近30年太湖沉水植物优势种遥感监测及变化分析
2016-10-19周兴东罗菊花
王 琪, 周兴东, 罗菊花, 陈 冲
(1.江苏师范大学城建与环境学部,江苏 徐州 221000; 2.中国科学院南京地理与湖泊研究所,江苏 南京 210008)
近30年太湖沉水植物优势种遥感监测及变化分析
王琪1, 周兴东1, 罗菊花2, 陈冲1
(1.江苏师范大学城建与环境学部,江苏 徐州221000; 2.中国科学院南京地理与湖泊研究所,江苏 南京210008)
利用1985—1986年、1995—1996年、2004—2006年和2014—2015年多时相遥感影像数据,构建了水生植物分类决策树模型,结合沉水植物的生活史差异,研究太湖近30年沉水植物优势种的演替模式。4个时期的沉水植物遥感分类总体精度与Kappa系数依次为64.86%和0.59,70.83%和0.65,63.93%和0.50,64.94%和0.60。研究表明:1985—2015年马来眼子菜由东山湾向西山岛、东太湖向南太湖扩张,竺山湾、梅梁湾的苦草呈锐减的趋势,近年来梅梁湾的沉水植物只有菹草1种,湖心区一直未监测到沉水植物,东太湖沉水植物分布面积最广、种类最多。近30年太湖沉水植物演替主要经历4个阶段,依次为苦草-眼子菜型、小眼子菜型、金鱼藻-伊乐藻型和眼子菜-伊乐藻型阶段。水体富营养化和围网养殖是沉水植物演替的主要原因。
沉水植物;遥感;优势种;太湖
改革开放以来,人类经济活动剧烈,而相应的环境保护措施不够及时,水体富营养化进程加剧,许多湖泊藻华频发,严重影响周边城市的供水安全。沉水植物作为大型草本植物,是湖泊生态系统营养循环的核心环节,对湖泊水质的改变和湖泊生态系统由“浊水态”向“清水态”转化具有重要的作用[1]。研究沉水植物多年的空间分布和面积变化对于改善湖泊生态环境具有重要的指导意义。
针对我国许多湖泊水生植被的变化已经开展了一系列的工作,如葛继稳等[2]通过对梁子湖主体湖1955—2001年以来45年间的水生植被演替的研究,为梁子湖湿地生态系统的恢复与重建提供科学依据;刘伟龙等[3]通过对洪泽湖水生植被过去50年的变化特征的研究表明,人类活动是水生植物种群变化的主要原因;郝孟曦等[4]通过对湖北长湖水生植物多样性及群落演替的研究表明,水体富营养化及大规模围网养殖是长湖水生植物多样性显著下降和群落发生演替的主要原因。目前对于水生植物的研究局限于传统的人工调查法,遥感技术能够快速获取大面积连续地物信息,故在水生植物时空分布信息监测方面具有传统方法不可比拟的优势。近年来国内外很多学者试图利用中等分辨率遥感数据监测水生植物的时空分布,并取得了一些成果,如Ma等[5]在太湖水生植物的遥感提取中,将水体透明度和水深的先验知识引入分类决策树,较准确地提取了太湖的水生植物。林川等[6]等基于中等分辨率TM数据提取湿地水生植被。对于沉水植物优势种遥感识别研究,高光谱或超光谱遥感可以区分不同种类沉水植物,如Hestir等[7]利用Hymap遥感影像对美国加州三角洲的水生植物进行了物种识别;Wang等[8]利用航空高光谱遥感数据对威尼斯一处水域的混合水生植物群落的分类进行了研究。然而高光谱和超光谱遥感影像的获取较为困难,不利于沉水植物优势种历史变迁研究。
本文以富营养严重的太湖为研究区,利用多时相中等分辨率的Landsat TM5/HJ-CCD影像,构建了水生植物分类决策树模型,结合不同沉水植物的生活史信息,对太湖大型沉水植物近30年的空间分布及消长规律进行研究,将为太湖沉水植物资源的合理开发利用和延缓湖泊沼泽化提供重要依据。
1 数据和方法
1.1研究区概况
太湖是我国的第三大淡水湖泊,位于30°55′40″N~31°32′58″N,119°52′32″E~120°36′10″E,中国科学院太湖湖泊生态系统研究站1989年以来监测数据显示,太湖的沉水植被主要分布于太湖东部、南部及北部,湖心区基本无水生植物分布。为了便于本研究对沉水植物监测结果定性描述和定量分析,参考已有的资料和文献[9],将太湖划分为8大湖区(图1),分别包括Ⅰ区湖北两湾(竺山湾、梅梁湾),Ⅱ区贡山湾,Ⅲ区(镇湖湾、光福湾),Ⅳ区(胥口湾、东山湾),Ⅴ区环西山岛附近水域,Ⅵ区东太湖,Ⅶ区南太湖和Ⅷ区湖心区。先后于2014年和2015年夏季对太湖开展2次野外样点采集,样点采集包括水生植物优势种的类型、盖度、透明度和GPS等,样点总数为174个。并结合以往太湖水生植被的样点调查结果[9-11]。
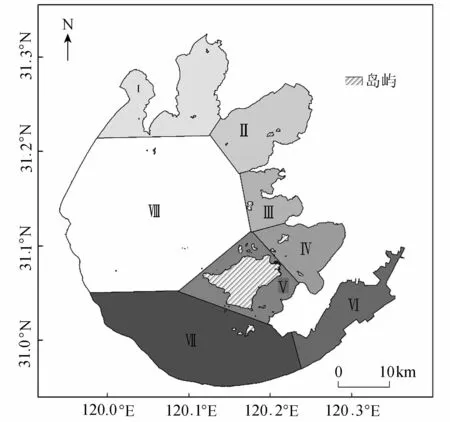
图1 太湖湖区划分
1.2数据收集与预处理
本研究下载了多时相的Landsat TM5/HJ-CCD影像共包括1985—1986年5景,1995—1996年5景、2004—2006年7景及2014—2015年12景太湖流域的遥感影像,各期影像基本无云覆盖,有关影像的具体信息见表1。通过ENVI软件对遥感影像进行辐射校正(辐射定标和大气校正),消除一切与辐射有关的误差。此外,利用精校正过的覆盖太湖的Landsat TM 8影像对所有影像开展几何校正,其校正误差控制在0.5个像元之内。
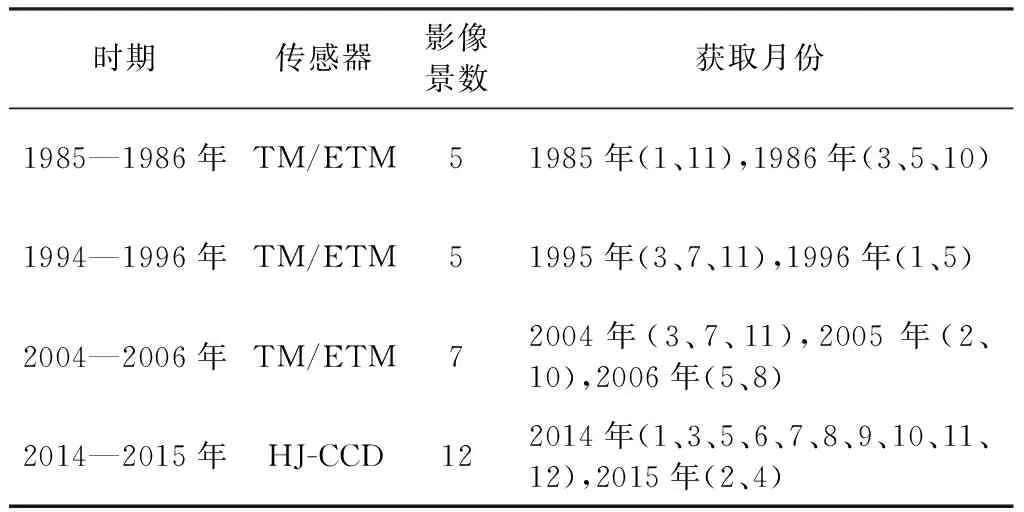
表1 研究区卫星遥感影像信息
1.3水生植物决策树分类模型构建
本研究基于TM5/HJ-CCD影像的蓝、绿、红和近红外4个光谱波段,通过波段组合和影像变换技术(主成分变换和缨帽变换)构建了3个光谱特征变量。大量的实验研究表明,第4波段和第1 、2 、3 波段之和的差值与4个波段之和的比值可以较好地区分挺水植物;主成分变换后的第二主成分能够较好地识别浮叶植物;缨帽变换后的亮度指数和绿度指数的差值对沉水植物具有很好的识别潜力。构建不同水生植物的分类特征指数,emergent vegetation index (EVI)用于分类挺水植物,floating-leaved vegetation index (FVI)用于分类浮叶植物,submerged vegetation index (SVI)用于分类沉水植物。在遥感分类中,决策树是制定分类规则的有效方法之一,构建决策树的关键是确定叶结点测试变量和分割阈值,本文采用Breiman等[12]提出的一种决策树构建算法—classification and regression tree (CART)算法。通过随机抽取样本的方式确定基尼系数的大小,基于该算法,本文构建了水生植物分类决策树见图2。

注:a,b,c为阈值。图2 基于CART算法构建的决策树

年份沉水植物分类方法1985年伊乐藻和菹草可以在0℃以下继续生长繁殖,1985年尚未引进伊乐藻,所以1985年1月份的沉水植物为菹草。微齿眼子菜在4-5月份大量萌生,1986年3月份沉水植物图层(微齿眼子菜较少)擦除1986年5月份沉水植物图层(微齿眼子菜较多)得到微齿眼子菜,马来眼子菜在10月下旬达到最大生物量,1985年11月份的沉水植物图层为马来眼子菜,合并总的沉水植物,剩余未分类的沉水植物为苦草1995年7月份菹草的生物量达到最小值,而此时伊乐藻的生物量达到最大值,1995年7月份的沉水植物图层(伊乐藻较多)擦除1996年1月份的沉水植物图层(菹草和伊乐藻)得到菹草图层,菹草图层擦除1996年1月份沉水植物图层得到伊乐藻图层,马来眼子菜、苦草、微齿眼子菜的分类方法同上2005年金鱼藻在8月下旬到9月份初达到最大生物量,随后很快消亡,2005年10月份沉水植物图层(金鱼藻较少)擦除2006年8月份沉水植物图层(金鱼藻最多)得到金鱼藻,马来眼子菜、苦草、微齿眼子菜、菹草、伊乐藻的分类方法同上2015年黑藻在5、6月份生长繁殖较快,2014年4月份沉水植物图层(黑藻较少)擦除2014年6月份沉水植物图层(黑藻较多)得到黑藻,在7月份狐尾藻达到生物量最大值,10月份其生物量相对最低,2014年10月份的沉水植物图层(狐尾藻较少)擦除2014年7月份沉水植物图层(狐尾藻最多)得到狐尾藻,马来眼子菜、苦草、微齿眼子菜、菹草、伊乐藻的分类方法同上
1.4沉水植物优势种分类方法
多样性分析表明,水生植物种类在4年内未发生明显变化[9],笔者认为在1985—1986年、1995—1996年、2004—2006年和2014—2015年间的太湖沉水植物并没有发生大范围的空间变化,借助GIS技术提取了4个时期29个月份遥感影像中的沉水植物,参照考虑生活史的太湖沉水植物优势种遥感监测方法[13],多次使用Arcgis空间叠加分析中的擦除功能,具体分类方法见表2。
2 结果与分析
2.1太湖沉水植物优势种的空间分布特征
执行上述分类操作流程,后提取的沉水植物需擦除先提取的沉水植物,避免重合交叉,最终合并成总的沉水植物图层,1985、1995、2005、2015年太湖沉水植物优势种空间分布结果见图3,太湖沉水植物主要分布于太湖东部,沉水植物总的分布面积呈线性增加(图4)。
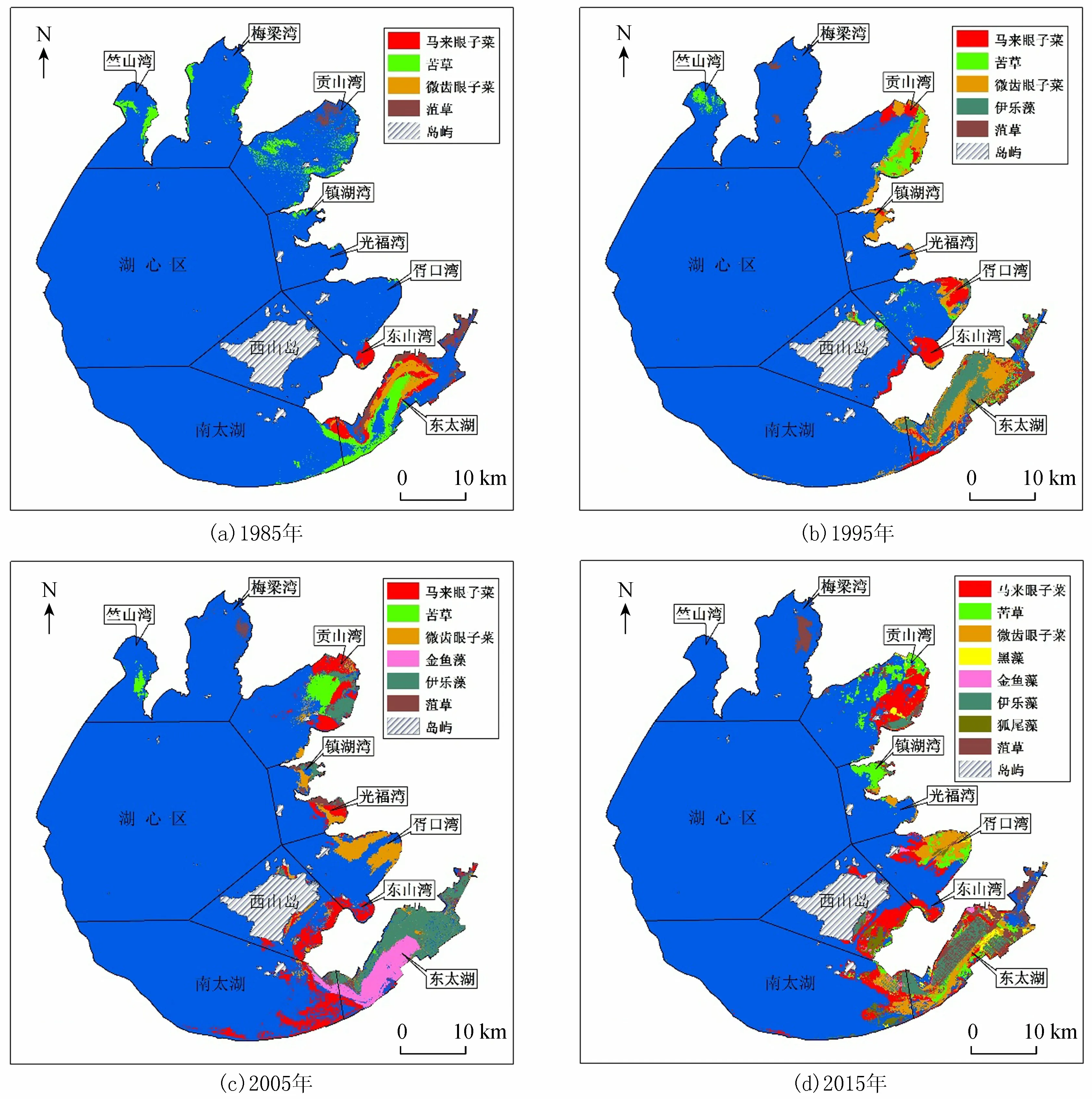
图3 1985—2015年太湖沉水植物优势种空间分布

图4 1985—2015年太湖沉水植物总的分布面积
1985年太湖沉水植物优势种共4种(图5(a)),苦草是太湖的主要优势种,主要分布于Ⅵ区、Ⅱ区、Ⅰ区和Ⅶ区。马来眼子菜为次优种,主要分布于Ⅵ区、Ⅳ区和Ⅶ区,大部分沉水植物集中于Ⅵ区,常构成苦草+马来眼子菜+微齿眼子菜+菹草群从。各个湖区的沉水植物面积总和的大小依次为Ⅵ>Ⅱ>Ⅶ>Ⅰ>Ⅲ>Ⅳ>Ⅴ> Ⅷ;1995年太湖沉水植物优势种共5种(图5(b)),微齿眼子菜的分布最为广泛,主要分布于Ⅵ区、Ⅱ区和Ⅳ区,常在Ⅵ区构成微齿眼子菜+伊乐藻群从,Ⅱ区构成微齿眼子菜+苦草+马来眼子菜群从,伊乐藻为次优种,主要分布在东太湖围网养殖区,菹草在梅梁湾已经有少量分布,各个湖区的沉水植物面积总和的大小依次为Ⅵ>Ⅱ>Ⅳ>Ⅲ>Ⅰ>Ⅴ>Ⅶ> Ⅷ;2005年太湖沉水植物优势种共6种(图5(c)),马来眼子菜主要分布于Ⅴ区、Ⅳ区和Ⅵ区,在Ⅳ区(东山湾)呈单优种分布。伊乐藻和金鱼藻是Ⅵ区的主要优势种,常构成伊乐藻+金鱼藻群从。微齿眼子菜主要呈单优种分布于胥口湾,各个湖区的沉水植物面积总和的大小依次为Ⅵ>Ⅱ>Ⅶ>Ⅳ>Ⅴ>Ⅲ>Ⅰ> Ⅷ;2015年太湖沉水植物优势种共8种(图5(d)),马来眼子菜主要分布于Ⅱ区、Ⅴ区、Ⅳ区和Ⅵ区。微齿眼子菜主要分布于Ⅳ区(胥口湾),苦草主要分布于Ⅱ区和Ⅲ区,菹草主要呈单优种分布于Ⅰ区(梅梁湾),伊乐藻为Ⅵ区的主要优势种,黑藻和狐尾藻主要分布于Ⅶ区,多为伴生种,各个湖区的沉水植物面积总和的大小依次为Ⅵ>Ⅱ>Ⅳ>Ⅶ>Ⅴ>Ⅲ>Ⅰ> Ⅷ。
2.2水生植被变化分析
借助Arcgis的空间叠加分析模块,本文对1985—1995年、1995—2005年和2005—2015年太湖各类沉水植物动态变化进行了统计和分析,结果见图6,并计算它们的转移矩阵,如表3~5所示,各类沉水植物和水体之间互有转换,具体变化特征如下。
1985—1995年苦草净减少53.58 km2,大部分转为水体和微齿眼子菜;1995—2005年苦草净减少17.94 km2,大部分转为水体和伊乐藻;2005—2015年苦草净增加37.36 km2,其转入大部分来自水体和微齿眼子菜,主要集中在Ⅵ区。随着Ⅰ区水域富营养化的加剧,近年来苦草已经消亡。30年间马来眼子菜的扩张最为显著,主要集中在Ⅳ区、Ⅴ区和Ⅶ区,1985年马来眼子菜已经在Ⅳ区(东山湾)呈单优种建群,1985—1995年净增加28.86 km2,1995—2005年净增加49.89 km2,2005—2015年净增加6.67 km2,其转入大部分来自于水体。这主要是因为马来眼子菜适应性强,能够分布于水较深、又有风浪的区域,一旦形成单一化的植物群落将无法为其他水生生物提供广泛适宜的生存条件,造成水生生物多样性的下降,湖泊生态平衡受到严重影响,近年来在Ⅴ区水域,马来眼子菜逐年扩张,成为该区域的先锋种。1985—1995年微齿眼子菜净增加72.25 km2,其转入大部分来自水体和苦草,1995—2005年微齿眼子菜净减少37.50 km2,其大部分转出为伊乐藻和金鱼藻,2005—2015年微齿眼子菜净增加1.56 km2,其转入和转出基本持平,主要变化区域为Ⅳ区和Ⅵ区。1985—1995年伊乐藻净增加48.58 km2,其转入大部分来自于微齿眼子菜和马来眼子菜,1995—2005年伊乐藻净增加43.32 km2,其转入大部分来自于微齿眼子菜,2005—2015年伊乐藻净增加2.51 km2,其转入和转入基本持平,东太湖围网养殖基本饱和,伊乐藻已经在Ⅵ区稳定建群。1985—1995年菹草净减少18.11 km2,其转出最多为微齿眼子菜,1995—2005年菹草净增加8.02 km2,其转入大部分来自于水体,2005—2015年菹草净增加15.8 km2,菹草主要分布在Ⅰ区,主要是由于梅梁湾3面环山,为相对静水的环境,同时菹草的生长季节避开了藻华爆发的高峰期。1995—2005年金鱼藻净增加49.11 km2,其转入大部分来自于微齿眼子菜和水体,2005—2015年金鱼藻净减少43.56 km2,其大部分转为伊乐藻,金鱼藻为无根类沉水植物,在Ⅶ区呈单优种分布。微齿眼子菜一旦被大量破坏,其生物量剧增。狐尾藻和黑藻对水质适用性强,对底质要求不严,在大湖的分布面广,主要为伴生种。

注:多年来Ⅷ区未监测到沉水植物图5 各年太湖沉水植物优势种各分区面积

图6 太湖主要沉水植物动态变化
表31985—1995年太湖沉水植物类型转移矩阵 km2
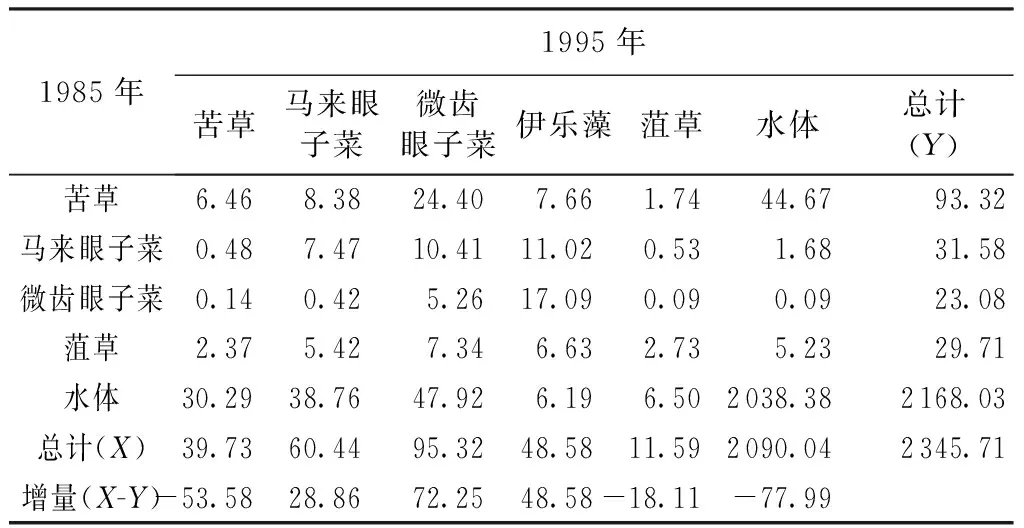
1985年1995年苦草马来眼子菜微齿眼子菜伊乐藻菹草水体总计(Y)苦草6.468.3824.407.661.7444.6793.32马来眼子菜0.487.4710.4111.020.531.6831.58微齿眼子菜0.140.425.2617.090.090.0923.08菹草2.375.427.346.632.735.2329.71水体30.2938.7647.926.196.502038.382168.03总计(X)39.7360.4495.3248.5811.592090.042345.71增量(X-Y)-53.5828.8672.2548.58-18.11-77.99
注:X为1985—1995年面积转入量;Y为1985—1995年面积转出量;X-Y为1985—1995年面积净增加量。
表41995—2005年太湖沉水植物类型转移矩阵km2
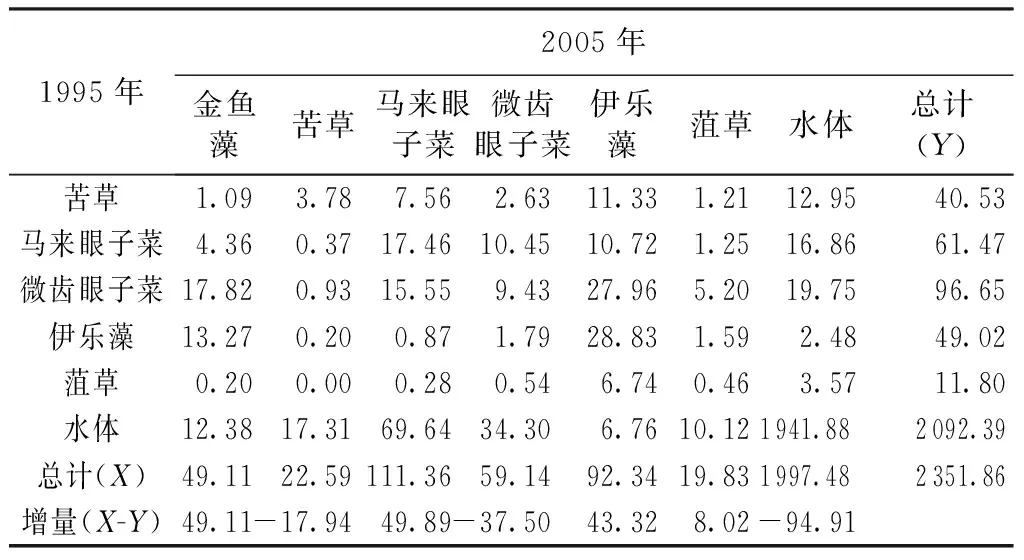
1995年2005年金鱼藻苦草马来眼子菜微齿眼子菜伊乐藻菹草水体总计(Y)苦草1.093.787.562.6311.331.2112.9540.53马来眼子菜4.360.3717.4610.4510.721.2516.8661.47微齿眼子菜17.820.9315.559.4327.965.2019.7596.65伊乐藻13.270.200.871.7928.831.592.4849.02菹草0.200.000.280.546.740.463.5711.80水体12.3817.3169.6434.306.7610.121941.882092.39总计(X)49.1122.59111.3659.1492.3419.831997.482351.86增量(X-Y)49.11-17.9449.89-37.5043.328.02-94.91
注:X为1995—2005年面积转入量;Y为1995—2005年面积转出量;X-Y为1995—2005年面积净增加量。
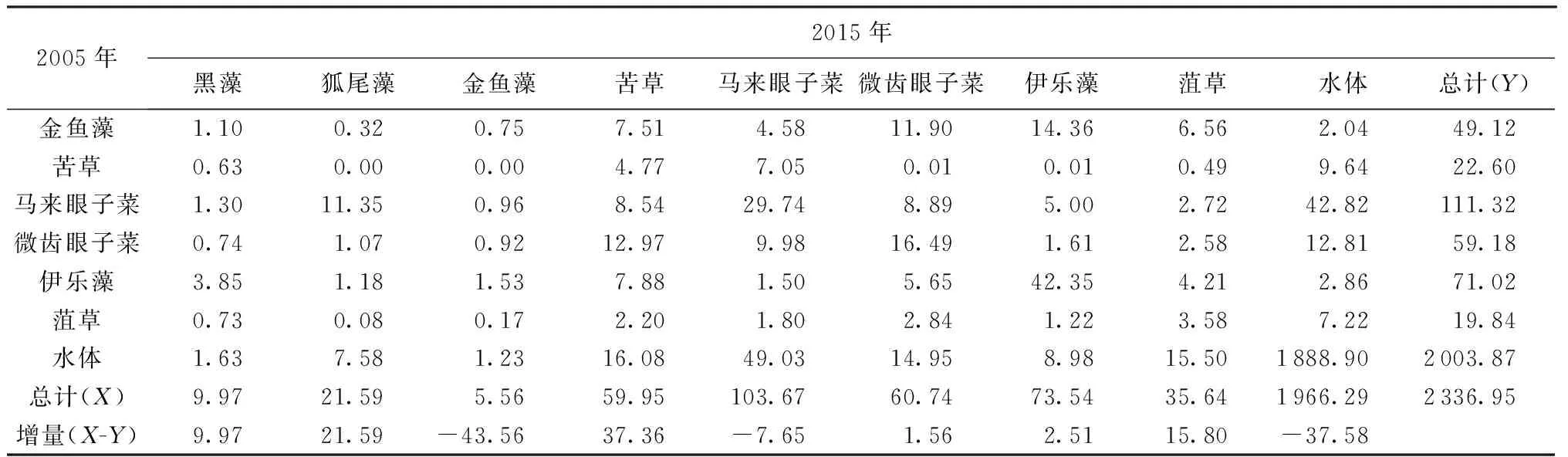
表5 2005—2015年太湖沉水植物类型转移矩阵 km2
注:X为2005—2015年面积转入量;Y为2005—2015年面积转出量;X-Y为2005—2015年面积净增加量。
2.3太湖沉水植物优势种遥感分类精度
通过2014、2015年野外调查样点及以往的太湖水生植被调查结果对沉水植物优势种识别的结果进行验证,具体情况见表6,1985、1995、2005、2015年太湖沉水植物遥感分类总体精度和Kappa系数分别为64.86%和0.59、70.83%和0.65、63.93%和0.50、64.94%和0.60。由于伊乐藻大部分分布在东太湖的网围养殖区,水生植物太过茂密而船只无法通过,采集样点极少,夏季是菹草的休眠期,也未在同期获得验证样点,所以两者未开展精度验证。
3 结 论
本文采用较长时间序列的遥感数据,分不同时期,研究了近30年太湖沉水植物优势种的变化,得到了以下结论:
a. 近30年来,东太湖的沉水植物面积增加,马来眼子菜由东山湾向西山岛、东太湖向南太湖水域扩张,伊乐藻逐渐占据整个太湖围网养殖区,胥口湾滋生大量微齿眼子菜,多年来湖心区未监测到沉水植物,竺山湾和梅梁湾的苦草呈锐减的趋势,近年来梅梁湾除有菹草分布外,苦草已基本消亡。
b. 太湖沉水植被演替主要包含4个阶段:苦草-眼子菜型阶段,小眼子菜型阶段,金鱼藻-伊乐藻型阶段和眼子菜-伊乐藻型阶段。1985年太湖围网养殖刚刚开始,水体营养水平较低,沉水植被以马来眼子菜、微齿眼子菜、菹草等眼子菜属植物为主,由于水质尚好,苦草的生物量和分布面积也较大。到了1995年,东太湖围网养殖规模迅速增大,东太湖在人为干预状态下,沉水植物次生演替的结果是适宜静水环境和耐污染的微齿眼子菜大量生长。2000年之后,围网养殖区域突破东太湖,向西太湖发展,到了2005年,七都以北、西山岛以南、东太湖以东的区域都出现大面积的围网养殖。伊乐藻自1986年引进太湖以来,开始逐渐入侵当地的苦草+马来眼子菜+微齿眼子菜群从,其分布面积呈逐年增加的趋势,是东太湖的主要优势种。近年来太湖藻华频发,水体透明度严重下降,西太湖沉水植物完全消失,太湖沉水植被优势种主要以耐污性较强的眼子菜类为主,遥感监测的结果显示菹草在梅梁湾呈单优种分布,在太湖东部未被污染的几个湖区仍然残留大量的马来眼子菜和微齿眼子菜。伊乐藻逐渐占据整个太湖养殖区。
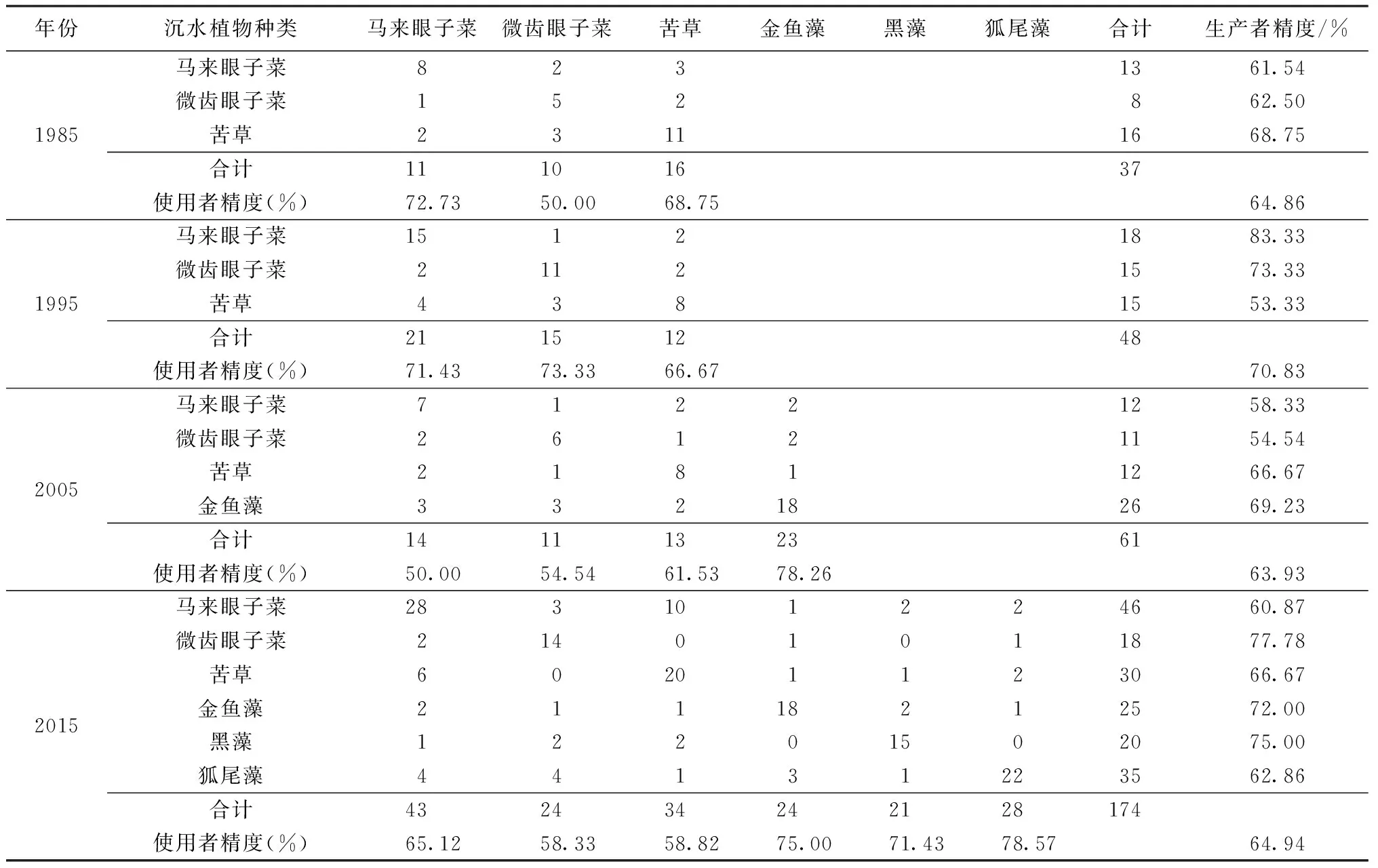
表6 4年沉水植物优势种分类后识别混淆矩阵
c. 重建以沉水植物为主的水生植物对于西太湖的富营养化的恢复至关重要,但绝不能盲目,湖泊恢复必须将工程措施和生态调控措施结合起来,根据湖泊水生植被自身演替规律和水生植物的生理生态特征,在早期可以选择耐污性强的沉水植物作为先锋种类,逐渐对水生植被的结构加以优化。
[ 1 ] 金相灿,刘树坤,章宗涉.中国湖泊环境[M].北京:海洋出版社,1995:234-302.
[ 2 ] 葛继稳,蔡庆华,李建军,等.梁子湖水生植被1955—2001年间的演替[J].北京林业大学学报,2004,26(1):14-20.(GE Jiwen,CAI Qinghua,LI Jianjun,et al.On aquatic vegetation succession of Lake Liangzihu from 1955 to 2001[J].Journal of Beijing Forestry University,2004,26(1):14-20.(in Chinese))
[ 3 ] 刘伟龙,邓伟,王根绪,等.洪泽湖水生植被现状及过去50多年的变化特征研究[J].水生态学杂志,2009,2(6):1-7.(LIU Weilong,DENG Wei,WANG Genxu,et al.Aquatic macrophyte status and variation characteristics in the past 50 years in Hongzehu Lake[J].Journal of Hydroecology,2009,2(6):1-7.(in Chinese))
[ 4 ] 郝孟曦,杨磊,孔祥虹,等.湖北长湖水生植物多样性及群落演替[J].湖泊科学,2005,27(1):94-102.(HAO mengxi,YANG Lei,KONG Xianghong,et al.Diversity and community succession of macrophytes in Lake Changhu,Hubei Province[J].Journal of Lake Sciences,2005,27(1):94-102.(in Chinese))
[ 5 ] MA R H,DUAN H T,GU X H,et al.Detecting aquatic vegetation changes in Taihu Lake[J].China Using Multi-Temporal Satellite Imagery Sensors,2008(8): 3988-4005.
[ 6 ] 林川,宫兆宁,赵文吉.基于中等分辨率TM数据的湿地水生植被提取[J].生态学报,2010,30(23):6460-6469.(LIN Chuan,GONG Zhaoning,ZHAO Wenji,The extraction of wetland hydrophytes types based on medium resolution TM data[J].Acta Ecologica Sinca,2010,30(23): 6460-6469.(in Chinese))
[ 7 ] HESTIR E L,KHANNA S,ANDREW M E,et al.Identification of invasive vegetation using hyperspectral remote sensing in the California Delta ecosystem[J].Remote Sensing of Environment,2008,112 (11):4034-4047.
[ 8 ] WANG C,MENENTI M,STOLL M P,et al.Mapping mixed vegetation communities in salt marshes using airborne spectral data[J].Remote Sensing of Environment,2007,107(4):559-570.
[ 9 ] 刘伟龙,胡维平,陈永根,等.西太湖水生植物时空变化[J].生态学报,2007,27(1):1-12.(LIU Weilong,HU Weiping,CHEN Yonggen,et al.Temporal and spatial variation of aquatic macrophytes in west Taihu Lake[J].Acta Ecologica Sinca,2007,27(1):1-12.(in Chinese))
[10] 鲍建平,缪为民,李劫夫,等.太湖水生维管束植物及其合理开发利用的调查研究[J].大连水产学院学报,1991,6(1):13-20.(BAO Jianpin,MIAO weimin,LI Jiefu,et al.Preliminary stuy on the auqatic weeds of the Tai Lake and their rational development and utilization[J].Journal of Dalian Fisheries College,1991,6(1):13-20.(in Chinese))
[11] 雷泽湘,徐德兰,黄沛生,等.太湖沉水和浮叶植被及其水环境效用研究[J].生态环境,2006,15(2):239-243.(LEI Zexiang,XU delan,HUANG Peisheng et al.Submersed and floating-leaved macrophytes in Taihu Lake and their water environmental effect[J].Ecology and Environment,2006,15(2):239-243.(in Chinese))
[12] BREIMAN L,FRIEDMAN J H,OLSHEN R A,et al.Classification and regression tree[M].Monterey: U.S.A.Wadsworth International Group,1984:1-358.
[13] 王琪,周兴东,罗菊花,等.考虑生活史的太湖沉水植物优势种遥感监测[J].湖泊科学,2015,27(5):953-961.(WANG Qi,ZHOU Xingdong,LUO Juhua,et al.The remote sensed monitoring of the dominant submerged vegetation in lake Taihu considering life history[J].Journal of Lake Sciences,2015,27(5):953-961.(in Chinese))
Remote sensing monitoring and analysis of dominant species of submerged vegetation in Taihu Lake over last 30 years
WANG Qi1, ZHOU Xingdong1, LUO Juhua2, CHEN Chong1
(1.DepartmentofUrbanConstructionandEnvironment,JiangsuNormalUniversity,Xuzhou221000,China;2.NanjingInstituteofGeographyandLimnology,ChineseAcademyofSciences,Nanjing210008,China)
Multi-temporal remote sensing image data over the periods from 1985 to 1986, from 1995 to 1996, from 2004 to 2006, and from 2014 to 2015 were used to construct a classification tree (CT) model for aquatic vegetation. With consideration of the differences in phenological information of submerged vegetation in Taihu Lake, the succession of the dominant species of submerged vegetation in the lake over the last 30 years was studied. The overall accuracies and Kappa coefficients of the remote sensing classification results for the four periods were 64.86% and 0.59, 70.83% and 0.65, 63.93% and 0.50, and 64.94% and 0.60, respectively. The results show thatPotamogetonmalaianushas expanded from Dongshan Bay to Xishan Island, and from East Taihu Lake to South Taihu Lake from 1985 to 2015. There was a declining trend in the growth ofVallisneriaspiralisin the Zhushan and Meiliang bays. There was only one kind of submerged vegetation (i.e.,Potamogetonorispus) in Meiliang Bay in recent years. No submerged vegetation was found in the central part of the lake. The submerged vegetation was widely distributed in East Taihu Lake, with the most species. The submerged vegetation has experienced the following four stages of succession in the last 30 years:Vallisneriaspiralis-Potamogetonaceae,Potamogetonpusillus,CeratophyllumDemersum-Elodeanuttalli, andPotamogetonaceae-Elodeanuttalli. Eutrophication and aquatic breeding are the main factors leading to the succession of submerged vegetations in Taihu Lake.
submerged vegetation; remote sensing; dominant species; Taihu Lake
10.3880/j.issn.1004-6933.2016.05.023
王琪(1990—),男,硕士研究生,研究方向为遥感技术与应用。E-mail: wangqi099x@163.com
X524
A
1004-6933(2016)05-0123-07
2015-11-23编辑:王芳)
