不同起源云杉林土壤有机碳密度及与物种多样性关系
2016-10-25关晋宏张克栋张建国何秋月盛1
关晋宏, 张克栋, 邓 磊, 张建国, 何秋月, 杜 盛1,
(1.中国科学院 水利部 水土保持研究所, 陕西 杨凌 712100; 2.西北农林科技大学水土保持研究所, 陕西 杨凌 712100; 3.西北农林科技大学 西部环境与生态教育部重点实验室,陕西 杨陵 712100; 4.甘肃省洮河林业局, 甘肃 甘南 747600; 5.中国科学院研究生院, 北京 100049)
不同起源云杉林土壤有机碳密度及与物种多样性关系
关晋宏1,5, 张克栋4, 邓 磊3, 张建国1,5, 何秋月2, 杜 盛1,2
(1.中国科学院 水利部 水土保持研究所, 陕西 杨凌 712100; 2.西北农林科技大学水土保持研究所, 陕西 杨凌 712100; 3.西北农林科技大学 西部环境与生态教育部重点实验室,陕西 杨陵 712100; 4.甘肃省洮河林业局, 甘肃 甘南 747600; 5.中国科学院研究生院, 北京 100049)
对甘南地区云杉人工林和天然林的不同林龄(包括幼龄林、中龄林、近熟林和成熟林)土壤有机碳密度及其群落物种多样性的关系进行研究。结果表明:不同起源云杉林在各林龄中均以表层土壤(0—10 cm)土壤有机碳含量最高,随土壤深度的增加逐渐减少;土壤有机碳密度除天然云杉成熟林在表层最高外,其他均以深层土壤(50—100 cm)最高,且土壤有机碳含量和有机碳密度随林龄的变化不显著(p>0.05)。冗余分析表明,人工林0—10,10—20,20—30,30—50 cm的土壤有机碳密度除与Ea指数呈负相关关系外,与丰富度(S)、Shannon-Wiener指数(H′)、Simpson指数(D)、Pielou指数(Jsw)均呈正相关关系;天然林0—10,10—20,20—30 cm的土壤有机碳密度与这5个指数均呈正相关关系。
云杉; 人工林; 天然林; 林龄; 土壤有机碳密度; 物种多样性
土壤有机碳库是陆地生态系统最大的碳库,贮有1 550 Pg C,约是陆地植被碳库的2~3倍[1]。森林作为陆地生态系统的主体,其土壤碳库约占陆地土壤碳库的73%[2]。土壤碳库的微小变化,都会引起大气CO2浓度的巨大变化,而导致其在全球碳循环中源、汇、库的转化[3],因此,土壤有机碳在全球气候变化下的碳循环研究中显得尤为重要。
近年来,许多学者从全国[4-5]及区域尺度[6-7]对土壤碳库进行了估算,但结果差异较大,且在区域尺度上存在一定的空间异质性。土地利用变化被认为是全球除化石燃料燃烧之外的第二大碳源,并影响着土壤的碳源、汇功能[8]。森林演替过程中,土壤有机碳的来源主要是地上部分的枯落物输入以及地下根系周转产生的碎屑,而物种的组成影响着土壤碳输入的数量和质量,从而影响森林生态系统土壤碳汇(源)功能。由于缺乏长期的定位观测数据,目前对土壤有机碳随林龄的变化研究较少,部分学者认为土壤有机碳会随林龄的增加呈现增长趋势,尤其表现在人工林中[9],也有学者认为区域尺度的土壤有机碳主要受土壤类型的影响,而受林龄的影响较小[10],总之,土壤有机碳随林龄的变化并无定论。
云杉林是甘南地区高海拔地带性植被的建群种,也是该区人工造林的主要树种,可以发育成稳定的亚高山针叶林顶级群落,是该区生物多样性最丰富、生产力最高的植被类型之一。由于20世纪六、七十年代人类不合理的乱砍滥伐,导致云杉面积大幅度下降,自“天保工程”以来,云杉林的生境有所恢复,面积大幅增加,年龄结构也发生了改变。对不同起源云杉林的土壤有机碳进行研究,是预测全球气候变化的一项重要的基础性工作。为此,在样地调查的基础上,本文分析比较了不同起源云杉林土壤有机碳密度及其群落物种多样性随林龄的变化,并尝试用物种多样性解释土壤有机碳对土地利用变化的响应,以期为区域尺度土壤碳库的精确估算及森林生态系统的经营提供参考。
1 研究区概况与研究方法
1.1研究区概况
甘南藏族自治州位于甘肃省西南部,33°06′—35°44′N,100°46′—104°44′E,青藏高原东北缘,平均海拔3 000 m。该区植被覆盖度较高,属典型的高原大陆性气候,高寒阴湿,多年平均降雨量为400~800 mm,降雨季节性明显,主要集中在7—9月。年平均气温仅1~3℃,年极端最高气温为33.8℃,年极端最低气温为-29.6℃,≥10℃的积温持续不足3个月,全年平均日照时数2 200~2 400 h[11]。云杉林为该区面积分布最广的亚高山针叶林,土壤类型以山地棕壤和山地褐土为主。
1.2研究方法
1.2.1样地设置在2011年,2012年植物生长旺盛期(7月,8月),选择云杉人工林和天然林群落设置样地,其中,人工林样地13块,林龄分布18~65 a;天然林样地11块,林龄分布34~140 a。林龄的确定主要以生长锥钻芯取样并结合当地林业局的相关资料得出。由于该区地貌以山地和高原为主,且地形较为破碎,根据地形条件,样地设置为50 m×20 m或30 m×20 m,每个样地内分别沿对角线设置3个2 m×2 m灌木样方和3个1 m×1 m草本样方。林内乔木层中云杉占绝对优势地位,林下灌木、草本发育良好。不同起源云杉林林分特征见表1。
1.2.2物种多样性调查对标准地内胸径大于2 cm,高度大于2 m的乔木进行每木检尺,测定胸径和树高。灌木和草本样方内,分别记录植物的种类、个体数(丛数)、高度和盖度等。根据样地调查所得不同物种相对盖度、相对多度和相对频度统计出各物种的重要值,再计算出群落的多样性指数,如物种丰富度指数(S),物种多样性指数Simpson指数(D)、Shannon-Wiener指数(H′)和均匀度指数Pielou指数(Jsw)、Alatalo指数(Ea),具体计算方法[13]如下:
H′=-∑PilnPi
Jsw=H′/lnS
式中:S——样方出现的物种数,即丰富度指数;Pi——相对重要值。
1.2.3容重测定在调查样地内选择未受人为干扰,植被结构和土壤具代表性的地段,挖掘土壤剖面,深至100 cm,不够100 cm至基岩为止。按照0—10,10—20,20—30,30—50,50—100 cm不同深度,每层用环刀取2个重复土样。野外取回土样后带回实验室105℃烘干24 h至恒重,并对每个土样内>2 mm的砾石称重,计算出土壤大于2 mm砾石所占体积百分比(%),根据环刀的容积(100 cm3)和烘干土的质量计算各土层土壤容重及土壤有机碳密度。
土壤有机碳密度(t/hm2)计算公式为:

式中:Ci——第i层土壤有机碳含量(g/kg);BDi——第i层土壤平均容重(g/cm3);Hi——第i层土壤厚度(cm),Ri——第i层土壤大于2mm砾石所占体积百分比(%)。

表1 不同起源云杉林林分特征
注:人工林和天然林林龄的划分参考甘肃省林业厅[12],天然林林龄为样地内标准木的平均年龄。
1.2.4土壤有机碳含量测定在调查样地内使用内径5 cm的土钻,按0—10,10—20,20—30,30—50,50—100 cm五个层次,随机重复取样3次,均匀混合成一个混合样。土壤尽快自然风干,去除砾石和植物根系等杂物,磨碎后过0.25 mm筛,采用重铬酸钾—硫酸氧化外加热测定土壤有机碳含量[14]。
1.2.5数据处理所有数据均为各处理结果的平均值,并用SPSS 16.0对其进行统计学检验。分不同起源,分别对同一林龄组不同土层、同一土层不同林龄组间土壤有机碳含量和有机碳密度,以及同一起源不同林龄间土壤总有机碳密度的关系采用单因素方差分析(One-way ANOVA)和最小显著差异法(LSD)进行相关性检验,α=0.05作为差异显著性检验的阈值。
为充分比较物种多样性对土壤有机碳密度的影响,本研究中同一林龄的样地尽可能选择多个重复,但由于甘南林区破坏严重,且地形较为破碎,各林龄样地数量不一致。天然云杉成熟林在调查中仅有一个样地,因此各类指标无重复,也未对其土壤有机碳含量、碳密度、物种多样性指数与其他林龄间进行统计学比较。
2 结果与分析
2.1不同起源云杉林土壤有机碳含量随林龄的变化
在不同起源云杉林中,不同林龄云杉林的土壤有机碳含量均以表层(0—10 cm)土壤最高,随土层的增加逐渐降低,土层20 cm以下的土壤有机碳含量变化范围较小(图1)。对不同深度的土壤有机碳含量比较发现,0—10 cm的土壤有机碳含量与10—20 cm的在各林龄间均无显著差异(p>0.05),而与深层土壤均存在显著差异(p<0.05)。20—30,30—50,50—100 cm深层土壤之间的土壤有机碳密度均无显著差异(p>0.05)。不同起源云杉林中,同一土层的土壤有机碳含量受林龄的影响均不显著(p>0.05)。
2.2不同起源云杉林土壤有机碳密度随林龄的变化
在不同起源云杉林中,随着土层深度的增加,土壤有机碳密度均出现先降低后升高的趋势(图2)。在云杉人工林中,方差分析表明同一土层不同林龄土壤有机碳密度之间均无显著差异(p>0.05),同一林龄不同土层的土壤有机碳密度之间也均无显著差异(p>0.05)。在云杉人工林中,各层土壤碳密度分配不均,除幼龄林为10—20 cm的土壤有机碳密度最高外,其余均为深层(50—100 cm)土壤有机碳密度最高。同一林龄土壤表层有机碳密度占土壤总有机碳密度变化波动较大,为13.14%~20.28%,说明土壤表层碳密度的变化对人为干扰等环境变化的响应更为敏感。
云杉天然林中,各土层土壤有机碳密度受林龄的影响也均不显著(p>0.05),而同一林龄不同土层间仅在幼龄林的表层(0—10 cm)土壤有机碳密度与深层(50—100 cm)土壤有机碳密度之间有显著差异(p<0.05),其余土层之间差异不显著(p>0.05)。除成熟林土壤有机碳密度主要集中在表层外,云杉天然林其他林龄的土壤有机碳密度主要集中分布在深层土壤,占土壤总有机碳密度的29.26%~41.25%。
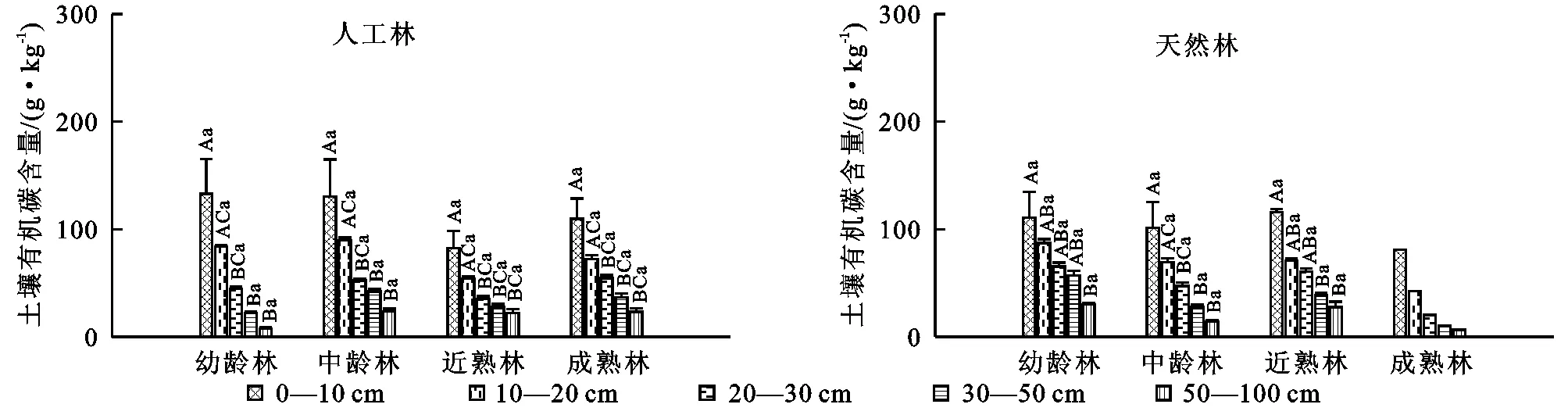
注:不同大写字母表示同一林龄不同土层土壤有机碳含量差异显著,不同小写字母表示同一土层不同林龄土壤有机碳含量差异显著。
图1不同起源云杉林土壤有机碳含量随林龄的变化
2.3不同起源云杉林土壤总有机碳密度随林龄的变化
不同起源云杉林土壤总有机碳密度(0—100 cm)随林龄的变化差异见图3。在云杉人工林中,土壤总有机碳密度随林龄的增加呈现波动性增加的趋势,以中龄林最高,为392.24 t/hm2,土壤总有机碳密度随林龄的变化总体表现为:中龄林(392.24 t/hm2)>成熟林(381.85 t/hm2)>近熟林(324.17 t/hm2)>幼龄林(327.92 t/hm2)。在云杉天然林中,土壤总有机碳密度呈现出波动式降低,以幼龄林最高(459.79 t/hm2),土壤总有机碳密度随林龄的变化为:幼龄林(459.79 t/hm2)>近熟林(431.75 t/hm2)>中龄林(299.92 t/hm2)>成熟林(236.41 t/hm2),且不同起源云杉林的土壤总有机碳密度受林龄的影响均不显著(p>0.05)。
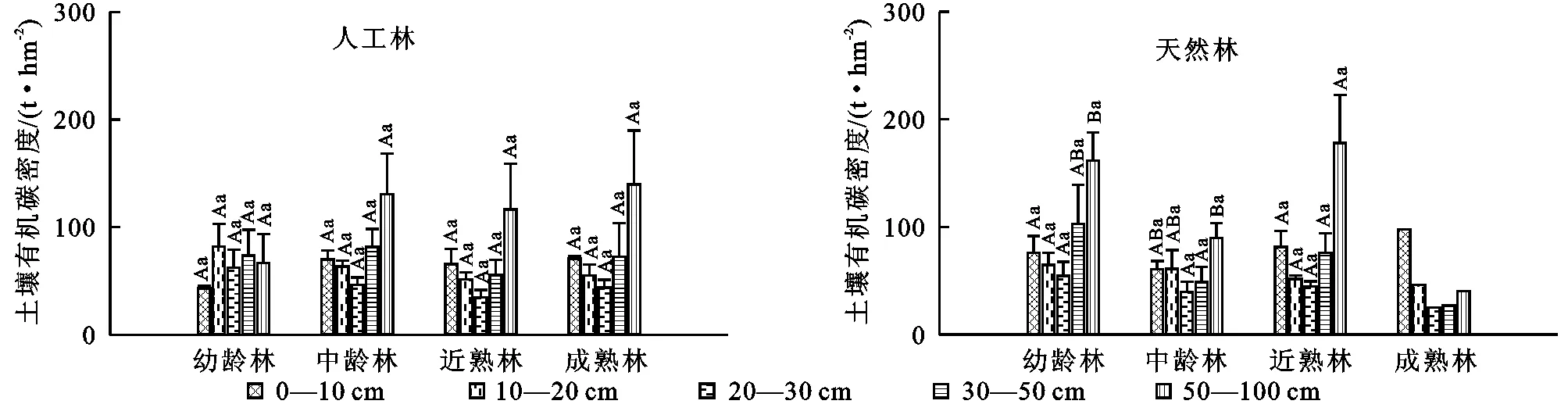
注:不同大写字母表示同一林龄不同土层土壤有机碳密度差异显著,不同小写字母表示同一土层不同林龄土壤有机碳密度差异显著。
图2不同起源云杉林土壤有机碳密度随林龄的变化

图3 不同起源云杉林土壤总有机碳密度随林龄的变化
2.4不同起源云杉林物种多样性随林龄的变化
为阐述不同起源云杉林物种多样性随林龄的变化关系,分别统计了丰富度指数(S),Shannon-Wiener指数(H′),Simpson指数(D)和均匀度指数Pielou指数(Jsw),Alatalo指数(Ea),综合表示为图4。对云杉人工林来说,各物种多样性指数均表现出从幼龄林到近熟林增加,近熟林到成熟林减小的趋势,但各指数之间受林龄的影响并不显著(p>0.05)。对云杉天然林来说,各物种多样性指数均为先降低后增加,除Ea指数外,其他各指数均受林龄的影响显著(p<0.05)。
2.5物种多样性对土壤有机碳密度的影响
植被演替主要通过物种之间竞争来推动,土壤作为植被演替过程中重要的环境因子,影响着物种的定植与生存。分别对云杉人工林和天然林的群落物种多样性指数和不同深度土壤有机碳密度进行冗余分析(RDA),结果如图5。在云杉人工林中,前两个组分量的累计贡献率为81%,其中前两轴分别解释58.7%和22.3%(图5a)。云杉人工林中,0—10,10—20,20—30,30—50 cm的土壤有机碳密度与S指数,H′指数,D指数,Jsw指数呈正相关关系,与Ea呈负相关关系;50—100 cm的土壤有机碳密度与S指数,H′指数,D指数,Jsw指数呈负相关关系,与Ea呈正相关关系,也就是说群落物种丰富度指数(S)、多样性指数(H′、D)和均匀度指数(Jsw)积极的影响0—50 cm的土壤有机碳密度。

注:不同大写字母表示人工林不同林龄物种多样性指数差异显著,不同小写字母表示天然林不同林龄物种多样性指数差异显著。
图4不同起源云杉林群落物种多样性指数随林龄的变化
在云杉天然林中,前两个组分量的累计贡献率为89%,其中前两轴分别解释69%和20%(图5b)。0—10,10—20,20—30 cm的土壤有机碳密度与S指数,H′指数,D指数,Jsw指数和Ea指数呈正相关关系,而与30—50,50—100 cm的土壤有机碳密度呈负相关关系,也就是说群落物种多样性指数积极的影响0—30 cm的土壤有机碳密度。

图5 不同起源云杉林土壤有机碳密度与物种多样性指数的RDA排序结果
3 讨论与结论
本文对甘南地区不同起源云杉林土壤有机碳密度随林龄的变化进行了研究,并与群落物种多样性之间的关系做了初步探讨。研究表明,不同林龄人工云杉林和天然云杉林土壤有机碳密度的变化范围分别为324.17~392.24 t/hm2和236.41~459.79 t/hm2。Gao等[15]研究得出宁夏地区48 a青海云杉人工林土壤有机碳密度为411 t/hm2,略高于本文云杉人工林土壤有机碳密度均值(356.55 t/hm2),二者均高于我国人工林平均土壤有机碳密度(107.1 t/hm2)[16]。阿米娜木·艾力等[17]研究了天山云杉在海拔(1 800~2 800 m)差异下,土壤有机碳密度变化范围为285.9~488.7 t/hm2,平均382.05 t/hm2,略高于本文云杉天然林土壤有机碳密度的均值(356.97 t/hm2),二者均高于我国天然林平均土壤有机碳密度(109.1 t/hm2)[16],说明甘南地区云杉林不论天然林还是人工林土壤有机碳密度均偏高。在气候条件一致的情况下,不同物种的根系分布以及凋落物的累积分解输入是土壤有机碳含量及碳密度发生变化的主要原因[18]。其次,由于云杉林位于高海拔地区,温度低、土壤相对湿度大、冻结时间长,土壤异养呼吸受到限制,有利于土壤有机质的积累,使得该区云杉林土壤有机碳密度较高,成为我国森林生态系统重要的土壤碳库。
目前关于土壤有机碳密度随林龄变化的结论存在分歧,有学者认为在无人为干扰的森林中,土壤有机碳含量和有机碳密度均随林龄的增加逐渐增大[9,19],也有学者认为林龄与土壤有机碳密度之间无显著差异[10,20]。本研究中,云杉人工林土壤总有机碳密度随林龄呈现波动性增加的趋势,云杉天然林呈现波动性降低的趋势。云杉天然林波动性降低的原因可能是本研究中天然林成熟林重复样地较少,由于六、七十年代以来甘南林区采伐严重,研究能到达的采样区内天然云杉成熟林面积较小;加之受采伐等人为活动的干扰,影响了凋落物的分解与输入,从而引起土壤有机碳的降低。尽管“天保工程”实施以来,甘南林区森林得到大面积改善,但土壤理化性质的改变是长期而缓慢的过程。人工林土壤有机碳密度的差异主要归因于细根的输入而不是地表凋落物的累积分解[21],本研究中云杉人工林幼龄林土壤有机碳密度小于中龄林和成熟林时期,与Jug等人[22]研究结果相近,即造林初期土壤有机碳密度会下降,之后开始慢慢积累。
土壤有机碳密度受气候变化、不同森林类型、年龄结构、森林管理等因素的影响外,森林群落的物种多样性也是保证土壤有机质增加的重要因素[16,23]。森林演替导致凋落物不断积累,凋落物分解后归还到土壤中,提高了土壤基质的稳定性,为更多物种的生存、定居、生长发育提供良好的条件,进而促进森林群落的演替以及生物多样性的维持[24]。但关于土壤有机碳密度与群落物种多样性的关系一直缺乏研究,本文尝试性的解释不同起源云杉林土壤有机碳密度与群落物种多样性的关系,结果表明,云杉人工林0—50 cm的土壤有机碳密度与丰富度指数(S)、多样性指数(H′、D)以及均匀度指数(Jsw)存在正相关关系,换句话说,增加乔、灌、草各层物种种类,并优化群落均匀度,使物种个体数目的分配更加均匀,能改善云杉人工林的土壤有机碳密度。云杉天然林0—30 cm的土壤有机碳密度与丰富度、多样性指数以及均匀度指数都有正相关关系,也就是说,提高群落物种多样性能改善云杉天然林的土壤有机碳密度。
综上,甘南地区云杉林土壤有机碳密度较高,是我国森林生态系统重要的土壤碳库之一。土壤有机碳密度与群落物种多样性的关系表明,今后该区的云杉森林生态系统管理中,对人工林而言,要采取一定的抚育措施,提高林地稳定性和可持续更新的潜力,进而促进表层土壤碳密度的增加;天然林在保证现有林地生境的基础上,保持乔、灌、草各层物种的多样性配制,可改善表层土壤有机碳密度。
[1]Turner B L, Clark W C, Kates R W, et al. The Earth as Transformed by Human Action:Global and Regional Changes in the Biosphere over the Past 300 Years[M]. New York:Cambridge University Press,1990.
[2]Dixon R K, Solomon A, Brown S, et al. Carbon pools and flux of global forest ecosystem[J]. Science,1994,265(5169):171-171.
[3]Smith P, Fang C M, Dawson J C, et al. Impact of global warming on soil organic carbon[J]. Advances in Agronomy,2008(7):1-43.
[4]潘根兴.中国土壤有机碳和无机碳库量研究[J].科技通报,1999,15(5):330-332.
[5]王绍强,周成虎,李克让,等.中国土壤有机碳库及空间分布特征分析[J].地理学报,2000,55(5):533-544.
[6]刘京,常庆瑞,陈涛,等.陕西省土壤有机碳密度空间分布及储量估算[J].土壤通报,2012,43(3):656-661.
[7]黄从德,张健,杨万勤,等.四川森林土壤有机碳储量的空间分布特征[J].生态学报,2009,29(3):1217-1225.
[8]Guo L B, Gifford R M. Soil carbon stocks and land use change:a meta analysis[J]. Global Change Biology,2002,8(4):345-360.
[9]苗娟,周传艳,李世杰,等.不同林龄云南松林土壤有机碳和全氮积累特征[J].应用生态学报,2014,25(3):625-631.
[10]魏亚伟,于大炮,王清君,等.东北林区主要森林类型土壤有机碳密度及其影响因素[J].应用生态学报,2013,24(12):3333-3340.
[11]邹德富,冯琦胜,梁天刚.甘南地区植被类型及其NPP研究[J].遥感技术与应用,2011,26(5):577-583.
[12]国家林业局.森林资源规划设计调查主要技术规定[S].GB/T 26524—2010.
[13]马克平,刘玉明.生物群落多样性的测度方法[J].生物多样性,1994,2(4):231-239.
[14]刘光崧.土壤理化分析与剖面描述[M].北京:中国标准出版社,1996.
[15]Gao Y, Cheng J M, Ma Z R, et al. Carbon storage in biomass, litter, and soil of different plantations in a semiarid temperate region of northwest China[J]. Annals of Forest Science,2014,71(4):427-435.
[16]刘世荣,王晖,栾军伟.中国森林土壤碳储量与土壤碳过程研究进展[J].生态学报,2011,31(19):5437-5448.
[17]阿米娜木·艾力,常顺利,张毓涛,等.天山云杉森林土壤有机碳沿海拔的分布规律及其影响因素[J].生态学报,2014,34(7):1626-1634.
[18]Zhou G Y, Guan L L, Wei X H, et al. Litterfall production along successional and altitudinal gradients of subtropical monsoon evergreen broadleaved forests in Guangdong, China[J]. Plant Ecology,2007,188(1):77-89.
[19]Luan J W, Liu S R, Zhu X L, et al. Soil carbon stocks and fluxes in a warm-temperate oak chronosequence in China[J]. Plant and Soil,2011,347(1-2):243-253.
[20]Fonseca W, Benayas J M R, Alice F E. Carbon accumulation in the biomass and soil of different aged secondary forests in the humid tropics of Costa Rica[J]. Forest Ecology and Management,2011,262(8):1400-1408.
[21]Wang H, Liu S R, Mo J M, et al. Soil organic carbon stock and chemical composition in four plantations of indigenous tree species in subtropical China[J]. Ecological Research,2010,25(6):1071-1079.
[22]Jug A, Makeschin F, Rehfuess K, et al. Short-rotation plantations of balsam poplars, aspen and willows on former arable land in the Federal Republic of Germany:Ⅲ. Soil ecological effects[J]. Forest Ecology and Management,1999,121(1/2):85-99.
[23]Schleuß P-M, Heitkamp F, Leuschner C, et al. Higher subsoil carbon storage in species-rich than species-poor temperate forests[J]. Environmental Research Letters,2014,9(1):14007-14009.
[24]崔宁洁,张丹桔,刘洋,等.不同林龄马尾松人工林林下植物多样性与土壤理化性质[J].生态学杂志,2014,33(10):2610-2617.
Soil Organic Carbon Density and Relationship with Species Diversity in Pedigreed Spruce at Natural Forest and Plantation Scales
GUAN Jinhong1,5, ZHANG Kedong4, DENG Lei3, ZHANG Jianguo1,5, HE Qiuyue2, DU Sheng1,2
(1.Institute of Soil and Water Conservation, Chinese Academy of Sciences and Ministry of WaterResources,Yangling,Shaanxi712100,China; 2.InstituteofSoilandWaterConservation,NorthwestA&FUniversity,Yangling,Shaanxi712100,China; 3.KeyLaboratoryofEnvironmentandEcologyofEducationMinistryinWestChina,NorthwestA&FUniversity,Yangling,Shaanxi712100,China; 4.GansuTaoheForestBureau,Gannan,Gansu747600,China; 5.UniversityofChineseAcademyofSciences,Beijing100049,China)
Different-aged forests (including young forest, middle-aged forest, pre-mature forest and mature forest) of both plantations and natural forests of spruce (Piceaasperata) were systematically investigated in Gannan Tibetan Autonomous Prefecture to explore the relationship between species diversity and soil organic carbon (SOC) density. Vertical distribution of SOC content and density, and species diversity, in both plantations and natural forests were analyzed. The results showed that SOC contents were highest in the top soil in different-aged forests of both natural forests and plantations, and decreased gradually with soil depth. SOC densities were highest in the deep soil layer(50—100 cm)in different-aged forests of both natural forests and plantations except the natural mature forest. Moreover, SOC content and density varied insignificantly with forest ages(p>0.05). SOC density in spruce plantations increased, whereas SOC density in spruce natural forests decreased along the ages. Redundancy discriminate analysis (RDA) showed that SOC densities of 0—10, 10—20, 20—30, 30—50 cm in spruce plantations had a positive relationship with species richness (S), Shannon-Wiener (H′), Simpson (D) and Pielou (Jsw), but a negative relationship with Alatalo (Ea). By contrast, SOC densities of 0—10, 10—20 and 20—30 cm in spruce natural forests had a positive relationship with the five indices mentioned above.
Piceaasperata; planations; natural forests; forest age; soil organic carbon density; species diversity
2015-05-07
2015-05-12
中国科学院战略性先导科技专项(XDA05050202);国家自然科学基金(41171419,41411140035)
关晋宏(1984—),女,山西宁武人,博士研究生,主要研究方向为森林生态、流域生态与管理。E-mail:cocogjh@163.com
杜盛(1965—),男,内蒙古鄂尔多斯人,博士,研究员,主要研究方向为森林生态、流域生态与管理。E-mail:shengdu@ms.iswc.ac.cn
S718.5
A
1005-3409(2016)02-0049-06
