入侵植物乙醇提取物对福寿螺胆碱酯酶活性的影响
2016-10-19邹湘辉谢东吴丽娜查广才孙延杰
邹湘辉 谢东 吴丽娜 查广才 孙延杰
摘要:探讨了入侵植物乙醇提取物对福寿螺(Pomacea canaliculata)乙酰胆碱酯酶(Acetylcholinesterase,AChE)和丁酰胆碱酯酶(Butyrylcholinesterase,BChE)活性的影响。采用乙醇提取法获得薇甘菊(Mikania micrantha)、南美蟛蜞菊(Wedelia trilobata)和五爪金龙(Ipomoea cairica)3种入侵植物茎和叶组织活性物质,改良Ellam法测定其对福寿螺胆碱酯酶活性的影响。结果表明,3种入侵植物的乙醇提取物对福寿螺AChE和BChE的活性均具有较强的抑制作用,薇甘菊和南美蟛蜞菊叶比茎的乙醇提取物抑制作用强,五爪金龙的茎则比叶的乙醇提取物抑制作用强。各提取物对胆碱酯酶的抑制作用存在浓度梯度依赖性特点,在质量浓度为0.100 0 mg/mL时的抑制率接近阳性对照组。五爪金龙的茎和薇甘菊的叶乙醇提取物对福寿螺两种胆碱酯酶的活性具有较强的抑制作用,抑制率可达70%以上,说明这两种入侵植物具有开发为福寿螺植物源杀螺剂的潜力。
关键词:入侵植物; 福寿螺(Pomacea canaliculata); 胆碱酯酶
中图分类号:S966.2;X176 文献标识码:A 文章编号:0439-8114(2016)06-1451-04
DOI:10.14088/j.cnki.issn0439-8114.2016.06.022
入侵物种已成为影响自然生态系统最严重的全球性问题,威胁到当地乃至全球的生态环境和经济发展[1]。菊科假泽兰属植物薇甘菊(Mikania micrantha)、菊科蟛蜞菊属植物南美蟛蜞菊(Wedelia trilobata)和旋花科番薯属植物五爪金龙(Ipomoea cairica)作为入侵植物已经对中国南方广大地区生态系统产生了严重的影响,它们生长迅速,环境适应性和抗逆性强,无性繁殖能力极强,不断入侵新的领地,同时产生化学物质,杀死其他植物,形成单种群落,破坏植被和原有的生態系统[2-4]。争夺本地植物的水分和养分而使其无法正常生长与繁殖,导致死亡。
福寿螺(Pomacea canaliculata)是近些年来备受国际关注的危害性外来入侵动物,软体动物门腹足纲前鳃亚纲腹足目瓶螺科瓶螺属,原产南美洲亚马逊河流域。福寿螺对环境适应性强、食性杂、个体大、生长迅速、繁殖力强,是危害生物多样性、农业生产和人类健康的世界性恶性入侵水生动物[5-8],2000年世界自然保护联盟外来入侵物种专家委员会将其列为世界100种恶性外来入侵物种之一。2003年3月国家环保总局将福寿螺列入首批入侵中国的16种“危害最大的外来物种”之一[9,10]。目前,中国对螺害的控制以化学杀螺剂防治为主,在一定程度为农业的发展发挥了较大作用,但是随着农业的发展,杀螺剂的使用越来越广泛,再加上不合理利用杀螺剂,造成了环境污染,给生物的生存带来了威胁[11]。因此寻找可持续开发的、环境友好型植物源杀螺剂,越来越受到研究者们的重视。
胆碱酯酶已成为杀虫剂开发研究的重要靶酶,主要存在于动物神经系统中的一种水解酶,它的作用是水解胆碱能神经末梢释放的神经介质乙酰胆碱。如果胆碱酯酶的作用被抑制,就会发生乙酰胆碱过剩而集聚的现象,引起胆碱能神经过度兴奋、类似有机磷中毒的症状[12,13]。胆碱酯酶抑制剂的使用能够抑制福寿螺胆碱酯酶的活性,突触后神经纤维长期处于兴奋状态,同时突触部位的正常神经冲动传导受阻塞,导致福寿螺中毒死亡,已成为螺害防治的重要途径。本试验通过研究入侵植物五爪金龙、南美蟛蜞菊和薇甘菊乙醇提取物对福寿螺乙酰胆碱酯酶(Acetylcholinesterase,AChE)和丁酰胆碱酯酶(Butyrylcholinesterase,BChE)活性的影响,以期获得以害治害的非化学防治福寿螺的有效途径,为入侵生物的防治工作提供参考。
1 材料与方法
1.1 动物材料
福寿螺成螺(9.56±2.30) g,采自广东省潮州市湘桥区社光村田间水沟,采集后洗净敲去壳,取斧足组织。
1.2 植物材料
薇甘菊、南美蟛蜞菊和五爪金龙均采集于韩山师范学院校内,经清洗后自然晾干,分离出茎和叶,剪成段,分别放入干燥箱,60 ℃干燥至恒重后磨碎,置于干燥处保存备用。
1.3 入侵植物乙醇提取物的制备
取每种磨碎的植物样品20 g,用95%乙醇100 mL浸泡,超声波提取30 min,过滤得到提取液后,放进60 ℃干燥箱蒸发浓缩成浸膏。称取适量浸膏,用去离子水配制成0.100 0、0.050 0、0.025 0、0.012 5 mg/mL的提取物溶液,4 ℃冰箱中保存备用。
1.4 酶液的制备
用手术刀将福寿螺的斧足组织切碎,磷酸缓冲盐溶液(Phosphate buffer saline,PBS)清洗后放入离心管中,按质量体积比1/10(g/mL)加入裂解缓冲液[10 mmol/L 4-羟乙基哌嗪乙磺酸、pH 7.5、1 mmol/L乙二胺四乙酸、1 mmol/L乙二醇双(2-氨基乙基醚)四乙酸、500 mmol/L氯化钠、0.5%曲拉通X-100、 10 μg/mL亮抑蛋白酶肽、20 μmol/L胃蛋白酶抑制剂、2.5 mmol/L盐酸苯甲脒]后,4 ℃匀浆,12 000 r/min 30 s,间隔30 s,重复2次,4 ℃ 1 600 r/min离心10 min,转移上清液至新的离心管中,12 000 r/min 4 ℃离心20 min,取上清液,-20 ℃冰箱中保存备用。
1.5 胆碱酯酶活性的测定及抑制率的计算
采用改进的Ellman等[14]方法在96孔酶标板上测定样品的AChE和BChE抑制活性。具体操作如下。
样品孔:在96孔酶标板中依次加入155 μL PBS(0.2 mol/L,pH 7.5),20 μL待测样品溶液,5 μL 酶液。4 ℃ 孵育20 min后,加10 μL 5,5'-二硫代双(2-硝基苯甲酸)[5,5'-Dithiobis-(2-nitrobenzoic acid),DTNB](12.5 mmol/L),室温静置30 min,加入10 μL底物溶液12.5 mmol/L碘化硫代乙酰胆碱(Acetylthiocholine iodide,ATCH)或碘化硫代丁酰胆碱(Butyrylthiocholine iodide,BTCH)。37 ℃孵育30 min后,用酶标仪在405 nm下测定其吸光度值。
样品本底对照孔:用5 μL PBS代替5 μL酶液,其他条件不变。
阴性对照孔:用20 μL PBS代替20 μL待测样品溶液,其他条件不变。
阳性对照孔:用20 μL 1-甲基-1,2,3,4-四氢异喹啉[1,5-Bis(4-Allyldimethylammoniumphenyl) pentan-3-one,Dibromide,BW284c51]或2 mmol/L四乙丙基焦磷酸亚胺(Tetraisopropyl pyrophosphoramide,iso-OMPA)代替20 μL待测样品溶液,其他条件不变。
抑制率计算公式为:抑制率=A阴性对照-(A样品-A样品本底)/A阴性对照×100%,试验数据用平均值±标准差(x±s)表示。
2 结果与分析
2.1 入侵植物乙醇提取物对福寿螺AChE活性的抑制效果
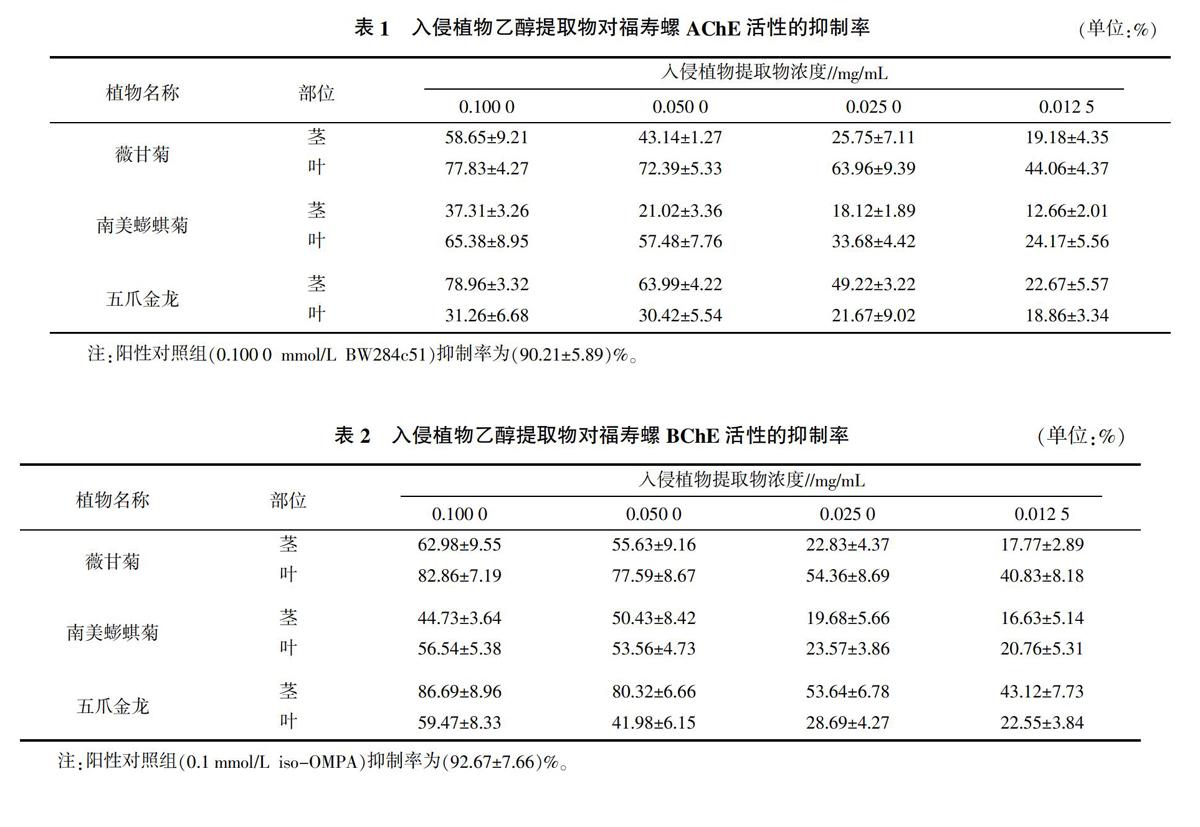
由表1可知,3种入侵植物乙醇提取物对福寿螺AChE活性均具有较强的抑制作用,但五爪金龙和薇甘菊比南美蟛蜞菊的抑制作用强。同种植物不同部位的乙醇提取物对福寿螺AChE的抑制强度也不同,薇甘菊和南美蟛蜞菊的叶乙醇提取物均比茎乙醇提取物的抑制作用强,五爪金龙茎乙醇提取物的抑制作用则比叶乙醇提取物强;在供试质量浓度为0.100 0 mg/mL时,五爪金龙的茎和薇甘菊的叶乙醇提取物对福寿螺AChE的抑制作用最强,抑制率分别为(78.96±3.32)%和(77.83±4.27)%,接近于阳性对照组的抑制率(90.21±5.89)%,其后依次为南美蟛蜞菊的叶和薇甘菊的茎乙醇提取物,抑制率分别为(65.38±8.95)%和(58.65±9.21)%,抑制率低于50%的为南美蟛蜞菊的茎和五爪金龙的叶乙醇提取物。同种植物同个部位的乙醇提取物对福寿螺AChE的抑制作用具有浓度依赖性特点,浓度越大抑制作用越强,随着浓度的降低,其抑制作用也减弱。
2.2 入侵植物乙醇提取物对福寿螺BChE活性的抑制效果
由表2可以看出,3种入侵植物乙醇提取物对福寿螺BChE活性均具有较强的抑制作用,五爪金龙和薇甘菊比南美蟛蜞菊的抑制作用强。同种植物不同部位的乙醇提取物对福寿螺BChE的抑制强度也不同,薇甘菊和南美蟛蜞菊的叶乙醇提取物均比茎乙醇提取物的抑制作用强,五爪金龙茎乙醇提取物的抑制作用比叶乙醇提取物强;在质量浓度为0.100 0 mg/mL时,五爪金龙的茎和薇甘菊的叶乙醇提取物对福寿螺BChE的抑制作用最强,抑制率分别为(86.69±8.96)%和(82.86±7.19)%,接近阳性对照组的抑制率(92.67±7.66)%,其后依次为薇甘菊的茎、五爪金龙的叶和南美蟛蜞菊的叶乙醇提取物,抑制率分别为(65.38±8.95)%、(59.47±8.33)%和(58.65±9.21)%,南美蟛蜞菊的茎乙醇提取物的抑制作用最小。同种植物同个部位的乙醇提取物对福寿螺BChE的抑制作用具有浓度依赖性特点,即浓度越大抑制作用越强,随着浓度的降低,其抑制作用也减弱。
3 小结与讨论
福寿螺的泛滥成灾给农业生产和人类健康造成了极大的危害。目前对福寿螺的防治方法主要包括人工防治、化学防治、生态防治和生物防治。化学防治虽然是福寿螺防治的有效方法之一,但使用化学农药存在杀伤非靶标生物的问题,而且也不排除福寿螺抗药性的产生,最终出现杀螺效果差而环境破坏大的严重恶果[15]。因此,寻找与环境友好的高效低毒的新型殺螺剂势在必行。目前,植物源杀螺剂的研究成为生物防治的研究热点之一,也是生物杀螺技术研究的重要内容。迄今为止,大约有1 500种植物被用于杀螺活性试验,并已发现不少植物具有杀螺活性[16-18]。而利用入侵植物资源对福寿螺进行治理,在有效控制目标生物的同时,泛滥入侵的植物也将得到有效治理,这样就能达到一举两得的效果。
本试验结果表明,薇甘菊和五爪金龙两种入侵植物乙醇提取液对福寿螺两种胆碱酯酶都具有较强的抑制作用。在质量浓度为0.100 0 mg/mL和0.050 0 mg/mL时,五爪金龙的茎乙醇提取液对福寿螺AChE具有较高的抑制作用,抑制率达(78.96±3.32)%和(63.99±4.22)%,对BChE的抑制率达(86.69±8.96)%和(80.32±6.66)%,接近阳性对照组的抑制率。五爪金龙乙醇提取物具有直接杀螺作用,并且五爪金龙乙醇提取液对水稻苗的生长没有不利的影响[19,20],因此,利用五爪金龙来防治福寿螺是一项环境友好型的非化学防治措施,具有良好的应用前景。薇甘菊的叶乙醇提取物对福寿螺AChE和BChE也具有较高的抑制效果,在质量浓度为0.100 0 mg/mL时,抑制率分别为(77.83±4.27)%和(82.86±7.19)%,也非常接近阳性对照组的抑制率。薇甘菊是一种检疫性杂草,严重影响生物多样性及国土生态安全[21-23]。利用薇甘菊来防治福寿螺,基本上能达到以害治害的目的,具有较好的开发福寿螺植物源杀螺剂的潜力。南美蟛蜞菊茎和叶乙醇提取物对福寿螺AChE和BChE的抑制效果相对较弱,在0.100 0 mg/mL的高浓度时,对福寿螺AChE和BChE的抑制率分别为(65.38±8.95)%和(56.54±5.38)%。
综上所述,通过利用入侵植物乙醇提取物对福寿螺胆碱酯酶活性的抑制作用研究,表明五爪金龙和薇甘菊这两种入侵植物在福寿螺防治方面具有开发潜力。本试验对入侵植物进行部位处理,但具体是植物中的哪些成分起抑制作用还需要进一步研究。对这些入侵植物进行进一步系统深入的生物活性成分研究,包括对其进行活性追踪、活性成分分离与鉴定,从而研制出高效的植物抑螺剂,对福寿螺的防治具有重要的意义。
参考文献:
[1] 向言词,彭少麟,周厚诚,等.生物入侵及其影响[J].生态科学,2001,20(4):68-72.
[2] 廖周瑜,赵则海,侯玉平,等.五爪金龙对薇甘菊的化感效应研究[J].生态环境,2007,16(3):939-943.
[3] 宋莉英,彭长连,彭少麟.华南地区3种入侵植物与本地植物叶片建成成本的比较[J].生物多样性,2009,17(4):378-384.
[4] 吴彦琼,胡玉佳.外来植物南美蟛蜞菊、裂叶牵牛和五爪金龙的光合特性[J].生态学报,2004,24(10):2334-2339.
[5] JOSHI R. Managing invasive alien molluse species in rice[J].International Rice Research Notes,2005,30(2):5-12.
[6] COWIE R H. Apple Snails(Ampullariidae) as Agricultural pests: Their biology, Impacts and Management. In:Baker GM (ed) Molluscs as Crop Pests[M].Wallingford:CABI Publishing,2002.
[7] 郭 靖,章家恩.福寿螺入侵机制的研究进展[J].生态学杂志,2014,33(3):806-815.
[8] CASTRO-VAZQUEZ A,ALBRECHT E A,VEGA I A,et al. Pigmented corpuscles in the midgut gland of Pomacea canaliculata and other Neotropical apple-snails (Prosobranchia, Ampullariidae):A possible symbiotic association[J]. Biocell,2002, 26(1):101-109.
[9] 章家恩,方 丽.关于我国农田福寿螺生物入侵需要加以研究的生态学问题[J].中国生态农业学报,2008,16(6):1585-1589.
[10] ZOU X H,XIE H Q H,ZHA G C,et al. Characterizations of cholinesterases in golden apple snail (Pomacea canaliculata)[J].Journal of Molecular Neuroscience,2014,53(3):424-428.
[11] 王志高,谭济才,刘 军,等.福寿螺综合防治研究进展[J].中国农学通报,2009,25(12):201-205.
[12] DOMINGUES I,AGRA A R,MONAGHAN K A,et al. Cholinesterase and glutathione-S-transferase activities in freshwater invertebrates as biomarkers to assess pesticide contamination[J].Environ Toxicol Chem,2010,29(1):5-18.
[13] MIAO Y Q,HE N Y,ZHU J J. History and new developments of assays for cholinesterase activity and inhibition[J].Chem Rev,2010,110(9):5216-5234.
[14] ELLMAN G L,COURTNEY K D,ANDRES V, et al. A new and rapid colorimetric determination of acetylcholinesterase activity[J].Biochemical Pharmacology,1961,7(2):88-95.
[15] 陈晓娟,何忠全,高 平.10种植物提取物对稻田福寿螺的毒杀活性[J].西南农业学报,2012,25(4):1294-1297.
[16] 閆小红,周 兵,蒋 平,等.16种植物提取物对福寿螺杀螺效果的研究[J].广东农业科学,2011,38(4):81-84.
[17] BAPTISTA D F,VASCONCELLOS M C,LOPES F E,et al. Perspectives of using Euphorbia splendens as a molluscicide in schistosomiasis control programs[J].Southeast Asian Journal of Tropical Public Health,1994,25(3):419-424.
[18] ALARD F,GEERTS S,TRIEST L,et al. Toxicity of the molluscicidal plant, Ambrosia maritima L., to aquatic non-target organisms[J].Toxicon,1991,29(6):745-750.
[19] 胡 飞,曾坤玉,张俊彦,等.五爪金龙乙醇提取物对福寿螺毒杀和水稻苗生长的影响[J].生态学报,2009,29(10):5471-5477.
[20] 曾坤玉,胡 飞,陈玉芬,等.四种与福寿螺(Ampullaria gigas)同源地入侵植物的杀螺效果[J].生态学报,2008,28(1):260-266.
[21] 王瑞龙,陈 颖,张 晖,等.薇甘菊萎蔫病毒寄主范围、传播媒介和危害特点[J].生态学杂志,2013,32(1):72-77.
[22] 全国明,章家恩,谢俊芳,等.薇甘菊入侵对中小型土壤动物群落结构特征的影响[J].应用生态学报,2011,22(7):1863-1870.
[23] 李鸣光,鲁尔贝,郭 强,等.入侵种薇甘菊防治措施及策略评估[J].生态学报,2012,32(10):3240-3251.
