青藏高原东北缘高寒草甸珠芽蓼斑块植被构成和种间关联
2016-10-09关士琪高红娟孔杨云于应文
魏 斌,王 莹,关士琪,高红娟,孔杨云,于应文
(草地农业生态系统国家重点实验室 兰州大学草地农业科技学院,甘肃 兰州 730020)
青藏高原东北缘高寒草甸珠芽蓼斑块植被构成和种间关联
魏 斌,王 莹,关士琪,高红娟,孔杨云,于应文
(草地农业生态系统国家重点实验室 兰州大学草地农业科技学院,甘肃 兰州 730020)
群落植物种构成和种间关系反映植物种群和群落分布格局上的互作关系。通过对珠芽蓼(Polygonumviviparum)斑块及其对照区域群落植物种重要值、物种多样性指数、生物量构成及植物种种间关联Jaccard指数和Spearman秩相关进行分析,揭示高寒草甸演替过程中的生态学机制。结果表明,珠芽蓼斑块的样方物种数显著高于对照(P<0.05),二者类群生物量与其重要值构成变化规律类似。珠芽蓼斑块的植被构成以杂类草和禾草为主,其占绿色生物量的60.9%和22.1%,且珠芽蓼种群约占绿色生物量的1/3;而对照则以禾草和杂类草为主,分别占绿色生物量的53.3%和27.1%。珠芽蓼斑块及对照群落中,显著关联的种对数分别占各自总种对数的16.4%和33.0%,多数植物种对的种间联结较松散,种对间独立性较强,二者正相关与负相关种对数比分别为0.83和1.40。珠芽蓼斑块群落处于稳定阶段,珠芽蓼斑块的形成,能增强高寒草甸的群落稳定性。
高寒草甸;珠芽蓼斑块;重要值;生物量构成;Jaccard关联度指数;Spearman秩相关
高寒草甸是青藏高原主要草地类型之一,它不仅是支撑高原畜牧业发展、维系地方经济发展的物质基础,而且对长江中下游平原和黄河、长江流域地下水、地表水的丰歉以及防止风、旱、涝、沙尘暴、水土流失等自然灾害的发生,起着不可替代的屏障作用[1]。然而,由于受气候、人口和经营方式等的影响,高寒草地的退化已威胁到当地生态环境维持、生物多样性保护和畜牧业经济发展。因此,退化高寒草甸的恢复治理是管理青藏高原高寒生态系统的关键。草地植被斑块状分布是天然放牧草地的基本特征,是植被对放牧和气候因素长期适应的结果[2-3]。植物种间联结是群落的本质特征之一,是指不同植物种在空间分布上的相关性,因群落生境差异影响物种分布而引起[4-5]。研究草地群落中斑块的群落构成特征和种间关联,反映植物物种空间上的相互关系,对于高寒草甸植被演替动态和稳定性机制的揭示具有重要意义。
近年来,在青藏高原高寒草甸上,诸多学者针对嵩草(Kobresia)和禾草斑块的植被特征[2,6],双子叶斑块的植被与土壤养分特征关系[7],禾草、莎草和双子叶植被斑块的植被构成及植物种空间分布[3,8],粪斑和鼠丘斑块植被和土壤养分变化[9-11]等进行了研究,这为草地植被斑块的深入研究提供了理论基础和分析方法。珠芽蓼(Polygonumviviparum)作为青藏高原高寒草甸草地常见植物种之一,常以斑块形式存在于草地中,在高寒草甸演替中起重要作用。诸多学者对珠芽蓼的药用价值和化学成分[12]、生理生化特性[13]以及繁殖特性[14]等方面进行了系统研究,但就珠芽蓼斑块植物种间关系的研究相对缺乏,虽然顾宁等[15]对山西芦芽山珠芽蓼群落中主要植物种的种间关联进行了分析,但尚无放牧条件下珠芽蓼斑块的植物种间关系研究。本研究以青藏高原东北缘高寒草甸草地中珠芽蓼斑块为研究对象,采用Jaccard种间关联度指数及Spearman秩相关方法,分析珠芽蓼斑块及其对照区域主要植物种的构成和种间关联,旨为珠芽蓼斑块的演替动态及稳定性机制的深入揭示提供理论基础和实践依据。
1 材料与方法
1.1研究区概况
研究区位于青藏高原东北缘的甘肃省天祝藏族自治县抓喜秀龙乡的高寒草甸上,地理坐标为102°40′-102°47′ E,37°11′-37°14′ N,海拔2 960 m。年均气温-0.1 ℃,1月均温-18.3 ℃,7月均温12.7 ℃,>0 ℃年积温1 380 ℃·d。年降水量416 mm,无绝对无霜期,仅分冷热两季,土壤为亚高山草甸土。草地主要植物种有矮嵩草(Kobresiahumilis)、线叶嵩草(K.capillifolia)、珠芽蓼、西北针茅(Stipakrylovii)、阴山扁蓿豆(Melilotoidsruthenicavar.inschanica)和球花蒿(Artemisiasmithii)等[9]。
1.2样地设置
于2015年7月中旬,在多年放牧牦牛和藏羊、草地利用率为80%~90%的冷季牧场(放牧期一般为11月-翌年4月末)的高寒草甸上,设置面积为0.3~0.5 hm2的样地3块。
1.3测定指标和方法
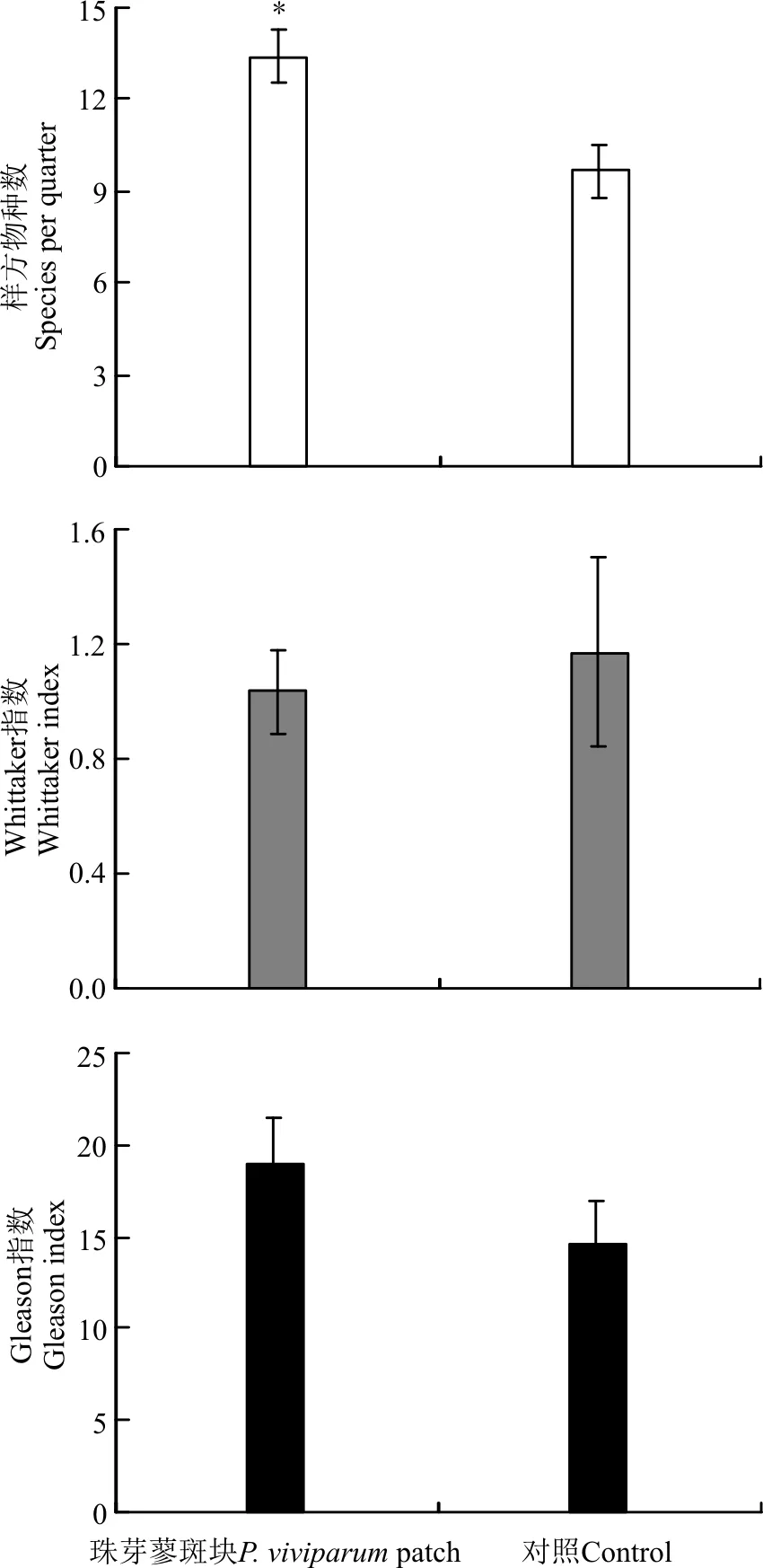
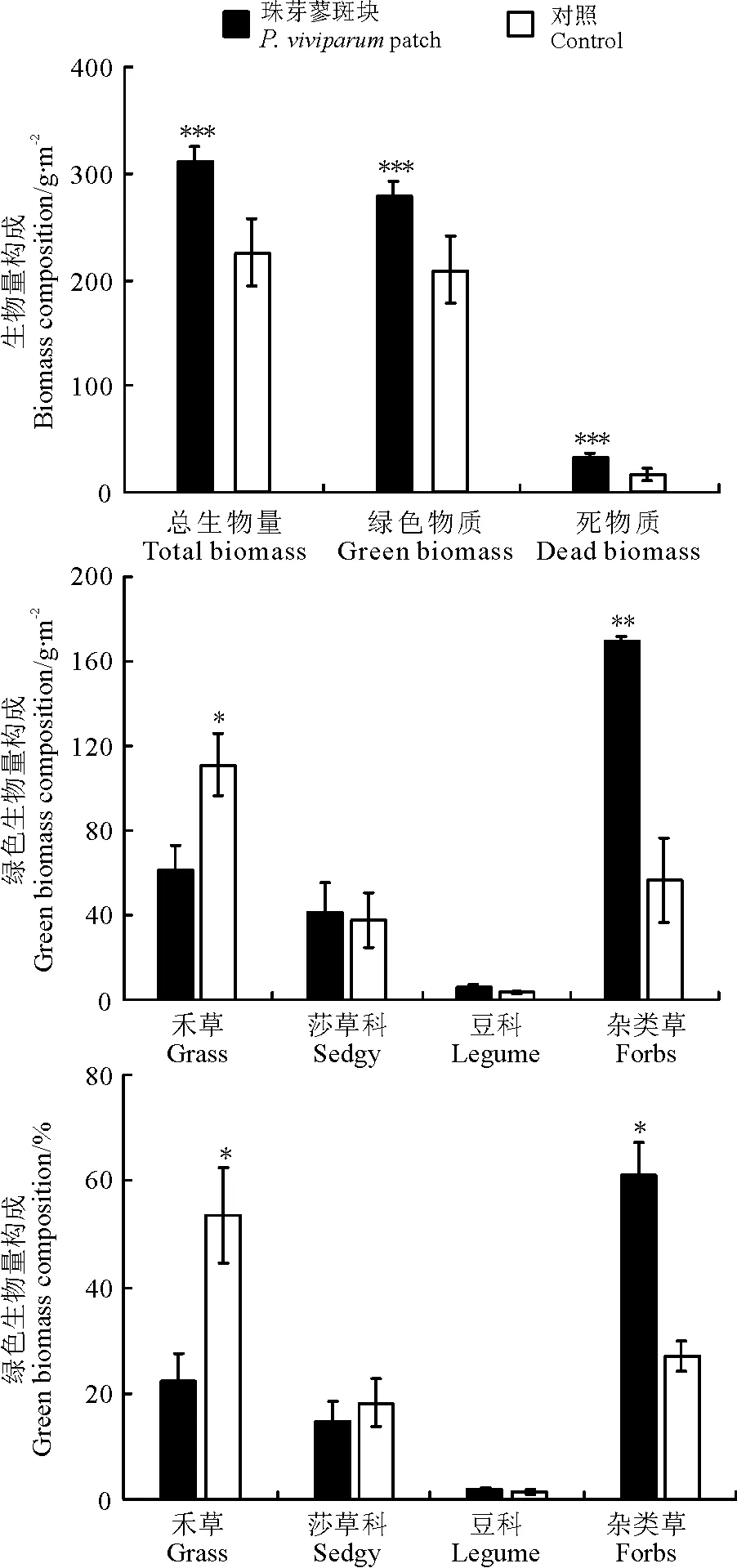
1.3.1群落构成特征2015年7月下旬-8月下旬,在各样地内,随机选择珠芽蓼斑块17个,珠芽蓼斑块的确定依据为在借鉴牟晓明等[3]和张卫国等[10]的草地斑块划分依据及高寒草甸珠芽蓼斑块实际调查基础上,将面积在1.0 m2以上的与周围植物群落组成明显不同、珠芽蓼种群聚集的斑块状草地命名为珠芽蓼斑块。以珠芽蓼斑块为中心,用0.5 m×0.5 m的样方,调查测定各珠芽蓼斑块和配对的距离珠芽蓼斑块边界约1.0 m处对照区域内所有植物种的高度、盖度和地上生物量。先将收获的地上生物量(total biomass,TB)按绿色(活)物质(green biomass,GB)和死物质(dead biomass,DB)分开,再将绿色物质按不同植物种分开,然后置入烘箱烘干称重。按公式IV=(RC+RH+RB)/3计算植物种重要值(important value),式中:IV为植物种重要值,RC、RH和RB分别为植物种的相对盖度(样方内某种的盖度/样方内所有种的盖度之和)、相对高度(样方内某种的高度/样方内所有种的高度之和)和相对生物量(样方内某种的生物量/样方内所有种的生物量之和)。
利用样方调查数据,按公式[4]DGI=S/lnA计算物种α多样性指数DGI(Gleason指数),式中:S为群落中的物种数目,A为样方面积;按公式[4]βw=(S/ma)-1计算物种β多样性指数βw(Whittaker指数),式中:S为研究系统中观测物种总数,ma为各样方或样本的平均物种数。
1.3.2植物种间关联度Jaccard指数Jaccard指数反映的是物种共同出现的百分率。采用Jaccard指数[5](简称JI指数)测定物种间关联程度,以2×2列关联表为基础计算,其公式为JI=a/(a+b+c)。式中:a为物种A和物种B同时出现的样方数;b和c分别为物种A或物种B出现一种的样方数。当a=0时,表示种A和种B不同时出现在同一样方内,即种A和种B完全独立或相异;当a=S(样方数)时,b=c=0,取值为1,表示物种A、B同时出现在所有样方中,即种A与种B绝对关联;当b=0时,表示种B与种A完全关联,即种A随种B的出现而出现,而种B可以单独出现;当c=0时,表示种A与种B完全关联,即种B随种A的出现而出现,而种A可以单独出现。本研究在借鉴张金屯[5]的划分方法基础上,将JI指数划分为4个等级,对珠芽蓼斑块主要植物种种间关联进行客观分析,0.75 1.3.3植物种间Spearman秩相关分析Spearman秩相关系数[5]属于非参数性检验,由植物种重要值计算。其公式为: 式中:r(i,k)为物种i与物种k的相关系数,N为样方数,dj=(xij-xkj),xij、xkj分别为物种i和物种k在样方j中的秩。 1.4数据分析 用Excel进行数据处理及制图。SPSS 16.0软件的One-Way ANOVA对斑块和对照区域的植物群落特征和生物量进行方差分析(F-检验)和显著性检验,数据用均值±标准误差表示。 2.1主要植物种重要值 表1 珠芽蓼斑块群落主要植物种重要值 2.2群落特征 珠芽蓼斑块和对照草地的草层高度分别为14.83±1.66和17.56±3.60 cm,植被盖度分别为98.6%±1.15%和94.8%±3.58%,且二者均在珠芽蓼斑块和对照区域之间差异不显著(P>0.05)。珠芽蓼斑块的群落物种数显著高于对照区域(P<0.05),其Gleason指数和Whittaker指数在珠芽蓼珠芽蓼斑块和对照区域之间差异不显著(P>0.05)(图1)。说明,珠芽蓼斑块有利于物种多样性的形成。 图1 珠芽蓼斑块和对照区域植物物种多样性 注:*,P<0.05。 2.3地上生物量构成 珠芽蓼斑块群落地上总生物量、绿色物质量和死物质量均极显著高于对照区域的(P<0.001),其禾草生物量及其比例表现为珠芽蓼斑块显著小于对照(P<0.05),莎草和豆科植物生物量及其比例在珠芽蓼斑块和对照之间均无显著差异(P>0.05)。珠芽蓼斑块群落地上绿色生物量主要由杂类草和禾草植物构成,分别占绿色生物量的60.9%和22.1%,且珠芽蓼种群几乎占绿色生物量的1/3;而对照区域以禾草和杂类草植物为主,分别占绿色生物量的53.3%和27.1%。珠芽蓼斑块和对照区域草地类群生物量比例与其重要值构成变化规律类似。 2.4主要植物种种间关联度Jaccard指数 珠芽蓼斑块及其对照区域主要植物种种间Jaccard指数显示,珠芽蓼斑块中强联结种对数为21对,次强联结种对数为30对,弱联结种对数为4对,无联结种对数为0对,分别占总种对数的38.2%、54.5%、7.3%和0。对照中强联结种对数为9对,次强联结种对数为36对,弱联结种对数为23对,无联结种对数为23对,分别占总种数的9.9%、39.5%、25.3%和25.3%(表2)。说明,珠芽蓼斑块植物种的种间关联性强,群落处于稳定阶段。 图2 珠芽蓼斑块和对照区域的群落生物量构成 注:*,P<0.05; **,P<0.01; ***,P<0.001。 表2 珠芽蓼斑块主要植物种种间Jaccard指数(JI) 2.5主要植物种种间Spearman秩相关 Spearman秩相关分析显示,珠芽蓼斑块中,正相关的种对数为25对,其中极显著(P<0.01)和显著(P<0.05)相关的各两对,不显著(P>0.05)相关的21对;负相关的种对数为 30对,其中极显著相关的种两对,显著相关的3对,不显著相关的25对。对照区域,正相关的种对数53对,其中极显著相关的6对,显著相关的11对,不显著相关的36对;负相关的种对数38对,其中极显著相关的3对,显著相关的10对,不显著相关的25对。珠芽蓼斑块和对照区域中,显著关联的种对数分别占各自总种对数的16.4%和33.0%,其中显著、极显著正相关的种对比例分别为7.3%和18.7%,显著、极显著负相关的种对比例分别为9.1%和14.3%。珠芽蓼斑块的55个种对和对照区域的91个种对中,绝大多数种对的联结关系未达到显著水平,种间联结较为松散,种对间的独立性相对较强。珠芽蓼斑块中正联结种对占总种对数的45.45%,负联结种对占总种对数的54.55%,正负关联比为0.83;而对照区域中正联结种对占总种对数的58.24%,负联结种对占总种对数的41.76%,正负关联比为1.40。说明,珠芽蓼斑块中,多数植物种对间联结性较弱,种间独立性强,两个对照的植物种对间联结性较强,两个群落处于稳定阶段,这与主要植物种种间关联度Jaccard指数所得结果类似。 图3 珠芽蓼斑块和对照区域主要植物种Spearman秩相关分析半矩阵图 本研究珠芽蓼斑块主要植物种种间关联度Jaccard指数和Spearman秩相关整体结果均显示珠芽蓼斑块群落处于稳定阶段。一方面由于珠芽蓼斑块具绝对优势种,其植物种优势度构成模式更利于群落的稳定[16];另一方面由于珠芽蓼斑块中的珠芽蓼因富含单宁等物质,家畜不喜采食[12],牛羊仅在夏末秋初采食其花果[17],且珠芽蓼斑块中可食植物如禾草等比对照区域的低,从而放牧家畜对珠芽蓼斑块的整体采食程度低,利于其群落稳定性的维持。而对照区域优势种西北针茅和矮嵩草的优势度较低,无明显亚优势种,且因家畜采食程度高而更易发生演替。这进一步表明,珠芽蓼斑块中植物种对之间的竞争整体相对较弱,群落成熟程度较高,各物种间以相互独立的关系求得资源的充分利用,从而保持斑块物种间的稳定发展;而对照群落中植物种间的整体竞争相对激烈,群落处于演替初、中期阶段,群落容易发生演替变化。本研究珠芽蓼斑块群落的物种数和Jaccard关联度均高于对照区域,可能是因为珠芽蓼斑块内群落稳定性较强,群落内种间竞争比对照区域小,而利于物种多样性的形成。由此说明,家畜不喜食或采食程度低的植被斑块的形成更利于群落稳定性和物种多样性的形成。 植物种间正关联,主要是由于它们的生物学特性相近、对生境具有相似的生态适应性[18-19]。本研究的珠芽蓼斑块中,亚优势种球花蒿与赖草种对间呈显著正相关,它们均具耐旱、耐寒、耐贫瘠,生态适应性较强的特征,从而二者具有相似的生境和利用资源的特征;对照区域中优势种西北针茅、矮嵩草与其它植物无显著正相关关系。植物种间负关联,主要是由于生物学特性不同,对生境具有不同的生态适应性或相互竞争所致[20]。本研究珠芽蓼斑块中,优势种珠芽蓼与伴生种线叶嵩草、矮嵩草、甘肃棘豆之间的关联均呈显著负相关,是由于这些伴生种根系细弱入土较浅,对土壤水分和养分的吸收能力较弱,而珠芽蓼具粗壮肥厚根茎[14],对水分和养分吸收的竞争能力较强,且其总酚类和黄酮含量高,抗氧化能力强[21],从而珠芽蓼对环境的适应性更强,由此,珠芽蓼与这些植物种之间存在明显排斥性;对照区域的优势种西北针茅、矮嵩草与早熟禾、秦艽、兰花韭、喉毛花、唐松草种对间呈显著或极显著负相关,是由于西北针茅和矮嵩草与这些伴生种之间具有不同生境适应性和资源利用特征,相互之间的竞争或生态适应性不同,即呈现相互排斥。此外,本研究珠芽蓼斑块和对照区域正相关与负相关的植物种对数均存在,这与顾宁等[15]在山西芦芽山珠芽蓼群落植物种间关联整体呈正关联的研究结果存在分异,可能与其珠芽蓼群落无放牧利用有关。 References: [1]龙瑞军.青藏高原草地生态系统之服务功能.科技导报,2007,25(9):26-28. Long R J.Functions of ecosystem in the Tibetan grassland.Science & Technology Review,2007,25(9):26-28.(in Chinese) [2]李以康,林丽,张法伟,梁东营,王溪,曹广民.嵩草草甸退化和恢复过程中主要牧草演替和地表特征变化.草业学报,2010,19(5):179-185. Li L K,Lin L,Zhang F W,Liang D Y,Wang X,Cao G M.The main forage succession and ground surface characteristic changes during degradation and restoration of alpineKobresiameadow.Acta Prataculturae Sinica,2010,19(5):179-185.(in Chinese) [3]牟晓明,于应文,王先之,张红梅,徐长林,花立民.青藏高原火绒草斑块群落空间格局分析.生态学报,2015,36(16):5306-5315. Mou X M,Yu Y W,Wang X Z,Zhang H M,Xu C L,Hua L M.Analysis of community spatial patterns ofLeontopodiumnanumpatches in Qinghai-Tibetan Plateau.Acta Ecologica Sinica,2015,35(16):5306-5315.(in Chinese) [4]覃林.统计生态学.北京:中国林业出版社,2009:75-77. Tan L.Statistical Ecology.Beijing:China Forestry Publishing House,2009:75-77.(in Chinese) [5]张金屯.数量生态学.北京:科学出版社,2011:101-107. Zhang J T.Quantitative Ecology.Beijing:Science Press,2011:101-107.(in Chinese) [6]张东杰.青藏高原高寒草甸植被与土壤特征.草业科学,2015,32(2):269-273. Zhang D J.Comparison of vegetation and soil characteristics of Qinghai-Tibet plateau.Pratacultural Science,2015,32(2):269-273.(in Chinese) [7]石明明,张永超,张典业,任运涛,宗文杰,傅华,牛得草.高寒草甸草地微斑块植物特征及其土壤性质的研究.草业学报,2015,24(9):197-205. Shi M M,Zhang Y C,Zhang D Y,Ren Y T,Zong W J,Fu H,Niu D C.Plant traits and soil properties in pastures mini-patches in an alpine meadow.Acta Prataculturae Sinica,2015,24(9):197-205.(in Chinese) [8]牟晓明.青藏高原东北缘高寒草甸植物群落空间格局研究.兰州:兰州大学硕士学位论文,2015. Mou X M.The study on the spatial patterns of vegetation community in alpine meadow on the northeast Qinghai-Tibetan plateau.Master Thesis.Lanzhou:Lanzhou University,2015.(in Chinese) [9]牟晓明,于应文,张红梅,孙红,王虎成,徐长林,花立民.牦牛粪对高寒草甸植被群落特征和生态位参数的影响.草业科学,2013,30(10):1594-1601. Mou X M,Yu Y W,Zhang H M,Sun H,Wang H C,Xu C L,Hua L M.Effects of yak dung deposition on community characteristics and niche parameters in alpine meadow.Pratacultural Science,2013,30(10):1594-1601.(in Chinese) [10]张卫国,黄文冰,杨振宇.草地微斑块与草地退化关系的研究.草业学报,2003,12(3):44-50. Zhang W G,Huang W B,Yang Z Y.The study on the relationship between mini-patch and degradation of pasture.Acta Prataculturae Sinica,2003,12(3):44-50.(in Chinese) [11]才让吉,卫万荣,张卫国.草原鼠类与草食家畜食物生态位分异初探.草业科学,2015,32(5):802-808. Cairangji,Wei W R,Zhang W G.The primary investigation of food niche differentiation between livestock and grassland rodents.Pratacultural Science,2015,32(5):802-808.(in Chinese) [12]翁华.珠芽蓼珠芽药用和营养成分分析.江苏农业科学,2011,39(3):449-450. Weng H.Analysis on the bulbil medicinal values and nutrient constituents ofPolygonumviviparum.Jiangsu Agricultural Sciences,2011,39(3):449-450.(in Chinese) [13]易现峰,贲桂英.高寒草甸珠芽蓼抗氧化物质含量的季节变化及与太阳辐射的关系.西北植物学报,2000,20(2):201-205. Yi X F,Ben G Y.Seasonal variation in antioxidants ofPolygonumviviparumand its relation to solar radiation in alpine meadow.Acta Botanica Boreali-Occidentalia Sinica,2000,20(2):201-205.(in Chinese) [14]邹园.珠芽蓼的开花结实与胎生繁殖.雅安:四川农业大学硕士学位论文,2009. Zou Y.Study on flowering,fructification and viviparity ofPolygonumviviparumL.Master Thesis.Ya’an:Sichuan Agricultural University,2009.(in Chinese) [15]顾宁,张金屯,曹杨.芦芽山珠芽蓼群落种间关联的研究.山西大学学报:自然科学版,2007,30(3) :415-418. Gu N,Zhang J T,Cao Y.Study on interspecific association ofPolygonumviviparumcommunities in Luya Mountains.Journal of Shanxi University:Natural Science Edition,2007,30(3):415-418.(in Chinese) [16]张文,张建利,莫本田,王普昶.喀斯特山地草地植物群落物种优势度及优势种分析.安徽农业科学杂志,2011,39(29):17966-17969. Zhang W,Zhang J L,Mo B T,Wang P C.Analysis of the species dominance & dominant on the Karst mountain grassland plant community.Journal of Anhui Agricultural Science,2011,39(29):17966-17969.(in Chinese) [17]赵忠,韩天虎,王安禄.青藏高原东缘珠芽蓼+线叶嵩草草地植物群落组成及生产力动态监测.草原与草坪,2006(5):8-11. Zhao Z,Han T H,Wang A L.Dynamic monitoring of plant community composition and productivity ofPolygonumviviparumandKobresiacapillifoliarangeland on Qinghai-Tibetan Plateau.Grassland and Turf,2006(5):8-11.(in Chinese) [18]闫美芳,上官铁梁,张金屯,曹杨.五台山蓝花棘豆群落优势种群的种间关系分析.生态学杂志,2006,25(4):354-358. Yan M F,Shangguan T L,Zhang J T,Cao Y.Interspecific association and correlation of dominant species inOxytropiscoeruleacommunity in Wutai Mountains.Chinese Journal of Ecology,2006,25(4):354-358.(in Chinese) [19]汲文宪,陈丽华,郭峰,卞西陈,王鹏.北沟林场天然次生林主要乔木树种种间关系分析.东北林业大学学报,2012,40(9):10-14. Ji W X,Chen L H,Guo F,Bian X C,Wang P.Interspecific relationships of main tree species in a natural secondary forest in Beigou forest farm.Journal of Northeast Forestry University,2012,40(9):10-14.(in Chinese) [20]尹林克,李涛.塔里木河中下游地区荒漠河岸林群落种间关系分析.植物生态学报,2005,29(2):226-224. Yin L K,Li T.Interspecific relationship analysis of desert riparian forest plant communities in the middle and lower reaches of the Tarim River.Acta Phytoecologica Sinica,2005,29(2):226-224.(in Chinese) [21]廉美娜,朱玉环,徐长林,尚占环,龙瑞军.祁连山东段高寒草甸植物总酚和黄酮含量及抗氧化活性研究.西北植物学报,2012,32(12):2492-2497.Lian M N,Zhu Y H,Xu C L,Shang Z H,Long R J.Contents of total polyphenols and flavonodis from alpine plants from Qilianshan Mountains and their antioxidant activity.Acta Botanica Boreali-Occidentalia Sinica,2012,32(12):2492-2497.(in Chinese) (责任编辑武艳培) The vegetation composition and interspecific association of patches distributing inPolygonumviviparumalpine meadow on northeast Qinghai-Tibetan Plateau Wei Bin, Wang Ying, Guan Shi-qi, Gao Hong-juan, Kong Yang-yun, Yu Ying-wen (State Key Laboratory of Grassland Agro-ecosystems, College of Pastoral Agriculture Science and Technology, Lanzhou University, Lanzhou 730020, China) The vegetation composition and interspecific relationship reflect the interaction relationship of spatial distribution pattern among plant populations and communities. We analyzed the important value, diversity index of species, biomass composition, the Jaccard association index and spearman rank correlation coefficient of main species growing on patch inPolygonumviviparumalpine meadow (P.viviparumpatch) and control (CK), in order to reveal the ecological mechanism of succession process in Alpine meadow. The results indicated that number of species inP.viviparumpatch was bigeer than that in CK, and the change pattern of each group was similar between important value and biomass composition.P.viviparumpatch dominated mainly by forbs and grasses, accounting for 60.9% and 22.1% of the green biomass, respectively, andP.viviparumaccounted for about one third of the green biomass; while it dominated mainly by grasses and forbs, accounting for 53.3% and 27.1% of the green biomass, respectively. InP.viviparumpatch and CK, the significantly associated species-pairs accounted for 16.4% and 33.0% of their respective total species-pairs. The interspecific association of most species-pairs was not significant, and the independence between species-pairs was stronger. The ratios of negative and positive correlations were 0.83 and 1.40, respectively. Plant communities ofP.viviparumpatch was a stable stage. The formation ofP.viviparumpatch can strengthen community stable of alpine meadow. alpine meadow;Polygonumviviparumpatch; important value; biomass composition; Jaccard association index; Spearman rank correlation Yu Ying-wenE-mail:yuyw@lzu.edu.cn 10.11829/j.issn.1001-0629.2015-0646 2015-12-17接受日期:2016-04-02 兰州大学国家级大学生创新创业训练计划项目(201610730107);教育部“草地农业系统耦合与管理”创新团队项目 魏斌(1996-),男,甘肃永登人,在读本科生。E-mail:weib14@lzu.edu.cn 于应文(1969-),男,甘肃永登人,副教授,博士,主要从事草地生态学研究。E-mail:yuyw@lzu.edu.cn Q948.158 A 1001-0629(2016)8-1618-07 魏斌,王莹,关士琪,高红娟,孔杨云,于应文.青藏高原东北缘高寒草甸珠芽蓼斑块植被构成和种间关联.草业科学,2016,33(8):1618-1624. Wei B,Wang Y,Guan S Q,Gao H J,Kong Y Y,Yu Y W.The vegetation composition and interspecific association of patches distributing inPolygonumviviparumalpine meadow on northeast Qinghai-Tibetan Plateau.Pratacultural Science,2016,33(8):1618-1624.
2 结果与分析




3 讨论与结论
