivatedcarbon0.8650.3650.942-0.0080.564-0.0810.6920.078土壤速效氮Soilactivetednitrogen0.849-0.1430.819-0.4050.342-0.5620.929-0.045土壤速效磷Soilactivetedphosphorus-0.084-0.008-0.321-0.6840.2490.2010.1890.744

表9 环境因子作为解释变量的前向选择和蒙卡罗检验分析Table 9 Forward selection with Monte Carlo permutation test of explanatory variable
上述结果体现了土壤养分综合作用与植物群落多样性的关系。进一步应用前向选择分析,每一个因子作为环境解释变量,被逐一引入模型分析(表9),并通过蒙特卡罗检验(Monte Carlo permutation test)评价其对植物变量解释的显著性(P<0.05)。不同林龄植物群落功能群变化的主要影响因子为土壤有机碳,灌丛内外土壤有机碳的单独解释量占土壤养分总解释量的67.8%(P=0.002;F=12.9)和87.8%(P=0.002;F=33.09)。不同林龄(灌丛内)RDA排序模型未达到显著水平,因此不进行前向选择分析。不同林龄(灌丛外)植物群落多样性变化影响因子为灌丛外土壤速效氮含量,其单独解释量占土壤养分总解释量的85.1% (P=0.002;F=12.25)。
3 讨 论
人工中间锦鸡儿林重建具有显著的土壤养分效应。本研究序列内,中间锦鸡儿林土壤有机碳、活性炭、速效氮得到明显恢复,基本呈现正向恢复演替趋势。土壤有机碳、活性炭和速效氮随林龄的增加而显著增加,但土壤速效磷无明显变化。与以往研究不同,土壤碳氮养分并未呈现出线性增加趋势[6],而是在中老龄林阶段(25 a)显著增长,这与杨新国等研究结果相似[5],也与干旱区自然条件下土壤质量恢复演变的时间尺度约束规律相符合[14]。同一林龄土壤碳氮含量在灌丛内高于灌丛外,在中老林龄(25 a)后表现出显著差异,说明灌丛土壤碳氮“肥岛效应”随林龄增加而增强,这与以往研究相同[15]。中间锦鸡儿对风蚀的抑制、枯枝落叶的累积、土壤结构的改善等作用使得灌丛斑块土壤比较肥沃,从而形成“肥岛效应”,这种效应会随着灌丛斑块的扩张而增强。不同林龄灌丛内速效氮含量25 a>37 a>CK>16 a,25 a显著高于其他林地,37 a,CK,16 a三者间差异不显著。说明了中间锦鸡儿在归还土壤氮的同时也在消耗土壤氮,中间锦鸡儿生长后期存在供氮不敷耗氮情况。
土壤基质的改善和恢复必然促使生物群落的恢复[9]。因此,土壤养分是影响群落组分和生物多样性的重要因子[16]。本研究结果表明:植物功能群在不同林龄间显著变化的决定性因子为土壤有机碳含量。尤其是禾本科植物总体重要值显著增加且与土壤有机碳含量高度相关,这是因为禾本科植物对土壤养分变化比较敏感[13]。豆科植物总体重要值显著降低,可能与中间锦鸡儿灌木与豆科草本之间的资源竞争有关。杂草类植物总体重要值先减小后增加可能与中间锦鸡儿自身的衰败有关。就植物多样性而言,物种数和香浓维纳指数在16 a中间锦鸡儿林显著高于其他林地,老林龄阶段(37 a)与天然草地差异不显著,均匀度指数、辛普森指数未表现出明显的变化规律,β多样性结果表明37 a与天然草地物种差异最小。伴随着中间锦鸡儿自身的退化,群落存在向天然草地演化的趋势,这与以往的研究结果相似[4, 10]。土壤养分对植物多样性的影响一直存在争议,有研究表明随土壤养分的增加物种多样性先增加后降低,但也有物种多样性降低或二者之间物相关性报道[8]。还有研究结果表明物种多样性是通过养分因子间的耦合效应有关。不同群落功能群对于土壤养分的响应不同,导致植物多样性与土壤养分关系较为复杂[13]。本研究结果表明:植物群落α多样性指数与土壤养分负相关,因为在中间锦鸡儿成熟林(25 a)以前,中间锦鸡儿对地表植被具有积极的正向调节作用,但是进入成熟林后,中间锦鸡儿林衰败与林间植被新的退化同时发生[5]。干旱区沙地人工灌木林植被在10~20年间物种数存在停滞或者下降的现象[17],黄土丘陵区中间锦鸡儿林也出现类似的现象[18]。这种现象可能与中间锦鸡儿自身对资源利用方式随时间发生变化有关,也可能是中间锦鸡儿的恢复效应随自身衰败而弱化或者丧失引起。β多样性与土壤养分正相关,说明中间锦鸡儿林对草地资源在空间上的重新分配,加速了物种的更替程度。一方面,中间锦鸡儿林自身的生态效应(防风固沙、固氮等)改善了退化草地生境条件,促使群落物种更替和植被恢复。另一方面,随中间锦鸡儿生长发育,土壤生态过程发生一定程度的改变,尤其是干旱区中间锦鸡儿林土壤水分逐年亏缺,一些隐花植物的发育阻碍降水入渗。浅层土壤丰富的水分和养分为一年生植物和浅根系半灌木的入侵和定居创造适宜条件,驱动植被向荒漠草原植被演变[19-20]。总之,无论植被是恢复演替还是逆向演替均会提高物种的更替程度和速率。土壤养分中,氮素是植物生产力首要限制资源,也是草原植物种类组成的主要因子[21]。本研究结果表明:不同林龄(灌丛外)植物多样性变化的决定性因子为土壤速效氮。氮素的增加促使优势植物快速增长,优势植物竞争作用增强,对其他物种的排斥降低植物群落多样性。本研究结果中,随林龄增加,土壤速效氮含量增加,禾本科替代豆科和杂草类植物成为优势种,导致群落多样性降低。但不同功能群的替代增强了物种的更替程度。相对于灌丛外,不同林龄(灌丛内)土壤养分并不能解释植物群落多样性的变化。灌丛内灌草间对资源的直接竞争是影响植物群落的一个重要因素,灌丛下土壤结皮的发育也可以显著推动植被的构成[20]。相对与灌丛外,灌丛内土壤养分对植物功能群和多样性变化的总解释程度较低,这一结果充分说明了灌丛内植物构成很可能是土壤养分与其他因素的综合作用,灌丛下土壤结皮、光照、水分等因素的改变是否可以决定植物群落构成改变有待进一步研究。
整体而言,土壤养分效应及正向演替趋势有利于物种的更替程度和速率,但草本植物群落存在向天然草地演替趋势;在本研究序列,植物多样性对25 a林龄响应最强烈,此林龄阶段可能是中间锦鸡儿林在恢复中的关键节点。未来研究中需将中间锦鸡儿林风蚀效应、水分分配、土壤结皮等因素与土壤养分相结合,加强老林龄阶段后的演变趋势和途径的跟踪研究。
[1]赵哈林,周瑞莲,赵学勇,等.呼伦贝尔沙质草地土壤理化特性的沙漠化演变规律及机制[J].草业学报,2012,21(2):1-7.
ZHAO H L, ZHOU R L, ZHAO X Y,etal. Desertification mechanisms and process of soil chemical and physical properties in Hulunbeir sandy grassland,Inner Mongolia[J].AtcaPrataculturaeSinica, 2012, 21(2):1-7.
[2]苏永中,赵哈林,张铜会,等.几种灌木、半灌木对沙地土壤肥力影响机制的研究[J].应用生态学报,2002,13(7):802-806.
SU Y Z, ZHAO H L, ZHANG T H,etal. Influencing mechanism of several shrubs and subshrubs on soil fertility in Keerqin sandy land[J].ChineseJournalofAppliedEcology,2002,13(7):802-806.
[3]曹成有,蒋德明,阿拉木萨,等.小叶锦鸡儿人工固沙区植被恢复生态过程的研究[J].应用生态学报,2000,11(3):349-354.
CAO C Y, JIANG D M, A L M S,etal. Ecological process of vegetation restoration inCaraganamirophyllasand-fixing area[J].ChineseJournalofAppliedEcology,2000,11(3):349-354.
[4]赵伟,杨明秀,陈林,等.荒漠草原人工柠条林草本层植被的结构与动态[J].浙江大学学报(农业与生命科学版),2015,41(6):723-731.
ZHAO W, YANG M X, CHEN L,etal. Structure and dynamics of herbaceous layer vegetation of artificialCaraganaintermediashrublands in desert steep[J].JournalofZhejiangUniversity(Agric.&life Sci).2015,41(6):723-731.
[5]刘仁涛,柴永青,徐坤,等.荒漠草原区柠条人工固沙林生长过程中地表植被-土壤的变化[J].应用生态学报,2012,23(11):2 955-2 960.
LIU R T, CHAI Y Q, XU K,etal. Variations of ground vegetation and soil properties during the growth process of artificial sand-fixingCaraganaintermediaplantations in desert steppe[J].ChineseJournalofAppliedEcology,2012,23(11):2 955-2 960.
[6]杨新国,赵伟,陈林,等.荒漠草原人工柠条林土壤与植被的演变特征[J].生态环境学报,2015,24(4):590-594.
YANG X G, ZHAO W,CHEN L,etal. Antidromal succession between soil and plant in the Caragana intermedia shrubland in the Desert Steppe[J].EcologyandEnvironmentalSciences,2015,24(4):590-594.
[7]李香真,张淑敏,邢雪荣.小叶锦鸡儿灌丛引起的植物生物量和土壤化学元素含量的空间变异[J].草业学报,2002,11(1):24-30.
LI X Z, ZHANG S M, XING X R.Spatial variation of plant biomass and soil chemical element contents induced byCaraganamicrophylla[J].AtcaPrataculturaeSinica,2002, 11(1):24-30.
[8]李新荣.干旱沙区土壤空间异质性变化对植被恢复的影响[J].中国科学(D辑:地球科学),2005,35(4):361-370.
LI X R.The effect of spatial heterogeneity changing in soil to vegetable restoration in sandy desert of arid region in China[J].ScienceChinaEarthScience, 2005,35(4):361-370.
[9]王长庭,龙瑞军,刘伟,等.高寒草甸不同群落类型土壤碳分布与物种多样性-生物量关系[J].资源科学,2010,32(10):2 022-2 029.
WANG C T,LONG R J, LIU W,etal. Relationships between soil carbon distribution and species diversity and community biomass at different alpine meadows[J].ResourcesScience,2010, 32(10):2 022-2 029.
[10]李淑君,李国旗,王磊,等.荒漠草原区不同林龄柠条林物种多样性研究[J].干旱区资源与环境,2014, 28(6), 82-87.
LI S J,LI G Q,WANG L,etal. A research on species diversity of artificialCaraganaintermediaforests in desert steppe[J].JournalofAridLandResourcesandEnvironment,2014,28(6):82-87.
[11]杨阳,刘秉如,杨新国,等.荒漠草原中不同密度人工柠条灌丛土壤化学计量特征[J].水土保持通报,2014,34(5):67-73.
YANG Y, LIU B R,YANG X G,etal. Soil stoichiometry characteristics of artificialCaraganaRorshinskiishrubs with different density in desert steppe[J].BulletinofSoilandWaterConservation,2014,34(5):67-73.
[12]宋乃平,杨新国,何秀珍,等.荒漠草原人工柠条林重建的土壤养分效应[J].水土保持通报,2012,32(4):21-26.
SONG N P,YANG X G,HE X Z,etal. Soil nutrient effect of desert steppe reconstructed by artificialCaraganamicrophyllastand[J].BulletinofSoilandWaterConservation,2012,32(4):21-26.
[13]刘晓琴,张翔,张立锋,等.封育年限对高寒草甸群落组分和物种多样性的影响[J].生态学报,2016, 36(16):1-14.
LIU X Q,ZHANG X,ZHANG L F,etal. Effects of exclosure duration on the community structure and species diversity of an alpine meadow in the Qinghai-Tibet Plateau[J].ActaEcologicaSinica,2016,36(16):1-14.
[14]张凤荣,安萍莉,王军艳,等.耕地分等中的土壤质量指标体系与分等方法[J].资源科学,2002, 24(2):71-75.
ZHANG F R,AN P L,WANG J Y,etal. Soil quality criteria and methodologies of farmland grading[J].ResourcesScience,2002, 24(2):71-75.
[15]潘军,安超平,吴旭东,等.荒漠草原2种锦鸡儿灌丛化过程中土壤养分分布规律[J].水土保持学报,2015,29(6):131-136.
PAN J,AN C P,WU X D,etal. Distribution pattern of soil nutrients in the thicketization of 2 Types of caragana in Desert steppe[J].JournalofSoilandWaterConservation,2015,29(6):131-136.
[16]HUANG P M, WANG S L, TZOU Y M,etal. Physicochemical and biological interfacial interactions: impacts on soil ecosystem and biodiversity[J].EnvironmentalEarthSciences, 2013, 68(8):2 199-2 209.
[17]李新荣,张景光,刘立超,等.我国干旱沙漠地区人工植被与环境演变过程中植物多样性的研究[J].植物生态学报,2000, 24(3):257-261.
LI X R,ZHANG J G,LIU L C,etal. Plant diversity in the process of succession of artificial vegetation types and environment in an arid desert region of China[J].ActaPhytoecologicaSinica,2000, 24(3):257-261.
[18]王国梁,刘国彬,周生路,等.黄土丘陵沟壑区小流域植被恢复对土壤稳定入渗的影响[J].自然资源学报,2003,18(5):529-535.
WANG G L,LIU G B,ZHOU S L,etal. The effect of vegetation on soil stable infiltration rates in small watershed of loess gully region[J].JournalofNaturalResources,2003,18(5):529-535.
[19]李新荣,张志山,黄磊,等.我国沙区人工植被系统生态-水文过程和互馈机理研究评述[J].科学通报,2013,58(5-6):397-410.
LI X R,ZHANG Z S,HUANG L,etal. Review of the ecohydrological processes and feedback mechanisms controlling sand-binding vegetation systems in sandy desert regions of China[J].ChineseSci.Bull.,2013,58(5-6):397-410.
[20]李新荣,张志山,王新平,等.干旱区土壤植被系统恢复的生态水文学研究进展[J].中国沙漠,2009,29(5):846-852.
LI X R,ZHANG Z S,WANG X P,etal. The ecohydrology of the soil vegetation system restoration in arid zones: A review[J].JournalofDesertResearch,2009,29(5):846-852.
[21]高英志,韩兴国,汪诗平,等.放牧对草原土壤的影响[J].生态学报,2004,24(4):790-794.
GAO Y Z, HAN X G,WANG S P.The effects of grazing on grassland soils[J].ActaEcologicaSinica,2004, 24(4):790-794.
(编辑:潘新社)
Effect of Soil Nutrient on Community Composition and Diversity ofCaraganaintermediain Desert Steppe
AN Chaoping1, WANG Xing2, SONG Naiping1,2*, ZHOU Juan1, SUI Jinming1
(1 Breeding Base for State Key Laboratory of Land Degradation and Ecological Restoration in Northwestern China, Key Laboratory for Restoration and Reconstruction of Degraded Ecosystem in Northwestern China of Ministry of Education, Ningxia University, Yinchuan 750021, China; 2 School of Agriculture, Ningxia University, Yinchuan 750021, China)
In order to understanding the characteristics and evolution trend ofCaraganaintermediaplantation herbaceous vegetation systematically and the response of soil nutrients in desert steppe, we used spatiotemporal substitution method, toke natural meadow as control, constructed threeC.intermediaage as research sequence, and investigated inside and outside of different soil vegetation and shrubs and analyzed the relationship between them. The results showed that: (1)with the increasing of the planting years ofC.intermedia, different groups of plants showed different trends to the overall important values: a significant increase in grasses, a significantly reduce in legume, and the weed species increase after the first decrease.(2) α diversity: the biodiversity index including Richness (S) Shannon Wiener index (H) had a highest value in 16a outside shrub (P<0.01). Evenness index (J) and Dominance index (P) did not showed significant trend in different stand ages. (3) β diversity: The change of Cody index(βc)and Morisita-Horn(CMH) similarity index indicated that: the species of 16 a, 25 a and 37 a were quite different, while the differences were relatively small between 25 a and 37 a. The composition between 37 aCaraganaforest and natural grassland species were similar. Theβwsindex had the highest value in 25 a. (4)There was a significant “fertile island effect” of soil carbon and nitrogen inCaraganaintermediaforest. The soil organic carbon had a significantly influence on community group for different artificialCaraganaforest. The effect of soil nitrogen on plant diversity was not significant in shrubs interior while was significant in shrubs external. The study illustrated that: the positive effect of soil nutrient succession could improve the plant diversity inC.intermediaforest and enhance the degree of replacement of plant species. However, plant community succession had a trend from presence to natural meadow on later (37 a). The vegetation succession direction ofC.intermediaforest was uncertain.
desert steppe region;Caraganaintermedia; soil nutrients; plant community; soil nutrient
1000-4025(2016)09-1872-10doi:10.7606/j.issn.1000-4025.2016.09.1872
2016-07-23;修改稿收到日期:2016-08-13
国家自然科学基金(41461046);国家重点基础研究发展计划前期专项(2012CB723206);宁夏大学211项目
安超平(1988-),男,在读硕士研究生,主要从事生态恢复研究,E-mail: 646218886@qq.com
宋乃平,教授,博士生导师,主要从事土地利用与生态过程研究,E-mail: songnp@163.com
Q948.15+4
A
