DNA条形码技术鉴定中国地方鸡品种的重新评估
2016-10-14黄勋和陈洁波何丹林张细权钟福生
黄勋和,陈洁波,何丹林,张细权,钟福生
DNA条形码技术鉴定中国地方鸡品种的重新评估
黄勋和1,陈洁波1,何丹林2,张细权2,钟福生1
(1嘉应学院生命科学学院,广东梅州 514015;2华南农业大学动物科学学院,广州 510642)
【目的】探讨COI基因作为标准DNA条形码技术鉴定外形差异较小的地方鸡品种的可行性。【方法】以华南地区9种优质地方鸡(怀乡鸡、清远麻鸡、惠阳胡须鸡、中山沙栏鸡、阳山鸡、杏花鸡、五华三黄鸡、文昌鸡和广西三黄鸡)和国外引进品种隐性白羽鸡为试验材料,测定标准的DNA条形码技术的线粒体细胞色素C 氧化酶亚基I (cytochromeoxidase subunit I,COI),同时下载已发表的31条家鸡和原鸡及绿头鸭的COI基因序列,分析品种遗传多样性与遗传距离,构建单倍型中介网络图和系统发生邻接树,界定区分品种特异的单倍型。【结果】除去PCR引物序列,获得了695 bp COI基因片段。根据标准的DNA条形码序列,截取648 bp 线粒体COI基因序列进行分析。10个鸡品种203个个体共检测到110个变异位点,占分析位点的16.98%,其中90个单一位点突变,20个简约信息位点。平均核苷酸多样性为0.00394(0.00349—0.00560),平均单倍型多样性为0.832(0.763—0.905),其中五华三黄鸡最高,中山沙栏鸡次之,文昌鸡最低。定义了84种单倍型,单倍型1为9个地方鸡种所共享,出现频率为64次;单倍型9和5为家鸡和隐性白羽鸡共享,出现频率分别为29次和19次;每个鸡品种均有品种特异的单倍型。广西三黄鸡、五华三黄鸡与中山沙栏鸡的单倍型数最多,为13个,隐性白羽鸡与清远麻鸡的最少,为8个。不同品种的单倍型分布差异较大,如杏花鸡的单倍型主要分布在1,清远麻鸡主要分布在1和9,惠阳胡须鸡主要分布在1、5和9,隐性白羽鸡主要分布在9和79。10个鸡种品种间遗传距离范围为0.003—0.006,净遗传距离为0—0.003;鸡品种间的遗传距离一般大于鸡品种内的遗传距离;绿头鸭与鸡品种间的遗传距离大于0.2。中介网络图将84个单倍型分为3条进化枝,呈现出一定的品种特异性,如以单倍型9为起点的进化枝没有广西三黄鸡和文昌鸡分布,但另外两枝未表现出此特征;1为祖先单倍型,由此逐渐衍生出其他单倍型。邻接树显示中国家鸡与红原鸡聚为一簇,与黑尾原鸡、灰原鸡和绿原鸡分开;中国地方鸡聚为同一簇,且存在明显的交叉现象,无显著的品种特异性。【结论】COI基因可作为研究鸡品种遗传多样性的候选分子标记。仅依靠标准的DNA条形码技术无法有效区分差异外形较小的地方鸡种,需要联合多种分子标记如COI基因、细胞色素b、 AFLP指纹技术、微卫星位点LEI0258、基因组SNP和品种特异的外貌特征。
线粒体COI基因;DNA条形码;地方鸡种;品种鉴定;遗传多样性
0 引言
【研究意义】作为分布最广泛的家禽,家鸡()在人类生产生活中扮演着重要角色。中国家鸡资源丰富,仅列入《中国畜禽遗传资源志·家禽志》的地方鸡品种就有107个[1]。随着外来引进品种和大量商品鸡的饲养,地方鸡的种质特性受到极大的影响。寻求快速简便的区分外形多样、遗传复杂的地方鸡品种的技术方法是科学家和育种专家急需解决的问题。由于线粒体细胞色素C 氧化酶亚基I(cytochromeoxidase I,COI)具有相对保守与稳定的变异性的特性,已作为DNA条形码(DNA barcoding)研究的标准基因广泛应用于脊椎动物和无脊椎动物的系统分类、种类鉴别、群体遗传多样性和分子进化学研究[2-7]。【前人研究进展】自加拿大生物学家Hebert等[8]首先倡导将648 bp COI基因作为DNA条形码编码技术的标准基因应用于生物物种鉴定以来,获得了蓬勃的发展。目前BOLD系统(The Barcode of Life Data System)记录的标本量达5 259 477件,其中条形码研究3 881 860,物种数为235 994[9]。Kerr等[10]通过研究643种北美鸟类COI基因,发现94%的种类拥有明显的条形码分支,剩余的6%大部分是有规律杂交的物种。Ward[11]总结了FISH-BOL(The Fish Barcode of Life Initiative)的研究成果表明,DNA条形码技术可以有效区分98%的海洋鱼类和93%的淡水鱼类。WILSON等[12]建立了马来西亚半岛鳞翅目蝴蝶的DNA条形码参考数据库并定义了亚种的参考阈值。孟玮等[13-16]分别验证了COI基因作为DNA条形码在鱼类物种鉴定、鸟类分类、银鲳群体遗传多样性、动物药材鉴定等方面的可行性。动物品种鉴定方面,高玉时等[2,17-18]采用DNA条形码技术验证了差异较大的地方鸡品种鉴定的可行性和有效性。【本研究切入点】之前的地方鸡DNA条形码鉴定研究主要针对于外观差异较大、地缘分布较远的品种,而对于外观相近,地缘集中的品种则尚未开展研究。【拟解决的关键问题】本研究以华南9个优质地方鸡和国外引进品种隐性白羽鸡作为试验材料,采用标准的DNA条形码技术对10个鸡品种进行分子鉴定,评估COI基因作为DNA条形码鉴定外形差异较小的鸡品种的可行性。
1 材料与方法
1.1 试验材料
以华南9种优质地方鸡和国外引进品种隐性白羽鸡共203个个体为试验材料(表1)。除五华三黄鸡之外,其他8种地方鸡种均入选《中国畜禽遗传资源志·家禽志》[1],而五华三黄鸡入选《中国禽类遗传资源》[19]。五华三黄鸡、阳山鸡采自其原产地,文昌鸡、清远麻鸡采自商品群,其余6种采自保种场,五华三黄鸡的样品类型为羽毛,其他品种均为血液;从基因库下载已发表的鸡品种如黄郎鸡、桃源鸡、仙居鸡等和原鸡如红原鸡、灰尾原鸡和绿原鸡等的COI基因全序列,同时下载绿头鸭COI基因作为计算遗传距离的参考外群(http://www.ncbi.nlm.nih.gov/nuccore/)。使用试剂盒HiPure Blood/Tissue DNA Mini Kit(美基生物,广州)提取样品基因组DNA,-20℃保存备用。
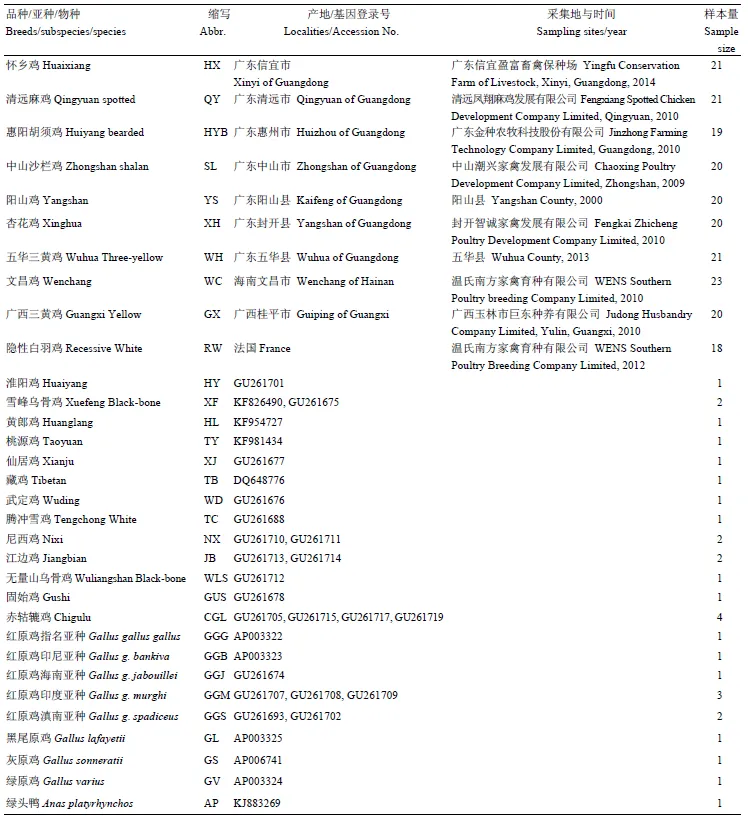
表1 样品信息
1.2 PCR扩增与序列测定
为提高PCR扩增的特异性和效率,参照鸟类DNA条形码通用引物和已发表的黄郎鸡线粒体全基因组序列(GU261701)微调COI基因的扩增引物(BirdF1:5′-TTCTCCAACCACAAAGACATTGGCAC-3′[20],COIR:5′- CGTGGGAAATTATTCCGAAACCTG -3′)。PCR反应体系为30 µL,含10×PCR buffer 3µL,dNTP mixture(2.5 mmol·L-1)2.4 µL,引物各0.3 µL(20 µmol·L-1),rTaq DNA聚合酶(宝生物, 大连)1.5 U,DNA模板100 ng。扩增条件为94℃预变性4 min,然后35个循环(94℃变性30 s,58℃退火1 min,72℃延伸50 s),最后72℃延伸7 min。PCR产物于1.5%琼脂糖凝胶电泳检测后送上海立菲生物技术有限公司(广州)双向测序。
1.3 序列分析
Bioedit 软件读取测序序列,对每个序列进行人工逐个碱基检查校对,并通过每个样品双向测序和每个单倍型重新测序进行双重校正。采用标准的648 bp DNA条形码序列(对应小鼠COI基因第58—705位碱基)作为分析数据。所获序列用Clustal X软件[21]进行对位排列后,用DnaSP5.0软件[22]定义单倍型,计算单倍型多样性(Haplotype diversity,)、核苷酸多样性(nucleotide diversity,π)和核苷酸差异均数()。通过NETWORK 4.6.1.4 软件[23]构建单倍型中介连接网络图(Median-joining networks)。以绿头鸭为外群(,KJ883269),用MEGA6.0 软件[24]中的Kimura 2-parameter(K2P)模型计算遗传距离;应用邻接法(neighbor-joining,NJ)构建系统进化树。
2 结果
2.1 COI基因序列的核苷酸位点变异、核苷酸多样性和单倍型多样性
除去PCR引物序列,获得了695 bp COI基因片段。根据标准的DNA条形码序列,截取648 bp 线粒体COI基因序列进行分析。10个鸡种203个个体线粒体COI基因共检测到110个变异位点,占分析位点的16.98%,其中单一位点突变90个(2型20个,3型2个),简约信息位点20个。五华三黄鸡的平均核苷酸差异数最大,为3.629,文昌鸡的最小,为1.455;相应地,五华三黄鸡的核苷酸多样性最高,文昌鸡的最小(表2)。在110个变异位点中,定义了84种单倍型,其中单倍型1出现频率最高,为64次(9个地方鸡种所共有,隐性白羽鸡除外),其次是单倍型9,为29次(8个鸡种共有,杏花鸡、文昌鸡除外),第三是单倍型5,为19次(9个鸡种共有,清远麻鸡除外)(表3)。广西三黄鸡、五华三黄鸡与中山沙栏鸡的单倍型数最多,为13个,隐性白羽鸡与清远麻鸡的最少,为8个。不同品种的单倍型分布差异较大,如隐性白羽鸡单倍型主要分布在9和79,清远麻鸡主要分布在1和9,惠阳胡须鸡主要分布在1、5和9,杏花鸡主要分布在1(表3)。
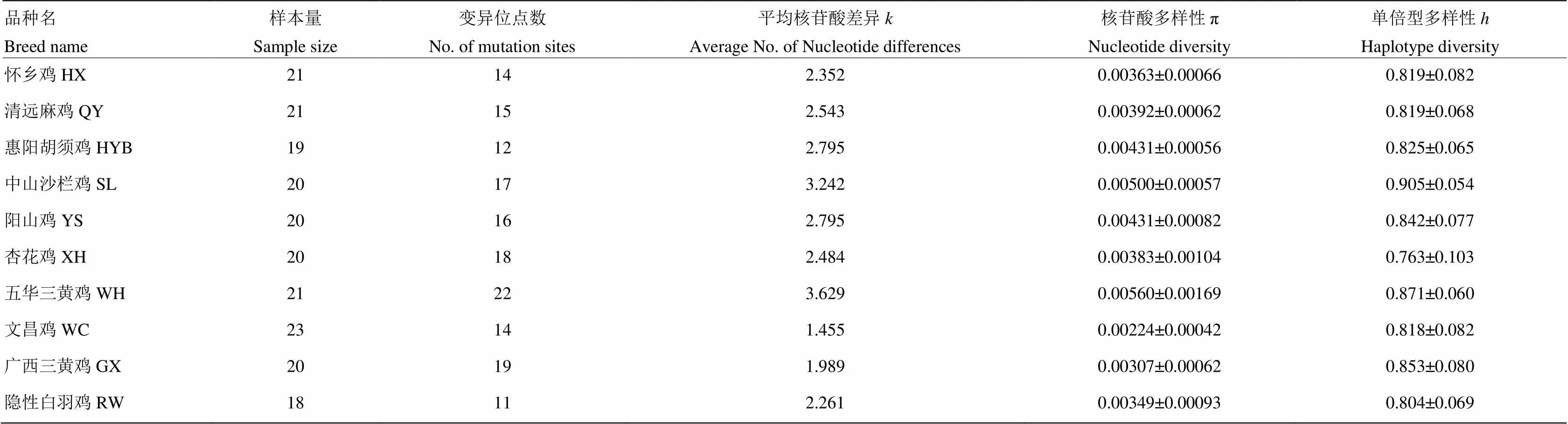
表2 10个鸡种COⅠ基因序列变异位点数、平均核苷酸差异、核苷酸多样性和单倍型多样性
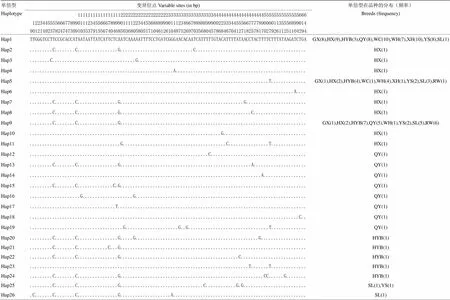
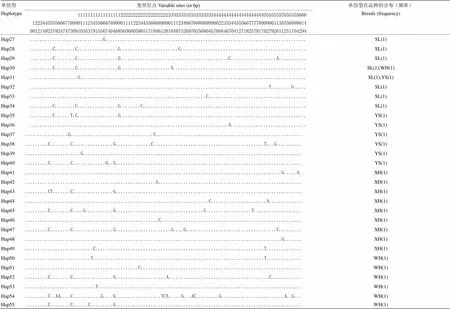
表3 10个鸡品种线粒体COI基因的单倍型及其在不同品种的分布
续表3 Continued table 3
单倍型Haplotype变异位点Variable sites (in bp)单倍型在品种的分布(频率)Breeds (frequency) 1111111111111111122222222222222222223333333333333333334444444444444444444455555555555555666122344555566677899011123455666789990111122344556888999011123466788888999002233344555667777890000113555689901490121492378247473091035379155674046850368058051710461261048752697035680457868467041271825781702792611251104294 Hap27.............................G................................................................................SL(1) Hap28.........C........C................G.......................G..................................................SL(1) Hap29.........C........C................G...........................................C..................A...........SL(1) Hap30.........C........C................G....................A.....................................................SL(1),WH(1) Hap31...................G..........................................................................................SL(1),YS(1) Hap32...............................................................................................T........G.....SL(1) Hap33......................................................................C.......................................SL(1) Hap34.........C........C................G........C.................................................................SL(1) Hap35.........C......T.C................G..........................................................................YS(1) Hap36...............................................................................A..............................YS(1) Hap37.................G.................................C..........................................................YS(1) Hap38.........C........C................G..............C............................................T...G..........YS(1) Hap39......................G.......................................................................................YS(1) Hap40.........C........C.............G..G..........................................................................YS(1) Hap41......................................................................................................G.....A.XH(1) Hap42....................................................G.........................................................XH(1) Hap43.........CT.......C................G..........................................................................XH(1) Hap44.........................................................................C......................A.............XH(1) Hap45.........C........C....G...........G...................................C..................T...................XH(1) Hap46.....................................................C........................................................XH(1) Hap47.........C........C................G......................G....G....................................C.........XH(1) Hap48......................................................................................................G.......XH(1) Hap49...........................C...................................................................T..............XH(1) Hap50..........................T....................................................................T..............WH(1) Hap51.............................................C................................................................WH(1) Hap52.........C........C................G....................A........................................C............WH(1) Hap53............................T.................................................................................WH(1) Hap54.........C..AA....C...........G....G..................TCA.....G...AC.........G.........................A..G...WH(1) Hap55.........C........C......C.........G..........................................................................WH(1)
续表3 Continued table 3
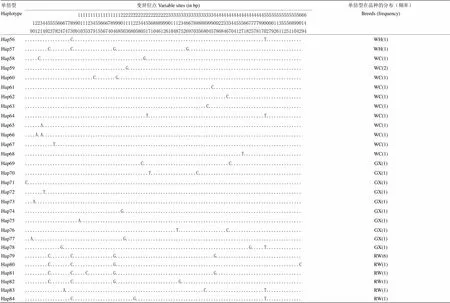
单倍型Haplotype变异位点Variable sites (in bp)单倍型在品种的分布(频率)Breeds (frequency) 1111111111111111122222222222222222223333333333333333334444444444444444444455555555555555666122344555566677899011123455666789990111122344556888999011123466788888999002233344555667777890000113555689901490121492378247473091035379155674046850368058051710461261048752697035680457868467041271825781702792611251104294 Hap56..................C............................................................................T..............WH(1) Hap57.........C........C................G............................G.............................................WH(1) Hap58.....C.........................................G..............................................................WC(1) Hap59........................................G.....................................................................WC(2) Hap60...........................C........G.........................................................................WC(1) Hap61..........................................................................C...................................WC(1) Hap62................................................................................C.............................WC(1) Hap63........................................................................C.....................................WC(1) Hap64................................................T..............................................T..............WC(1) Hap65......A.......................................................................................................WC(1) Hap66....A.A.......................................................................................................WC(1) Hap67...........T..................................................................................................WC(1) Hap68......................................................................................T.......................WC(1) Hap69..............................................C..................................C............................GX(1) Hap70.................................................T..................C.........................................GX(1) Hap71C.............................................................................................................GX(1) Hap72.......T......................................................................................................GX(1) Hap73...A..........................................................................................................GX(1) Hap74......................................G.......................................................................GX(1) Hap75.....................A........................................................................................GX(1) Hap76............................................................T...................C.............................GX(1) Hap77..A....................................G......................................................................GX(1) Hap78..............G..........................................................................G.....T..............GX(1) Hap79.........C........C................G.......................................G..................................RW(6) Hap80.........C........C................G.........................................................................CRW(1) Hap81.........C........C.....C..........G.......................................G..................................RW(1) Hap82.........C........C................G.........................G................................................RW(1) Hap83...............A.......................................................C.......................T..............RW(1) Hap84..................C........................G...................................................T..............RW(1)
圆点表示与单倍型Hap1具有相同的碱基 Dots within the nucleotide position indicate the same nucleotides as given in Hap1
2.2 10个鸡品种群体内与群体间遗传距离
对10个鸡品种COI基因序列K2P距离和净遗传距离(Da)研究显示,绿头鸭与鸡品种的遗传距离明显大于鸡品种之间的遗传距离(表4)。鸡品种间遗传距离范围为0.003—0.006,隐性白羽鸡与9种地方鸡种的遗传距离较远;五华三黄鸡品种内的遗传距离最大,为0.006,文昌鸡品种内遗传距离最小,为0.002。10个鸡种品种净遗传距离为0—0.003(表4)。
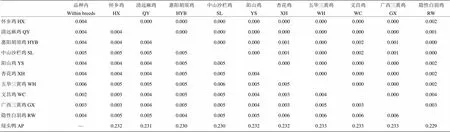
表4 10个鸡品种间Kiumura 双参数距离和净遗传距离(Da)
上三角为净遗传距离Da,下三角为Kiumura 双参数距离
Above dialogue was net distance (Da), down diagonal was K2P distance of COIgene among 10 chicken breeds
2.3 COI基因中介网络图和系统发育树
利用表3的单倍型数据构建中介网络图。中介网络图主要分为3个进化枝,第1枝是以Hap1为中心节点,另外2枝是从Hap1衍生出来的以Hap5、Hap9为中心节点(图1)。Hap1最有可能是祖先单倍型,逐渐衍生出其他单倍型。以Hap9为中心的进化枝表现出一定的品种特异性,如文昌鸡和广西三黄鸡不在此枝;另外两枝则未表现出此特征。
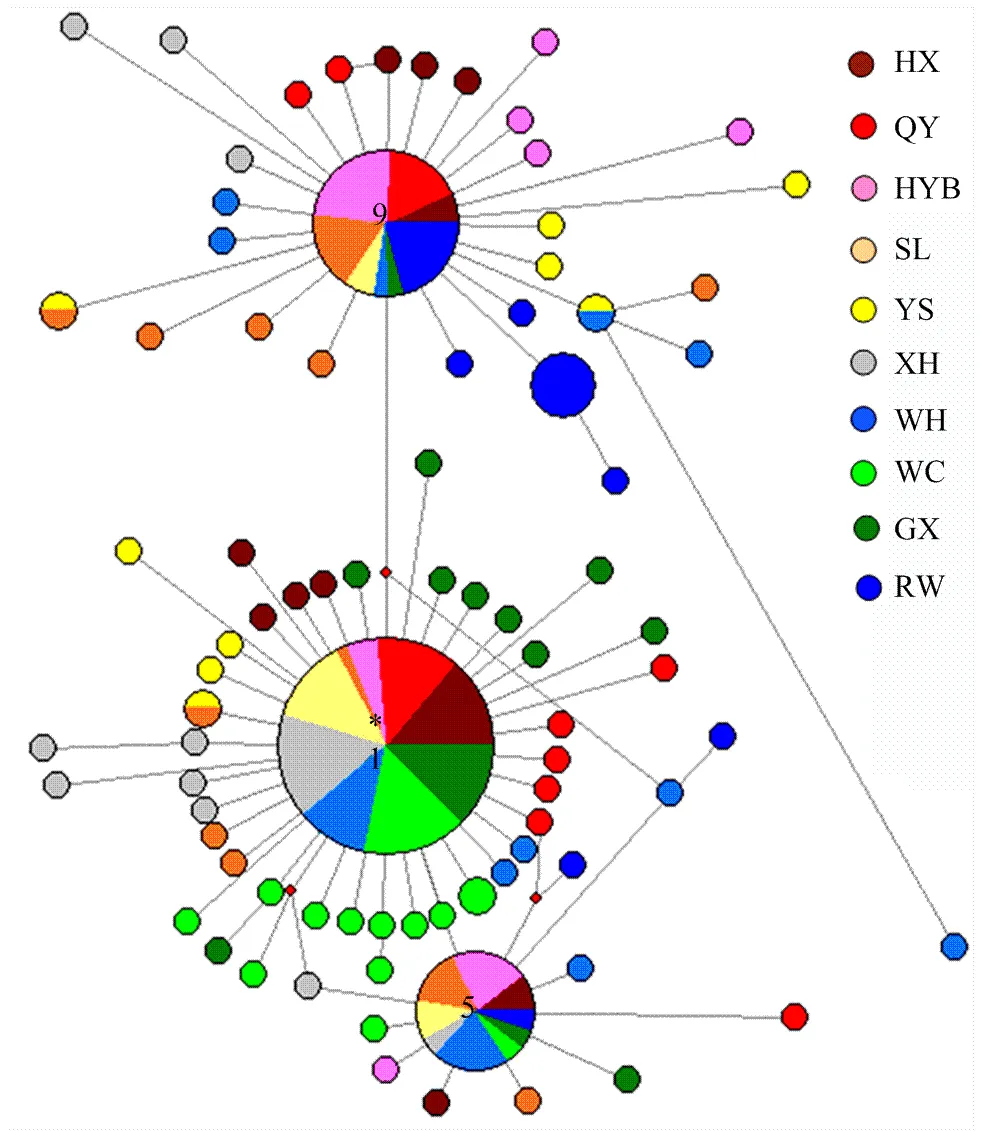
红色方块是软件NETWORK生成的媒介载体,代表在实际样本未观察到的推测的中间单倍型。圆的大小对应于单倍型频率。不同鸡品种用不同颜色标注
采用K2P模型构建本研究定义的单倍型(每个品种所有的单倍型,共121条序列)和下载已发表的鸡COI基因序列(648 bp,29条序列)的系统发育树(邻接树)。发育树分为三枝,第一枝是黑尾原鸡和灰原鸡,第二枝是绿原鸡,第三枝是包括五个红原鸡亚种在内的地方鸡(图2)。第三枝又细分为两枝,存在亚种/品种的个体交叉现象,没有发现亚种/品种特异性。
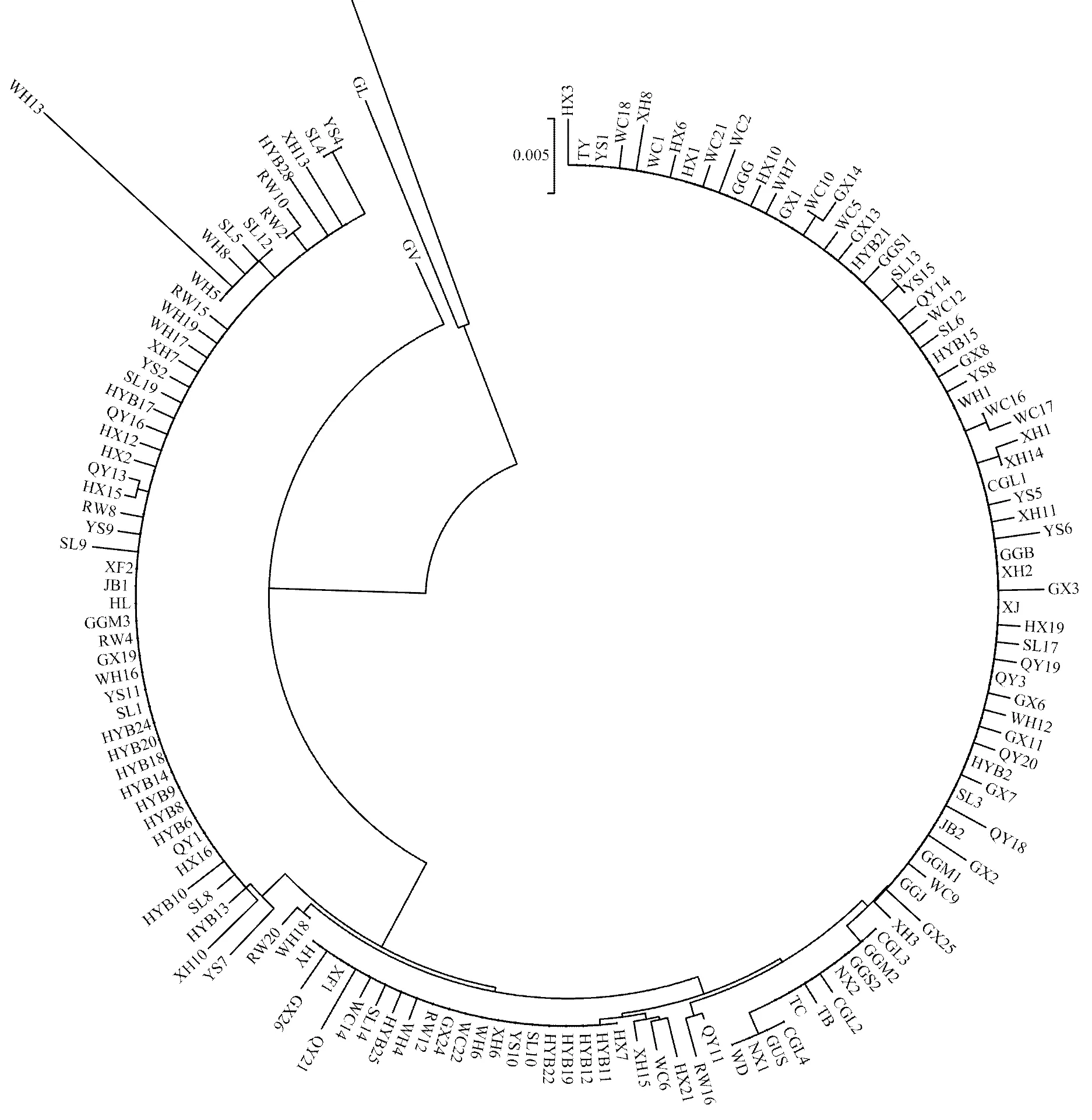
品种名称缩写见表1,数字表示样品编号 The breed Abbr. please see Table 1 in details, digit represent the sample number
3 讨论
3.1 10个鸡品种COI基因序列的遗传多样性
本研究获得了9个地方鸡种和1个引进鸡品种共203个个体的648 bp线粒体COI基因序列,共检测到110个变异位点,定义了84种单倍型。其中五华三黄鸡的遗传多样性最高,中山沙栏鸡次之,文昌鸡最低。由于分布在偏远山村,基因交流较少,未进行系统选育且样品为随机采集如五华三黄鸡[25],或采自祖代群如中山沙栏鸡,或采样时间较早如阳山鸡,这些品种保留着较高的遗传多样性。而来自商品群的地方鸡遗传多样性则较低,如文昌鸡(品种内遗传距离0.002,表4)。84种单倍型在不同的品种的分布有所差异,如单倍型1出现64次,除了隐性白羽鸡,其他9个地方鸡都有分布;其次是单倍型9和5;隐性白羽鸡主要分布在单倍型9和79,清远麻鸡主要分布在单倍型1和9,惠阳胡须鸡主要分布在单倍型1、5和9,杏花鸡分布在单倍型1,而其他7个品种单倍型分布较为分散(表3)。本文研究的鸡品种的COI基因序列变异位点数、平均核苷酸差异、核苷酸多样性和单倍型多样性比同等类型研究相近或更高[2,17-18],说明本文选取的鸡品种保护潜力较大,证明标准DNA条形码用于研究鸡品种遗传多样性的有效性。另外,以绿头鸭为外群计算遗传距离时,绿头鸭与鸡品种间的遗传距离明显大于鸡品种间的遗传距离,表明COI基因可作为计算鸡品种间的遗传距离的分子标记。
3.2 中国地方鸡品种COI基因系统发生关系
邻接树显示中国地方鸡与红原鸡分为两枝,虽然有些品种分布较为集中(如惠阳胡须鸡),但均存在亚种/品种的个体交叉现象(图2)。当提高建树可信度时(1000步bootstrap、50% cut-off),所有中国地方鸡与红原鸡聚为一枝(数据未显示),说明应用标准的DNA条形码技术无法将中国地方鸡品种有效区分开来。而中国地方鸡和红原鸡与黑尾原鸡、灰原鸡、绿原鸡分开,间接支持了中国地方鸡与红原鸡亲缘关系较近的观点。主要原因可能有两点:一是本研究的9个地方鸡种在广东饲养量较大,因地域邻近和经贸频繁,不同的鸡种在品种形成与进化过程中不可避免存在不同程度的杂交,出现了个别品种间的交叉现象(图2);二是COI基因的相对保守性,品种间的遗传差异较小(表4)。在本研究中,虽然部分单倍型为特定品种所有,如单倍型12—19为清远麻鸡特有,但这些单倍型与其他鸡品种部分单倍型聚在同一枝(图2);并且该结果是基于有限的样品(10个品种,每个品种18—23只,共203只)得出的,在增加样品数量或品种后是否还是清远麻鸡特有还有待研究。因此,杂交使得亚种/品种鉴定变得困难,分析时需要在足够的样品数量和联合多种方法后才能得出比较可靠的结论[2,12]。
3.3 应用COI基因鉴定地方鸡品种
研究表明DNA条形码对鉴定差异较大的鸡品种鉴定具有可行性和有效性[2,26]。在相近的COI基因片段长度下,本研究在10个鸡种203只个体共检测到110个变异位点,多于15个差异较大的鸡种227个个体的38个变异位点[2],说明本研究采用的COI基因片段信息含量较高。虽然不同的品种有特异的单倍型,但分布不集中,且大多只有1个个体(隐性白羽鸡除外,单倍型79有6个个体),因此,无法确定是否能够作为品种鉴定的诊断依据,需加大样本量进行验证。在本研究的9种华南家鸡中,外形特征较为显著,同时按照品种标准采集样品,因同品种异名而导致COI无法区分开这9种地方鸡品种的可能性极低。此外,有趣的是,单倍型1为9个地方鸡品种所共有,隐性白羽鸡除外,可作为中国地方鸡品种的鉴定依据。由于地方鸡品种复杂的形成与进化历史和COI基因自身的属性,运用标准的DNA条形码技术无法鉴定本研究的9个亲缘关系较近、地域邻近的华南地方鸡种。因此,有效的品种鉴定还需联合多种分子标记如COI基因[26]、AFLP指纹技术[27]、细胞色素b[28]、LEI0258[29]、基因组SNP[30]等,以及与品种特定的外形特征如清远麻鸡的“一楔、二细、三麻身”、杏花鸡的“两细(头细、脚细)、三黄(羽黄、脚黄、喙黄)、三短(颈短、体躯短、腿短)”。
4 结论
COI基因可用于地方鸡的遗传多样性研究,但标准的DAN条形码技术无法有效区分差异较小的鸡品种,需要联合其他分子标记以及外形特征进行鉴定。
致谢:感谢邹志冠、刘少丰提供部分试验样品,感谢杜炳旺教授、古文良、陈浩、刘怡然、温金星、黄丰勋、徐迪宗、陈勇杰、曾德鑫在收集样品时提供的帮助。
References:
[1] 国家畜禽遗传资源委员会. 中国畜禽遗传资源志·家禽志. 北京: 中国农业出版社, 2011.
China National Commission of Aniamal Genetic Resources.:. Beijing: China Agriculture Press, 2011. (in Chinese)
[2] 高玉时, 唐修君, 屠云洁, 陆俊贤, 薛茂云, 施祖灏, 张小燕. 基于线粒体COI基因15个鸡种的DNA 编码研究. 中国农业科学, 2011, 44(3): 587-594.
GAO Y S, TANG X J, TU Y J, LU J X, XUE M Y, SHI Z H, ZHANG X Y. Studies on the DNA barcoding of fifteen chicken breeds by mtDNA COI gene., 2011, 44(3):587-594. (in Chinese)
[3] TAVARES E S, GONÇALVES P, MIYAKI C Y, BAKER A J. DNA barcode detects high genetic structure within Neotropical bird species., 2011, 6(12): e28543.
[4] BUCKLIN A, STEINKE D, BLANCO-BERCIAL L. DNA barcoding of marine metazoa., 2011, 3(1): 471-508.
[5] LERAY M, KNOWLTON N. DNA barcoding and metabarcoding of standardized samples reveal patterns of marine benthic diversity., 2014, 112(7): 2076-2081.
[6] SAITOH T, SUGITA N, SOMEYA S, IWAMI Y, KOBAYASHI S, KAMIGAICHI H, HIGUCHI A, ASAI S, YAMAMOTO Y, NISHIUMI I. DNA barcoding reveals 24 distinct lineages as cryptic bird species candidates in and around the Japanese Archipelago., 2015, 15(1): 177-186.
[7] BAMANIYA D C, PAVAN-KUMAR A, GIREESH-BABU P, SHARMA N, REANG D, KRISHNA G, LAKRA W S. DNA barcoding of marine ornamental fishes from India.,:,,, 2016, 27(5): 3093-3097.
[8] HEBERT P D, RATNASINGHAM S, DEWAARD J R. Barcoding animal life: cytochromeoxidase subunit 1 divergences among closely related species., 2003, 270(S1): S96-S99.
[9] RATNASINGHAM S, HEBERT P D. BOLD: The barcode of life data system., 2007, 7(3): 355-364.
[10] KERR K C, STOECKLE M Y, DOVE C J, WEIGT L A,FRANCIS C M, HEBERT P D. Comprehensive DNA barcode coverage of North American birds., 2007, 7(4): 535-543.
[11] WARD R D. FISH-BOL, a case study for DNA barcodes., 2012, 858: 423-439.
[12] WILSON J J, SING K W, SOFIAN-AZIRUN M. Building a DNA barcode reference library for the true butterflies (Lepidoptera) of peninsula malaysia: What about the Subspecies?, 2013, 8(11): e79969.
[13] 孟玮, 杨天燕, 海萨, 海沙尔·阿那斯. 基于线粒体COI基因序列的亚东鲑DNA条形码研究. 水产学杂志, 2010, 23(1): 6-10.
MENG W, YANG T Y, HAI S, HAI S E. Study of DNA barcoding based on the mitochondrial COI Gene Sequences in., 2010, 23(1): 6-10. (in Chinese)
[14] 马明义, 闫颖, 王译伟, 李静, 蔡延森, 李佳凌. 我国32 种鸟类DNA条形码分析. 四川动物, 2012, 31(5): 729-733.
MA M Y, YAN Y, WANG Y W, LI J, CAI Y S, LI J L. A study of DNA barcoding on 32 species of bird in China., 2012, 31(5): 729-733. (in Chinese)
[15] 彭士明, 施兆鸿, 侯俊利. 基于线粒体D-loop区与COI基因序列比较分析养殖与野生银鲳群体遗传多样性. 水产学报, 2010, 34(1): 19-25.
PENG S M, SHI Z H, HOU J L. Comparative analysis on the genetic diversity of cultured and wild silver pomfret populations based on mt D-loop and COI gene., 2010, 34(1): 19-25. (in Chinese)
[16] 张辉, 姚辉, 崔丽娜, 杜鹤, 林喆, 高晓晨, 郎雪, 宋经元, 罗焜, 石林春, 陈士林. 基于COI条形码序列的《中国药典》动物药材鉴定研究. 世界科学技术-中医药现代化, 2013, 15(3): 371-380.
ZHANG H, YAO H, CUI L N, DU H, LIN Z, GAO X C, LANG X, SONG J Y, LUO K, SHI L C, CHEN S L. Application of COI-based DNA barcoding for identifying animal medical materials in the Chinese pharmacopoeia., 2013, 15(3): 371-380. (in Chinese)
[17] 屠云洁, 高玉时, 周新民, 张学余, 王克华, 唐修君. 我国6个地方鸡品种线粒体COI基因遗传多样性分析. 扬州大学学报(农业与生命科学版), 2007, 28(3): 31-33.
TU Y J, GAO Y S, ZHOU X M, ZHANG X Y, WANG K H, TANG X J. The genetic diversity analysis of mtDNA COI genes in six indigenous chicken breeds in China., 2007, 28(3): 31-33. (in Chinese)
[18] 高玉时, 屠云洁, 童海兵, 王克华, 陈宽维, 顾荣. 6个地方鸡种线粒体COI基因的DNA条形码. 农业生物技术学报, 2007, 15(6):924-930.
GAO Y S, TU Y J, TONG H B, WANG K H, CHEN K W, GU R. DNA barcoding application of mtDNA COI Gene in identifying six indigenous chicken breeds in China., 2007, 15(6): 924-930. (in Chinese)
[19] 陈国宏, 王克华, 王金玉, 丁铲, 杨宁. 中国禽类遗传资源. 上海: 上海科学技术出版社, 2004.
CHEN G H, WANG K H, WANG J Y, DING C, YANG N. Shanghai: Shanghai Scientific and Technical Publishers, 2004. (in Chinese)
[20] HEBERT P D, STOECKLE M Y, ZEMLAK T S, FRANCIS C M. Identification of birds through DNA barcodes., 2004, 2(10): e312.
[21] THOMPSON J D, GIBSON T J, PLEWNIAK F, JEANMOUGIN F, HIGGINS D G. The Clustal_X windows interface: flexible strategies for multiple sequence alignment aided by quality analysis tools., 1997, 25(24): 4876-4882.
[22] LIBRADO P, ROZAS J. DnaSP v5: a software for comprehensive analysis of DNA polymorphism data., 2009, 25(11): 1451-1452.
[23] BANDELT H J, FORSTER P, RÖHL A. Median-joining networks for inferring intraspecific phylogenies., 1999, 16(1): 37-48.
[24] TAMURA K, STECHER G, PETERSON D, FILIPSKI A, KUMAR S. MEGA6: Molecular evolutionary genetics analysis version 6.0., 2013, 30(12): 2725-2729.
[25] 黄勋和, 李威娜, 陈珊, 陈洁波, 钟福生. 五华三黄鸡群体遗传多样性与遗传结构分析. 中国家禽, 2016, 38(1): 56-58.
HUANG X H, LI W N, CHEN S, CHEN J B, ZHONG F S. Population genetic diversity and genetic structure of Wuhua three- yellow chicken., 2016, 38(1): 56-58.(in Chinese)
[26] BONDOC O L, SANTIAGO R C. The use of DNA barcodes in the evolutionary analysis of domestic breeds and strains of chicken () in the Philippines., 2012, 95(4): 358-369.
[27] 高玉时, 屠云洁, 钱勇, 李慧芳, 陈宽维, 童海兵. 12个地方鸡种遗传多态性AFLP指纹分析. 农业生物技术学报, 2006, 14(4): 498-502.
GAO Y S, TU Y J, QIAN Y, LI H F, CHEN K W, TONG H B. AFLP Fingerprinting analysis of genetic polymorphism in 12 indigenous chicken breeds., 2006, 14(4): 498-502. (in Chinese)
[28] YACOUB H A, FATHI M M, SADEK M A. Using cytochromegene of mtDNA as a DNA barcoding marker in chicken strains., 2015, 26(2): 217-223.
[29] HAN B, LIAN L, QU L J, ZHENG J X, YANG N. Abundant polymorphisms at the microsatellite locus LEI0258 in indigenous chickens., 2013, 92(12): 3113-3119.
[30] KWAK W, SONG K D, OH J D, HEO K N, LEE J H, LEE W K, YOON S H, KIM H, CHO S, LEE H K. Uncovering genomic features and maternal origin of korean native chicken by whole genome sequencing., 2014, 9(12): e114763.
(责任编辑 林鉴非)
DNA Barcoding of Indigenous Chickens in China: a Reevaluation
HUANG Xun-he1, CHEN Jie-bo1, HE Dan-lin2, ZHANG Xi-quan2, ZHONG Fu-sheng1
(1School of Life Sciences, Jiaying University, Meizhou 514015, Guangdong;2College of Animal Science, South China Agricultural University, Guangzhou 510642)
【Objective】 The aim of this study is to determine the feasibility of utility of mitochondrial cytochromeoxidase subunit I (COI) gene as DNA barcoding to identify indigenous chicken breeds with nearer appearances.【Method】 COI gene of 648 bp in length was obtained from nine indigenous chicken breeds of South China (Huaixiang, Qingyuan spotted, Huiyang bearded, Zhongshan shalan, Yangshan, Xinghua, Wuhua three-yellow, Wenchang and Guangxi yellow) and one commercial breed, Recessive White, with the method of direct sequencing of PCR products; while other COI sequences were downloaded from GenBank, including Chinese indigenous chickens and wild jungles and mallard (). These sequences were then used to analyze genetic diversity and genetic distance, construct median-joining network and phylogenetic tree based on haplotypes, as well as define breed specific haplotypes. 【Result】 COI gene of 695 bp in length was obtained after deletion of the primers sequences. And then 648 bp of standard barcoding was used for analysis. A total of 110 mutation sites were detected from 203 individuals of 10 breeds with 16.98% in all sites, of which 90 were singleton variable sites and the remaining 20 were parsimony informative sites. The average nucleotide diversity and haplotype diversity were 0.00394 (0.00349-0.00560) and 0.832 (0.763-0.905), respectively. Wuhua three-yellow chicken had the highest levels of genetic diversity, Zhongshan Shalan chicken had the second higher one, but Wenchang chicken had the lowest ones. A total of 84 haplotypes were defined, haplotype 1 had the highest frequency in nine indigenous chicken breeds. Haplotyes 9 and 5 were sharing both in indigenous and Recessive White chicken breeds, with the frequencies of 29 and 19, respectively. Each breed had its own haplotypes. Guangxi yellow, Wuhua three-yellow and Zhongshan shalan chickens had most 13 haplotypes, while Qingyuan spotted and Recessive White had least 8 haplotypes. The distribution of haplotypes of different breeds had a little of difference. For example, the haplotypes of Xinghua chicken was mostly distributed at 1, where Qingyuan spotted, Huiyang bearded and Recessive White was mostly distributed at 1 and 9, 1, 5 and 9, and 9 and 79, respectively. The genetic distance and net genetic distance between 10 breeds were ranged from 0.003 to 0.006 and from 0 to 0.003, respectively. The genetic distance among breeds was higher than those of within a breed; those of betweenand chickens were higher than 0.2. The 84 haplotyes of median-joining networks of were classified into three clusters with the characteristic of breed specific. For example, the cluster originated from in haplotype 9 had no Guangxi yellow and Wenchang chicken breeds. Other haplotypes were the descendent of 1. The Neighbor-joining tree showed that indigenous chickens in China andwere clustered into one branch, separating from,and. No branch with breed specificity was found. 【Conclusion】The results presented herein indicated that COI gene can be used as a candidate molecular marker for elucidate genetic diversity of indigenous chickens. It’s less effectivity of utility of standard COI gene as DNA barcoding to identify indigenous chicken with nearer appearance, there is highly need to incorporate multiple molecular markers such as COI gene, Cytochrome b gene, AFLP (Amplified Fragment Length Polymorphism), SNP (Single Nucleotide Polymorphism) and breed specific appearances.
mitochondrial cytochromeoxidase subunit I gene; DNA barcode; indigenous chicken; breed identification; genetic diversity
2015-06-23;接受日期:2016-05-14
广东省自然科学基金(2014A030307018)、嘉应学院“创新强校工程”项目(CQX019)、广东省公益研究与能力建设项目(2015A020208020)
黄勋和,E-mail:hxh826@126.com。通信作者钟福生,E-mail:zfs@jyu.edu.cn
