根茎萌发与幼苗期的虉草水分生态幅
2016-09-27国志昌陈冰祥吴海英
国志昌, 曹 昀,2*, 陈冰祥, 吴海英, 郑 祥
( 1. 江西师范大学 地理与环境学院, 南昌330022; 2. 江西师范大学 鄱阳湖湿地与流域研究教育部重点实验室, 南昌 330022; 3. 江西师范大学 生命科学学院, 南昌 330022 )
根茎萌发与幼苗期的虉草水分生态幅
国志昌1, 曹昀1,2*, 陈冰祥1, 吴海英1, 郑祥3
( 1. 江西师范大学 地理与环境学院, 南昌330022; 2. 江西师范大学 鄱阳湖湿地与流域研究教育部重点实验室, 南昌 330022; 3. 江西师范大学 生命科学学院, 南昌 330022 )
为揭示虉草根茎萌发和幼苗期的水分生态幅,该研究通过盆栽控制试验,设置8个水分梯度:(2 ± 0.5)%、(10 ± 0.5)%、(15 ± 0.5)%、(20 ± 0.5)%、(25 ± 0.5)%、(30 ± 0.5)%、(40 ± 0.5)%(水分饱和)和淹水(2 cm),通过萌发率、各形态指标和生理指标对虉草水分生态幅进行了探讨。结果表明:(1)土壤水分含量过低和过高都会降低虉草根茎萌发率。(2)随土壤水分含量的增加,虉草幼苗株高和地上生物量都呈现先上升后下降的趋势,两者并呈指数函数关系y= 0.203 8e0.048 6x(R2= 0.803)。(3)虉草通过调整自身结构特点,降低个体大小、减少叶片数量和叶面积来适应胁迫环境,土壤水分含量为10%~30%时虉草含水率能保持相对稳定的水平。虉草的光和色素(Chl a、Chl b、Car)和相对叶绿素(SPAD)含量均随土壤水分含量的增加呈现出先上升后下降的趋势,Chl a/b则相反,呈先下降后上升趋势。(4)Chl/Car各组之间差异不显著。综合上述研究,利用高斯模型分析得出虉草的水分生态幅为7.48%~52.20%,最适水分生态幅为18.99%~40.60%。
虉草, 土壤水分含量, 根茎, 萌发, 幼苗期, 水分生态幅
土壤水分作为植物生长的主要环境因子(刘祖贵等,2006),极端的土壤水分条件(干旱和淹水)会影响植物的形态结构和生理过程,如生物量、株高、叶片特征、光合作用、气孔行为,叶绿素荧光合代谢物积累(Ohashi et al,2006)。此外湿地植被的分布也与土壤含水量密切相关(刘永等,2006; Castelli et al,2000),枯水期与丰水期的交替和局部气候的变化使湿地水文条件复杂,土壤水分含量变化较大,因此水分因子对湿地植物变得尤为重要。
虉草(Phalarisarundinacea)以营养繁殖为主,为禾本科(Gramineae)虉草属(Phalaris)多年生草本植物,杆较粗壮,高为60~150 cm,叶片宽为5~15 mm,分布在我国东北、华北、华中、江苏、浙江等地,全球温带地区广布,多生于水湿处,对环境要求不高,具有比较广的地理分布和生境范围(中国科学院北京植物研究所,1979; Martina & Ende,2012)。有关虉草的研究主要集中在盐分胁迫(丛丽丽等,2011;张永亮等,2014)、种子破眠与萌发(张浩等,2012; 徐高峰等,2011; 斯日古楞等,2014)以及饲料青贮(李平等,2013; 李平等,2014)等方面。但有关土壤水分含量对虉草影响的研究较少见有报道,仅宋家壮等(2012)研究了水分胁迫对虉草生理生化特性的影响。
本研究通过水分控制试验,分析土壤水分含量对虉草根茎萌发、幼苗生理和形态的影响,探求其水分生态幅和最适土壤水分含量。有利于理解虉草的自然分布和对土壤水分变化的适应性,以揭示不同繁殖方式及幼苗生长对不同土壤水分含量的适应机制,为退化湿地的生态治理提供参考。
1 材料与方法
1.1 材料
试验于2015年4月在阳光培养室进行,共计60 d。虉草根状茎取自鄱阳湖湿地(116.32° E,28.9° N),选取大小均一的根状茎为试材,根据芽的位置切4~5 cm,栽培到塑料盆(长35 cm,宽26 cm,高13 cm)内,每盆装土8.5 kg,埋深为10 cm。培养基质为砂壤土。基本性状:pH 5.25,有机质24.64 g·kg-1,全氮1.42 g·kg-1,土壤田间持水量为30.76%。
1.2 试验处理
试验共设置8个水分梯度:C1(2 ± 0.5)%、C2(10 ± 0.5)%、C3(15 ± 0.5)%、C4(20 ± 0.5)%、C5(25 ± 0.5)%、C6(30 ± 0.5)%、C7饱和(40 ± 0.5)%和C8淹水(2 cm),设3个重复。使用HH2型土壤水分仪(Moisture Meter),每天19:00-20:00进行测定并补水,根据所测土壤体积含水量求出所需水分体积,用量筒量取所需水补充至设置值。
1.3 测定指标和方法
每天计算萌发率、萌发速率指数(GR)。每两天测量幼苗株高,计算绝对增长速率(HAGR);测量叶长、叶宽,计算叶片面积。试验结束(60 d)测量各处理组叶长、叶宽和单株地上生物量(湿重)。将其70 ℃烘干,测量地上生物量(干重),计算含水率。采用Arnon法测定叶绿素和类胡萝卜素含量,SPAD-502叶绿素仪测定相对叶绿素SPAD值。
1.4 数据处理和模型
采用Excel进行数据分析,用SPSS软件进行相关性分析和单因素方差分析,并运用LSD 检验法对不同处理组的各生理指标进行差异性分析比较。运用高斯模型计算水分生态幅和最适水分生态幅。高斯模型方程:y=cexp[-0.5(x-u)2/t2]。
2 结果与分析
2.1 土壤水分含量对虉草株高及根茎萌发率的影响
萌发试验中,C1组无萌发,C2~C8组1~15 d萌发率变化较快,15 d后各组萌发率保持稳定,其中C8组与其它各组存在极显著差异(P<0.01)(图1)。株高1~30 d增长较快,30 d后增长减慢,各处理组生长趋势一致。其中C2、C8与C4、C5组之间显著极差异(P<0.01),C2、C8与C3、C6组之间C7与C4、C5之间差异显著(P<0.05),C3、C4、C5组间差异不明显(图2)。由于除了C1、C8外各处理组萌发均集中在15 d内,所以选取前期(1~20 d)的萌发速率指数进行分析,结果显示随着土壤水分含量的增加,虉草根茎萌发率和1~20 d的萌发速率指数(GR)均先升高后降低(图3)。C1,C8(淹水)组虉草根茎萌发率为0和45.8%。C5,C7萌发率为95.8%和87.5%,C3、C4、C6萌发率达100%。水分含量在10%以下和40%以上都会抑制虉草根茎的萌发。1~20 d的萌发速率指数值,C5处理组最高为1.81。除去C1组整体上看GR值,C2与C6差异显著(P<0.05),C5、C7、C8与其他各种都存在极显著差异(P<0.01)。这说明极端土壤水分条件影响了虉草根茎萌发率与萌发速率指数。
土壤水分含量在15%~30%之间时虉草幼苗长势较好,其中C5组虉草长势最好,平均株高为61.86 cm。C2组萌发率达96%,但C2、C8组幼苗长势较差,株高为43.25 cm、41.92 cm与C3~C6组差异较大。在试验的3个时期,虉草幼苗株高的绝对增长速率HAGR差异较大,随着生长时期的延长绝对增长速率HAGR呈逐渐变小趋势,40 d之后增长速率明显减小但仍在生长。1~20 d绝对增长速率最大,各处理组之间绝对增长速率随土壤水分含量增加呈先升高后降低的趋势(图4)。土壤含水量与萌发率,株高数据进行二次拟合,萌发率拟合曲线R2=0.828,株高拟合曲线R2=0.824,达到高斯模型要求。
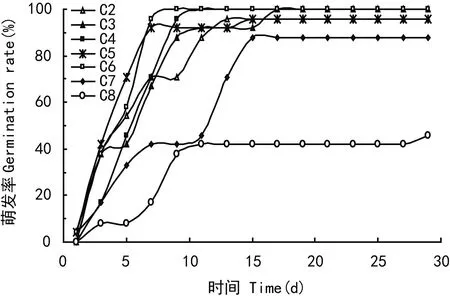
图 1 萌发率随时间的变化 Fig. 1 Temporal variation of germination rate of Phalaris arundinacea
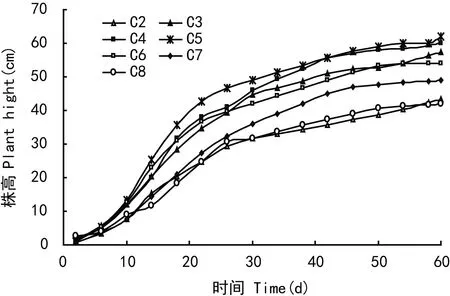
图 2 幼苗的生长随时间的变化Fig. 2 Temporal variation of plant height of the young Phalaris arundinacea shoots
2.2 土壤水分含量对虉草叶片形态学特征的影响
虉草幼苗叶长、叶宽、叶片面积与叶片数各处理组均随土壤水分含量的增加先上升后下降。C4、C5、C6叶片各特征值相对较大,长势较好。对各特征值进行二次拟合,拟合程度均较好,达到高斯模型要求(图5)。极度干旱(C1)、饱和(C7)和淹水(C8)组叶片形态特征值与其它组呈极显著差异(P<0.01),说明极端的土壤水分条件(干旱和淹水)对虉草叶片生长有一定影响。
2.3 土壤水分含量对虉草光合色素含量的影响
虉草幼苗叶绿素和相对叶绿素含量变化趋势一致,随土壤水分含量的增加先升高后降低。Chl a/b随土壤水分含量的增加呈现出先下降后上升的趋势,土壤水分含量为10%、40%和淹水时Chl a/b明显大于其他处理组。类胡萝卜素 (Car) 各组间差异不大,Car与Chl/Car各处理组间无明显变化趋势(图6)。
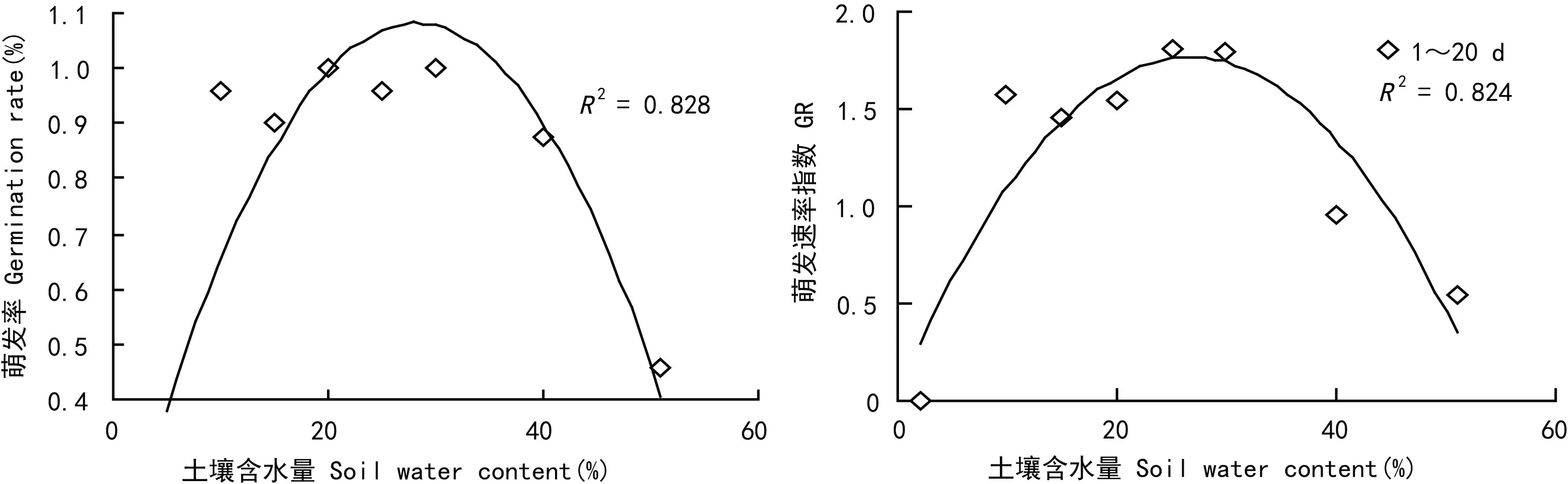
图 3 土壤水分含量对根茎萌发和萌发速率指数的影响Fig. 3 Effects of soil moisture content on rhizomatic germination and germination rate of Phalaris arundinacea
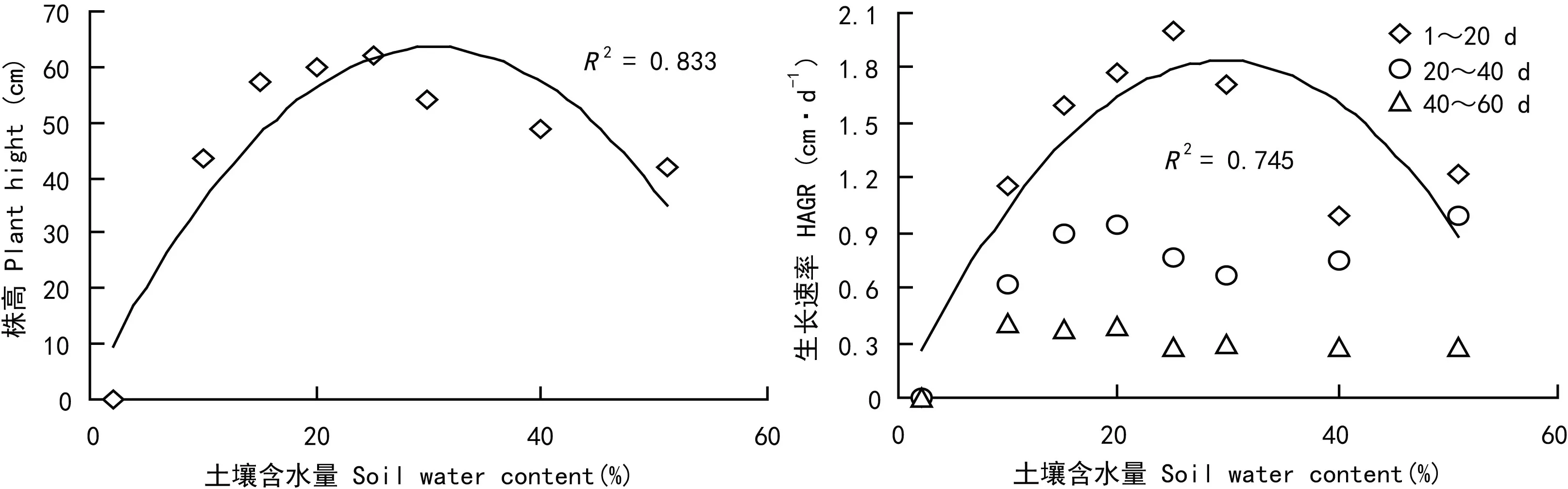
图 4 土壤水分含量对虉草株高和生长速率的影响Fig. 4 Effects of soil moisture content on height and HAGR of Phalaris arundinacea
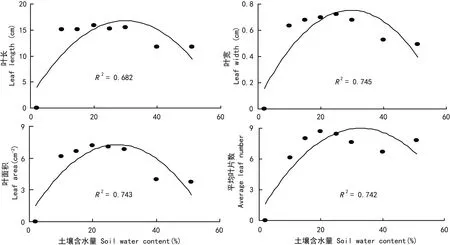
图 5 土壤水分对虉草叶片形态特征的影响Fig. 5 Effects of soil moisture content on leaf properties of Phalaris arundinacea
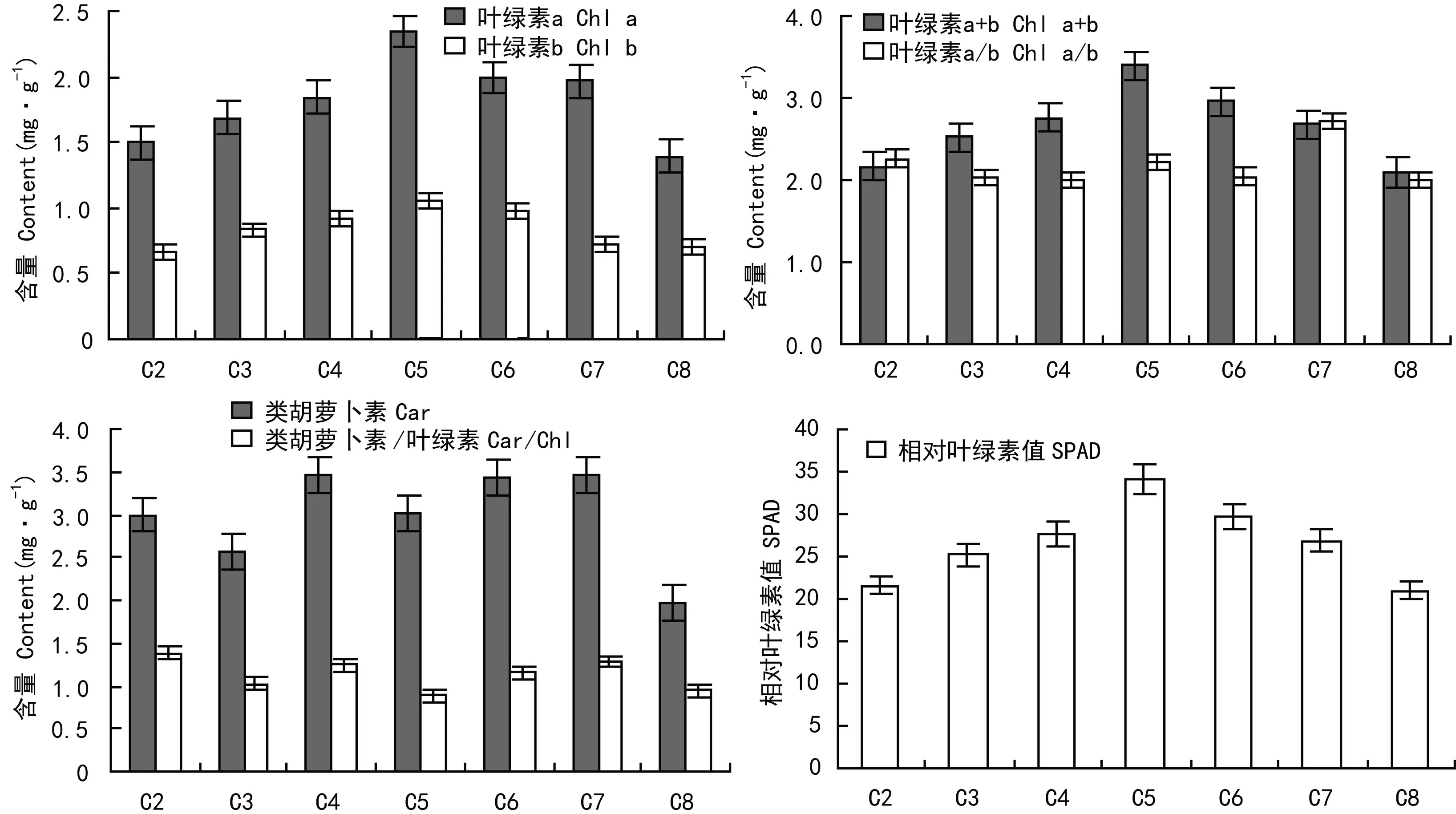
图 6 土壤水分对虉草光合色素含量的影响Fig. 6 Effects of soil moisture content on photosynthetic pigments contents of Phalaris arundinacea
2.4 土壤水分含量对虉草地上生物量和含水率的影响
虉草幼苗地上生物量在土壤水分含量为15.5%~30.5%时较高,C4组最大达到3.99 g,C8组最小仅1.33 g,整体呈先升高后降低趋势(图7)。虉草幼苗含水率在土壤水分含量10%~30%之间相对稳定,土壤水分含量大于30%时虉草幼苗含水率随土壤水分含量的增加而增大(图8)。对虉草幼苗各特征值进行相关性分析,各特征值间相关系数在0.75~0.98之间,萌发率与地上生物量、叶片数,地上生物量与叶片数、Chl a、Car,Car与 Chl b在P<0.05水平存在显著相关,其余各特征之间均在P<0.01水平上显著相关(表2)。根据各生物特征的相关分析,株高与生物量之间显著相关(P<0. 01)并呈指数函数关系y= 0.203 8e0.0486x(R2= 0.803),由此得出虉草幼苗生物量的时间变化(图9)。
2.5 虉草水分生态幅分析
将萌发率与各形态和生理指标进行自然对数转换,二次多项式函数拟合,再根据拟合函数得出高斯回归方程(表2)。求出各指标对土壤水分含量的耐受度t,最适点u和指标最大值,进而得出耐受范围和最适土壤水分含量(表3)。综合各指标取生态幅和最适生态幅的交集,得出虉草生长的最适水分含量值在18.99%~40.60%处,极限水分为7.48%~52.20%,求出的最适生态幅可以很好地反映虉草生长需要的最适土壤水分含量。
3 讨论与结论
3.1 虉草根茎萌发与形态和生理特征对不同土壤水分含量的响应
植物萌发与生长直接受到土壤水分含量的影响。本研究发现,随土壤水分含量增加虉草根茎萌发率呈现先升高后降低的趋势,土壤水分含量为2%时受到干旱胁迫,全部死亡未萌发,土壤水分含量为10%时, 萌发率为96%, 其趋势与奇异虉草和小子虉草种子萌发的研究结果相一致,但根茎萌发率高于种子萌发率(徐高峰等,2011)。土壤水分处于饱和状(40%)和淹水状态时萌发率为87.5%和45.8%。随土壤水分含量增加1~20 d的萌发速率指数(GR)先升高后下降,并且各处理组之间差异性显著,说明虉草根茎的萌发和萌发速率受极端土壤水分状况的影响。
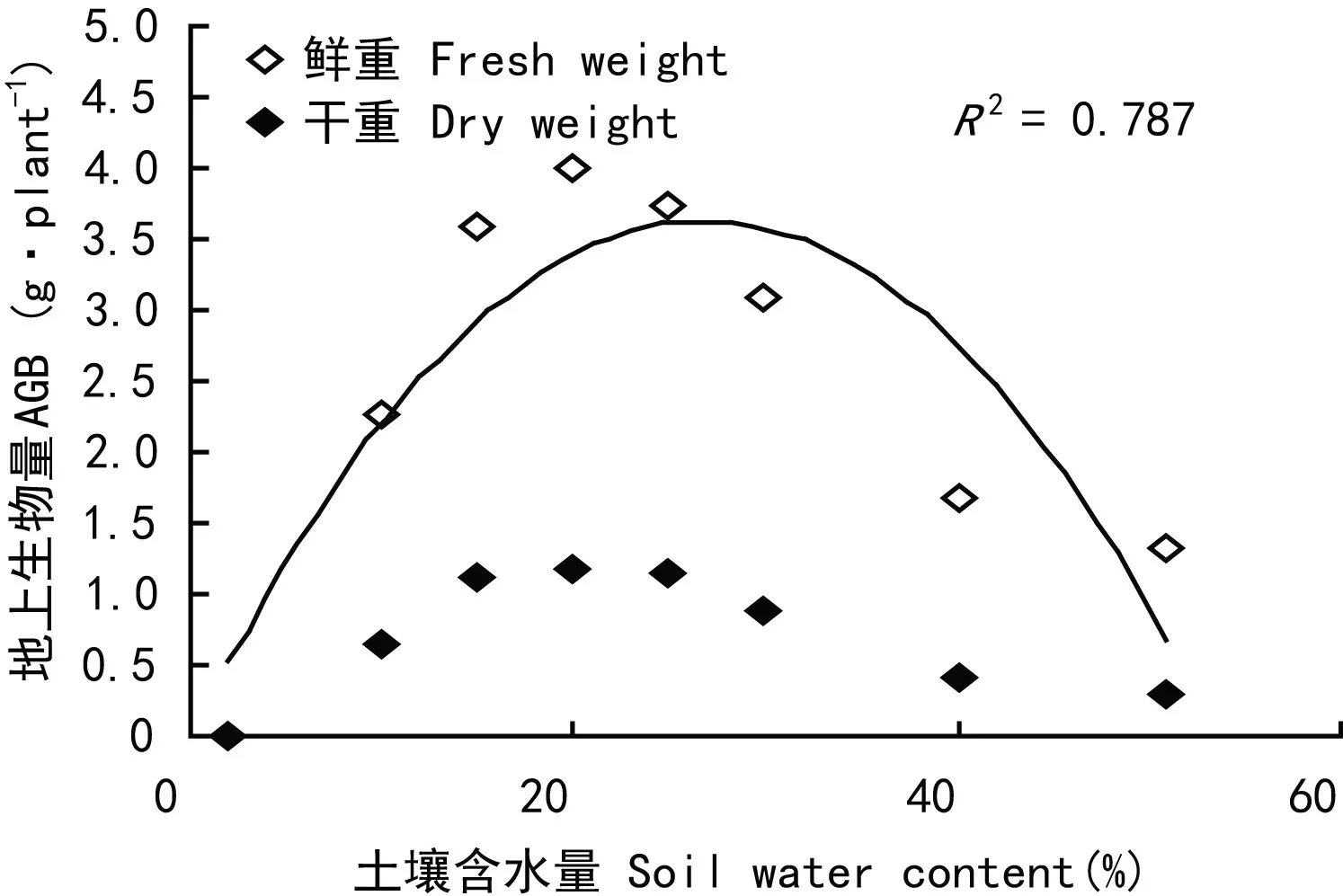
图 7 土壤水分对虉草地上生物量的影响Fig. 7 Effects of soil water content on aboveground biomass of Phalaris arundinacea
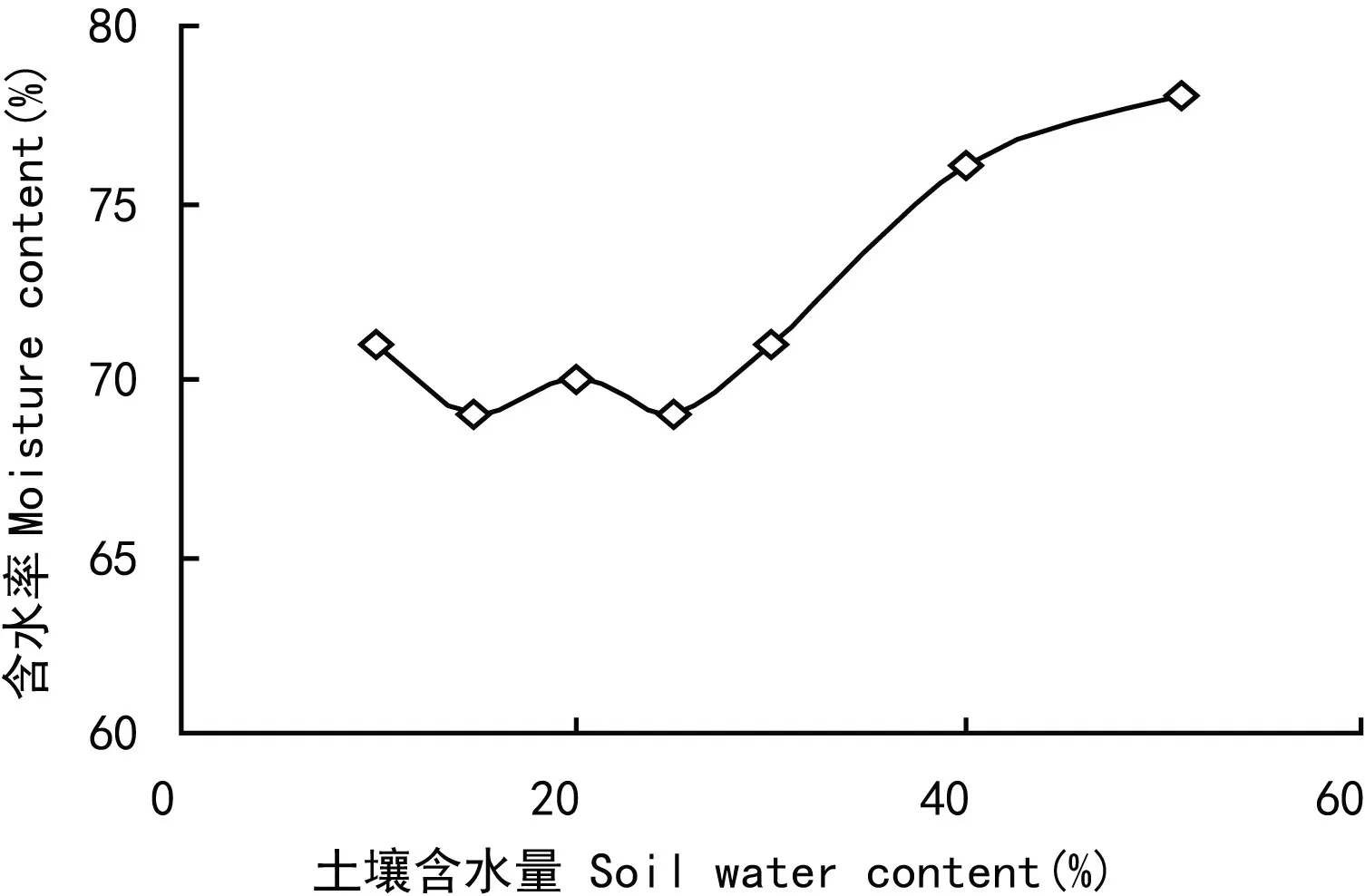
图 8 土壤水分对虉草含水率的影响Fig. 8 Effects of soil water content on moisture rate of Phalaris arundinacea
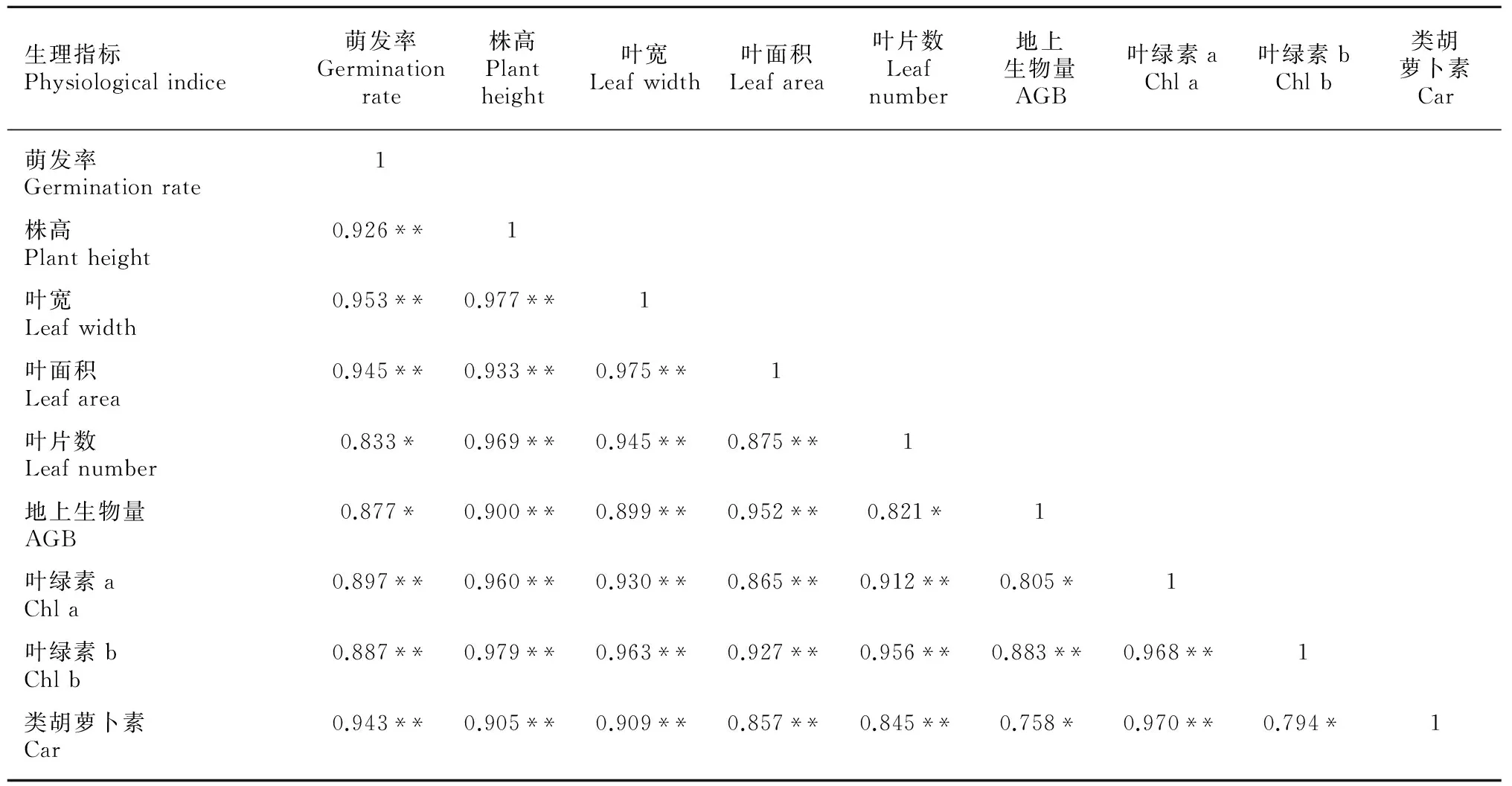
表 1 虉草形态和生理指标的相关性
注: * 表示在0.05水平(双侧)上显著相关,** 表示在0.01水平(双侧)上显著相关。
Note:* means significant difference atP<0.05,** means significant difference atP<0.01(bilateral).
土壤水分含量在15%~30%时幼苗生长良好,土壤水分含量为25%时株高最高达到61.86 cm,当低于10%高于40%时株高为43.25 cm、41.92 cm。随着土壤水分含量的增加虉草幼苗的株高和绝对增长速率(HAGR)都呈现先上升后下降的趋势,同一水分处理下绝对增长速率(HAGR)随时间呈现逐渐降低的趋势,与灰化苔草研究结果相一致(杨阳等,2015)。这说明在干旱或土壤水分过饱和的条件下虉草的株高和绝对增长速率 (HAGR) 会受到影响。
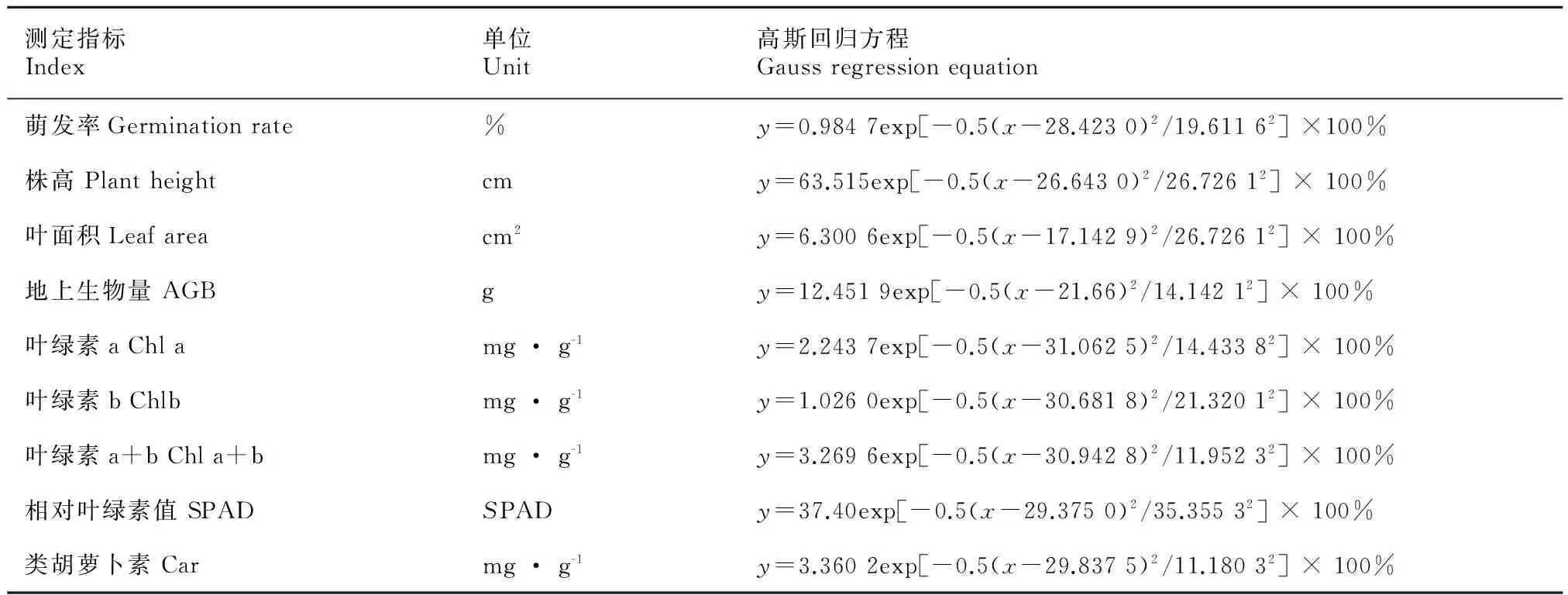
表 2 虉草形态和生理指标的高斯回归方程
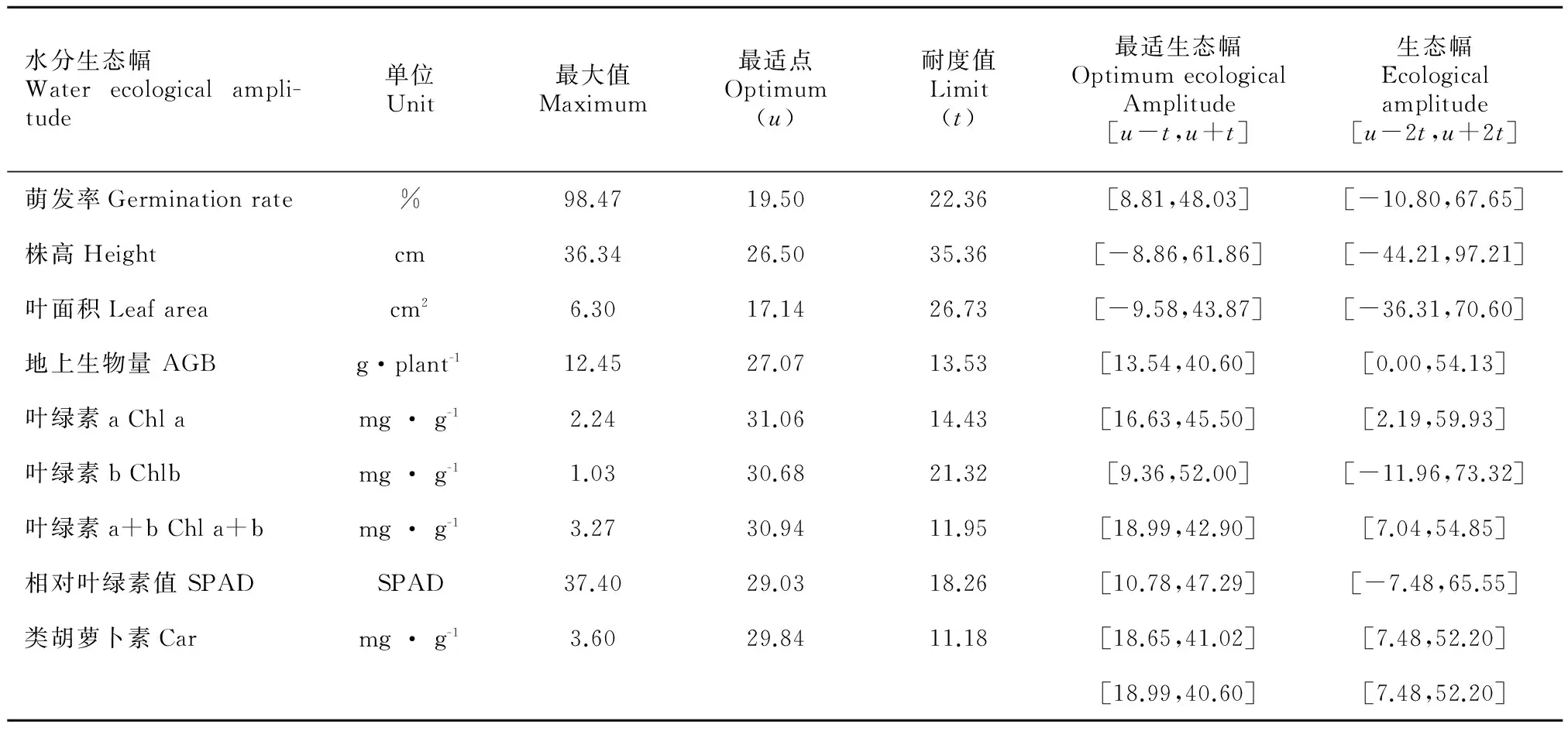
表 3 虉草的水分生态幅

图 9 虉草地上生物量的时间变化Fig. 9 Temporal variation of simulated aboveground biomass of Phalaris arundinaceas
叶片是保证植株存活的重要器官。当生长环境发生变化时,植物会在形态和生物量分配上做出响应(Gianoli et al,2005; 黄迎新等,2008)。在极端土壤水分含量下植物会通过减小叶面积和生物量适应环境(Roux et al,1998; 曹昀等,2007)。本研究发现,随土壤含水量增加形态学特征和生物量呈现先增高后降低的趋势,在干旱环境中含水率保持相对稳定,说明虉草在干旱和水分过饱和的土壤水分条件下通过调整自身结构特点,降低个体大小、减少叶片数量和叶面积来适应胁迫环境,具有一定的抗旱性。
光合色素含量可以影响植物光合效率和植株生长 (王忠,2002)。叶绿素a/b表示植物中捕光色素蛋白复合物的含量,叶绿素a/b升高,蛋白复合物的含量降低(Thornber,2003)。本研究发现极端土壤水分状况会抑制虉草光合色素合成,使光合色素含量降低。本研究中Chl a/b随土壤含水量的增加呈现出先下降后上升的趋势,土壤水分含量为10%、40%和淹水时Chl a/b明显大于其他处理组,说明虉草为适应极端的土壤水分条件(干旱和淹水)降低了光能捕获量以保证光合机构免于遭受光氧化破坏。类胡萝卜素(Car)能清除活性氧,防止膜脂过氧化(米海莉等,2004)。本研究中类胡萝卜素在各土壤水分含量下较稳定,在光合色素中的比值也差异不大,有利于平衡光合生理活动,保证个体存活。
3.2 虉草幼苗的水分生态幅
本研究得出虉草的水分生态幅为7.48%~52.20%,最适水分生态幅为18.99%~40.60%,超过水分生态幅的上限和下限虉草均不能存活。其中,根茎萌发的最适水分含量为8.81%~48.03%,水分含量范围较大,说明虉草根茎具有比较强的耐受性,与种子相比采用根状茎作为繁殖体进行繁殖具有更强抗旱性。地上生物量与叶绿素含量对土壤水分含量的响应较明显,最适范围为13.54%~40.60%和18.99%~42.90%,说明极端的土壤水分条件对光合作用的抑制较大进而限制了植株生物量的积累。确定虉草的水分生态幅和最适土壤水分含量,对进一步研究虉草的潜在分布与水文波动下湿地的群落演替和生态系统的恢复和保护具有重要意义。
CAO Y,WANG GX,LIU Y,2007. Effects of flooding on germination and seedlings growth of sweet flag(AcoruscalamusL. ) [J]. J Lake Sci, 19(5) :577-584. [曹昀,王国祥,刘玉,2007. 淹水对菖蒲萌发及幼苗生长的影响[J]. 湖泊科学, 05:577-584.]
CASTELLI RM,CHAMBERS JC,TAUSCH RJ,2000. Soil-plant relations along a soil-water gradient in Great basin riparian meadows[J]. Wetlands, 20( 2) : 251-266.
CONG LL, ZHANG YW, CHEN X, et al, 2011. Effect of NaCl stress on seed germination and physiological and biochemical characteristics of seedlings ofPhalarisarundinacea[J]. Prat Sci, (12):2 136-2 142. [丛丽丽,张蕴薇,陈新,等, 2011. 盐胁迫对虉草种子萌发与幼苗生理生化的影响[J]. 草业科学, 12:2 136-2 142.]
GIANOLI E , GONZALEZ -TEUBER M, 2005. Environmental heterogeneity and population differentiation in plasticity to drought inConvolvuluschilensis(Convolvulaceae)[J]. Evol Ecol, 19(6):603-613.
HUANG YX, ZHAO XY, ZHANG HX, et al, 2008. Responses of agriophyllum squarrosum phenotypic plasticity to the changes of soil nutrient and moisture contents and population density [J]. Chin J Appl Ecol, (12):2 593-2 598. [黄迎新,赵学勇,张洪轩,等, 2008. 沙米表型可塑性对土壤养分、水分和种群密度变化的响应[J]. 应用生态学报, 12:2 593-2 598.]
INSTITUE OF BOTANY, THE CHINESE ACADEMY OF SCIENCE,1997. Iconographia gormophytorum sinicorum tomus V[M]. Beijing: Science Press:100. [中国科学院北京植物研究所,1976.中国高等植物图鉴(第5册)[M]. 北京:科学出版社:100.]
LI P, BAI SH, YOU MH, et al,2013. Effects of different additives on the silage quality ofPhalarisarundinaceaL. with low water content [J]. Prat & Anim Husb, (6):1-5.[李平,白史且,游明鸿,等, 2013.不同添加剂对低水分虉草青贮品质的影响[J]. 草业与畜牧, 6:1-5.]LI P, YAN JJ, BAI SH, et al, 2012. Preliminary study on silage ofElymussibiricnsL. andPhalarisarundinaceaL. in Alpine pastoral regions of North Sichuan[J]. Acta Agrestia Sin, (2):368-372.[李平,鄢家俊,白史且,等, 2012. 川西北高寒牧区老芒麦和虉草青贮效果初步研究[J]. 草地学报, 2:368-372.]
LIU Y, GUO HC,ZHOU F, et al,2006. Role of water level fluctuation on aquatic vegetation in lakes [J]. Acta Ecol Sin, (9):3 117-3 126. [刘永,郭怀成,周丰,等, 2006. 湖泊水位变动对水生植被的影响机理及其调控方法[J]. 生态学报,第9期(9):3 117-3 126.]
LIU ZG, CHEN JP, DUAN AW, et al, 2006. Effects of different soil moisture treatments on physiological characteristics of summer maize leaves [J]. Agric Res Arid Areas, 24(1):90-95.[刘祖贵,陈金平,段爱旺, 2006.不同土壤水分处理对夏玉米叶片光合等生理特性的影响[J]. 干旱地区农业研究, 24(1):90-95.]
MARTIN JP, ENDE CNV,2012. Highly plastic response in morphological and physiological traits to light, soil-N and moisture in the model invasive plant,Phalarisarundinacea[J]. Environ Exp Bot, 82(5):43-53.
MI HL, XU X, LI SH, et al, 2004. Effect of Soil water stress on contents of chlorophyll, soluble sugar, starch, C/N of two desert plants (CynanchumkomaroviiandGlycyrrhizauralensis)[J]. Acta Bot Boreal-Occident Sin, (10):1 816-1 821.[米海莉,许兴,李树华,等, 2004.水分胁迫对牛心朴子、甘草叶片色素、可溶性糖、淀粉含量及碳氮比的影响[J]. 西北植物学报, 10:1 816-1 821.]
OHASHI Y, NAKAYAMA N, SANEOKA H, et al, 2006. Effects of drought stress on photosynthetic gas exchange, chlorophyll fluorescence and stem diameter of soybean plants [J]. Biol Plant, 50(1):138-141.
ROUX X L, BARIAC T,1998. Seasonal variations in soil, grass and shrub water status in a West African humid savanna.[J]. Oecologia, 113(4):456-466.
(Continueonpage979)(Continuefrompage1007)
SI RGL, WANG MJ, LIU XJ, et al, 2014. Seed dormancy mechanism and breaking techniques of two varieties ofPhalarisarundinaceaL. [J]. J NE For Univ, (05):47-51.[斯日古楞,王明玖,刘雪娇,等. 2014.2个品种虉草种子破眠方法的比较[J]. 东北林业大学学报, 05:47-51.]
SONG JZ, LI PP, FU WG, 2012. Effect of water stress and rewatering and physiological and biochemical characteristics ofPhalarisarundinacea[J]. Acta Prat Sin, 21(2):62-69.[宋家壮,李萍萍,付为国, 2012.水分胁迫及复水对虉草生理生化特性的影响[J]. 草业学报, 02:62-69.]
THORNBER JP,2003. Chlorophyll-proteins: Light-harvesting and reaction center components of plants[J]. Ann Rev Plant Physiol, 26(4):127-158.
WANG Z,2002. Plant physiology[M]. Beijing: China Agriculture Press: 128-131.[王忠, 2002.植物生理学[M].北京:中国农业出版社:128-131.]
XIU GF, ZHANG FD, LI TL, et al, 2011. Influence of environmental factors on seed germination ofPhalarisparadoxaandPhalarisminor[J]. Acta Bot Boreal-Occident Sin, 31(7):1 458-1 465.[徐高峰,张付斗,李天林,等, 2011.环境因子对奇异虉草和小子虉草种子萌发的影响[J]. 西北植物学报, 31(7):1 458-1 465.]
YANG Y, CAO Y, ZHANG SJ, 2015. Effects of soil moisture regime on rhizomatic germination and young shoot growth ofCarexcinerascens. [J]. J Ecol Rur Environ, 31(2):180-187.[杨阳,曹昀,张素娟, 2015.水分条件对灰化苔草根茎萌发及幼苗生长的影响[J]. 生态与农村环境学报, 02:180-187.]
ZHANG H, ZHANG YL, GAO K,2012. Preliminary study on the dormancy mechanism and dormancy breaking metholds inPhalarisarundinaceaseeds [J]. Chin J Grassl, 34(6):7-12.[张浩,张永亮,高凯, 2012.虉草种子休眠机理与休眠破除方法初探[J]. 中国草地学报, 6:7-12.]
ZHANG YL, WANG XY, LIU Y, et al, 2014. Effect of different NPK fertilizer rates on the ion absorption and distribution of reed Canarygrass (Phalarisarundinacea) under salt stress [J]. Acta Agr Sin,(6):1 318-1 323.[张永亮,王学愿,刘杨,等, 2014.盐胁迫下氮磷钾肥对虉草离子吸收与分布的影响[J]. 草地学报, 6:1 318-1 323.]
Water ecological amplitude ofPhalarisarundinaceaat rhizomatic germination and seedling stage
GUO Zhi-Chang1, CAO Yun1,2*, CHEN Bing-Xiang1, WU Hai-Ying1, ZHENG Xiang3
( 1.CollegeofGeographyandEnvironment,JiangxiNormalUniversity, Nanchang 330022, China; 2.KeyLaboratoryofPoyangLakeWetlandandWatershedResearch,MinistryofEducation,JiangxiNormalUniversity, Nanchang 330022, China; 3.CollegeofLifeSciences,JiangxiNormalUniversity, Nanchang 330022, China )
We carried out an indoor culturing experiment to cultivatePhalarisarundinaceausing rhizome to explore water ecological amplitude at rhizomatic germination and seedling stage. We exposedP.arundinaceato C1(2 ± 0.5)%, C2(10 ± 0.5)%, C3(15 ± 0.5)%, C4(20 ± 0.5)%, C5(25 ± 0.5)%, C6(30 ± 0.5)%, C7(40 ± 0.5)% (water saturation) and C8 flooding(2 cm) for 60 d, and determined the germination rate and various physiological indexes and morphological indexes. The results were as followes: (1) Rhizomatic germination rate of the plants was lowered by drought or excessive saturated of soil water content, but the rhizome still had stronger drought-resistance than seeds. Plant height and aboveground biomass increase firstly and then decreased with the increase of soil water content. The relationship between the plant height and aboveground biomass could be described as an exponential function:y= 0.203 8e0.0486x(R2= 0.803 2). (2) The plants could adapt to the stress environment by adjusting the structural features, reducing the size, and decreasing the leaf number and the leaf area. Leaf length, leaf width, leaf number and leaf area showed the similar trend of increasing firstly and then decreasing with the increase of soil water content. When the soil water content ranged between 10%-30%, the water content of plant were able to maintain stability, and then increased with the increase of soil water content. (3) Chl a, Chl b, Car and SPAD contents in young shoots followed the same trend in variation, increasing firstly and then decreasing with the increase of soil water content. But Chl a/b content decreased firstly and then increased, did no differ much in the eight treatments. (4) There is no significant difference of Chl/Car in these treatments. In drought or excessive saturated of soil water content, the plants adjusted the distribution of Chl a and Chl b, raised Chl a /b, maintained Car /Chl, in order to maintain the regular photosynthesis process. Based on the above analysis, quantitive analysis on rhizomatic germination rate, morphological indexes and physiologic indexes by Gaussian Model, it is clear that the water ecological amplitude ofP.arundinaceafor rhizomatic germination and seedling stage was 7.48%-52.20%, and the optimum water ecological amplitude was 18.99%-40.60% .
Phalarisarundinacea, soil water content, rhizome, germination, seedling stage, water ecological amplitude
10.11931/guihaia.gxzw201601016国志昌, 曹昀, 陈冰祥, 等. 根茎萌发与幼苗期的虉草水分生态幅 [J]. 广西植物, 2016, 36(8):1000-1007
GUO ZC, CAO Y, CHEN BX, et al. Water ecological amplitude ofPhalarisarundinaceaat rhizomatic germination and seedling stage [J]. Guihaia, 2016, 36(8):1000-1007
2016-01-12
2016-05-03
国家自然科学基金 (41361017); 江西省自然科学基金 (20142BAB204006) [Supported by the National Natural Science Foundation of China (41361017); Natural Science Foundation of Jiangxi (20142BAB204006)] 。
国志昌(1990-),男,山东济宁人,硕士研究生,主要从事恢复生态学研究,(E-mail)xiaogui3009@163.com。
曹昀,博士,副教授,从事植物生态学研究,(E-mail)yun.cao@163.com。
Q948
A
1000-3142(2016)08-1000-08
