地锦营养器官及乳汁管的解剖研究
2016-09-26李双江焦铸锦许笑蒙闫光兰阚云超李春奇
李双江,焦铸锦,许笑蒙,闫光兰,阚云超,李春奇
(1.河南农业大学生命科学学院,河南 郑州 450002; 2.南阳师范学院,河南省伏牛山昆虫生物学重点实验室,河南 南阳 473061)
地锦营养器官及乳汁管的解剖研究
李双江1,焦铸锦2,许笑蒙1,闫光兰2,阚云超2,李春奇1
(1.河南农业大学生命科学学院,河南 郑州 450002; 2.南阳师范学院,河南省伏牛山昆虫生物学重点实验室,河南 南阳 473061)
通过组织切片和显微观察法,研究了地锦营养器官的解剖特征及乳汁管的分布和发育过程。结果表明,地锦根的初生木质部为三原型;次生结构由周皮、次生韧皮部、维管形成层和次生木质部构成,维根比约72.4%。茎的初生结构由表皮、皮层、维管束和髓构成,次生结构中无周皮。叶为异面叶,上表皮气孔比下表皮多,叶肉中具有与碳4(C4)植物相同的花环状结构。乳汁管主要分布在主根、侧根和茎的韧皮部外侧以及叶肉内,为无节乳汁管,乳汁管中有较多颗粒状晶体物质。成熟的乳汁管直径差异较大,由大到小依次为茎﹥主根﹥侧根﹥叶。
地锦;营养器官;解剖结构;乳汁管
地锦(EuphorbiahumifusaWilld)俗称铺地锦,大戟科(Euphorbiaceae)大戟属(EuphorbiaL.)地锦草组,一年生草本植物[1],是重要的药用植物。因含有鞣质、黄酮、甾醇类化合物,临床上具有抗菌消炎、清热解毒、利湿退黄、凉血止血等功能,常用于肠炎、病毒性肝炎、出血症(咳血、尿血、便血)、痢疾、皮肤病等疾病的治疗[2]。目前,对地锦药用价值已有广泛报道[3],但是有关地锦营养器官的结构及乳汁管的研究尚未见报道。本研究利用植物显微技术,对地锦的根、茎、叶的结构及乳汁管的分布和发育进行了系统研究,以期为地锦的开发利用提供理论依据。
1 材料与方法
1.1材料、仪器与试剂
供研究的地锦群落1 300 株左右,分布在约1 000 m2的建设取土区域内。该区域位于南阳师范学院校园内(N:32°976′,E:112°496′,H:262 m)。研究样地年平均气温14.4~15.7 ℃,年均降雨量约1 000 mm。使用的仪器设备有体视显微镜(NIKON)、制刀机(Leica EM KMR3)、轮转切片机(Leica RM 2265)、生物组织摊片机(YT-7F)、倒置显微镜(NIKON TE2000U)。数据分析使用Excel软件。使用的试剂主要有:戊二醛、磷酸二氢钠、磷酸氢二钠、乙醇、环氧丙烷、环氧树脂(EPON 812)、甲苯胺蓝-O、加拿大树胶。
1.2方法
1.2.1 材料选择 随机选取成熟且健康的地锦植株10株,对其结构特征进行观测,主要记录地锦的器官大小和生长规律。乳汁管直径以同一器官的10个横切面为基础,每一横切面上乳汁管直径取长轴值计算平均值。
1.2.2 营养器官的制片 选取健康植株的根、茎、叶,蒸馏水冲洗干净。用双面刀片截取长度2~3 mm的组织块作为样品。样品放入2.5%戊二醛磷酸固定液(0.1 mol磷酸缓冲液)中,真空泵抽气30 min,室温下固定12~24 h。然后用0.1 mol磷酸缓冲液(pH值6.8)冲洗3次,每次30 min。再经体积分数为30%、50%、70%、85%、95%、100%乙醇脱水,每个梯度处理20 min。将样品移入体积分数为50%乙醇+体积分数为50%环氧丙烷的混合液中浸泡30 min后,再用100%的环氧丙烷冲洗3次,每次20 min。此后样品置于体积分数为50%环氧丙烷+体积分数为50%环氧树脂的混合液中过渡4 h,再经100%的环氧树脂处理12 h。最后,样品置入磨具内,环氧树脂包埋,60 ℃聚合24 h以上。取聚合良好的样品,在轮转切片机上修块、切片,切片厚度为2 μm。切片经甲苯胺蓝-O染色,树脂封片,显微镜下观察并拍照[4]。
1.2.3 叶表皮制片 取成熟的地锦叶片,放入装有1 mL体积分数75%酒精的1.5 mL EP管中,煮沸15 min后蒸馏水冷却清洗,然后放在载玻片上,剥离上、下表皮,甘油封片,显微镜下观察拍照[5]。
2 结果与分析
2.1根的结构
2.1.1 根的初生结构 地锦根的初生结构包括表皮、皮层和维管柱(图1-1、图1-7、图1-8)。主根的表皮细胞1层。皮层包括外皮层、皮层薄壁细胞和内皮层。外皮层细胞一层,排列紧密;皮层薄壁细胞3~4层,近圆形,相对较大,排列疏松,细胞间隙大,所占主根横切面的比例较大;内皮层细胞一层,排列紧密,有凯氏带加厚。维管柱的最外一层是维管柱鞘;在维管柱内部,初生木质部与初生韧皮部相间排列,原生韧皮部内侧分化出韧皮薄壁细胞、筛管和伴胞后构成后生韧皮部,原生木质部内侧分化出口径较大的导管和木薄壁细胞后组成后生木质部(图1-2)。地锦根的初生结构的发育方式是外始式,木质部为三原型。侧根比主根细小(图1-7),直径约(149.1±7.03)μm,维管柱直径约(93.36±3.55)μm,导管直径约(12.22±3.69)μm;除有主根和侧根外,地锦的茎节处还产生不定根(图1-8),直径约(242.32±11.42)μm,皮层薄壁细胞所占比例较大,细胞形状呈多边形,胞间隙大,厚度约(135.76±15.39)μm,维管柱直径约(99.13±8.12)μm。
2.1.2 根的次生结构 主根的次生结构由外向内由周皮、次生韧皮部、维管形成层和次生木质部组成(图1-3)。由于根的加粗,使表皮和皮层撑破,木栓形成层形成新的保护组织周皮,包括木栓层、木栓形成层和栓内层(图1-6)。次生结构生长初期,韧皮部内侧弧形轮廓的薄壁细胞恢复分裂能力向外扩展推移,与初生木质部脊处具分裂能力的中柱鞘细胞相连,形成波状维管形成层环。此后,维管形成层进行平周分裂,向内产生次生木质部、向外产生次生韧皮部。次生木质部数量较多,所占体积也相对较大,次生木质部导管直径较大,管壁厚约(2.28±0.19)μm,管径约(19.4±4.57)μm,导管为网纹导管或环纹导管(图1-4),多分布于木射线之间。维管射线由2~3列细胞构成。不定根中没有次生结构。

ep.表皮;c.皮层;v.维管柱;p.中柱鞘;po.原生木质部;me.后生木质部;cp.皮层薄壁细胞;pe.周皮;ex.外皮层;en.内皮层;pp.初生韧皮部;px.初生木质部;vc.维管形成层;pg.木栓形成层;pl.栓内层;ck.木栓层;pr.髓射线;pa.韧皮射线;xr.木射线;sx.次生木质部;sp.次生韧皮部;ve.导管;av.环纹导管;rv.网纹导管;la.乳汁管;sa.气孔器;ue.上表皮;le.下表皮;pt.栅栏组织;st.海绵组织;vb.维管束鞘细胞;fc.束中形成层;ic.束间形成层;pi.髓;cr.晶体(标尺=50 μm),下同。ep.epidermis; c.cortex; v.vascular bundle; p.pericycle; po.protoxylem; me.metaxylem; cp.cortical parenchyma; pe.periderm; ex.exodermis; en.endodermis; pp.primary phloem; px.primary xylem; vc.vascular cambium; pg.phellogen; pl.phelloderm; ck.cork; pr.pith ray; pa.phloem ray; xr.xylem ray; sx.secondary xylem; sp.secondary phloem; ve.vessel; av.annular vessel; rv.reticulate vessel; la.Laticifer; sa.stomatal apparatus; ue.upper epidermis; le.lower epidermis; pt.palisade tissues; st.spongy tissues; vb.vascular bundle sheath cells; fc.fascicular cambium;ic.interfascicular cambium;pi.pith;cr.crystal(Bar=50 μm).The same as below.1.主根的初生结构,示初生木质部和初生韧皮部的分化;2.主根的初生分生组织,示原生木质部和后生木质部的分化;3.主根的次生结构,示次生韧皮部、次生木质部和维管射线;4.主根导管的类型,示环纹导管和网纹导管的分化;5.乳汁管在主根次生结构中的分布;6.主根的纵切面,示周皮;7.侧根的横切面,示乳汁管的分布;8.不定根的横切面,示各部分比例;9.主根的纵切面,乳汁管的分布。1.The primary structure of taproot,showing the differentiation of primary phloem and primary xylem; 2.The section of primary meristem in the taproot,showing the differentiation of protoxylem and metaxylem; 3.The secondary structure of taproot,showing the secondary phloem,secondary xylem and vascular ray; 4.Vessels in taproot,showing the annular vessel and reticulate vessel; 5.The distribution of laticifers in the secondary structure of taproot; 6.Longitudinal section of the taproot,showing the periderm; 7.Cross section of the lateral root,showing the distribution of laticifers; 8.Cross section of the adventitious root,showing the ratio of each parts; 9.Longitudinal section of the taproot,showing the distribution of laticifers.图1 地锦根的解剖结构Fig.1 The anatomical structures of roots of E.humifusa
2.2茎的结构
2.2.1 茎的初生结构 地锦茎的初生结构由表皮、皮层、维管柱3部分组成(图2-1、图2-4),直径约(559.54±38.59)μm。
表皮细胞1层,排列紧密,厚度约(11.66±1.64)μm,其上分布有气孔器(图2-3);外有角质层,厚度约(1.56±0.25)μm。
皮层由1层厚角组织细胞和5~7层薄壁细胞构成,厚度约(107.84±13)μm,靠近韧皮部外侧的皮层薄壁细胞分化出环状排列的乳汁管。
维管柱直径约(294.47±15.02)μm,包括维管束、髓和髓射线。11个无限外韧维管束环状排列,由初生韧皮部、束中形成层、初生木质部组成;原生韧皮部内侧分化出伴胞、韧皮纤维和韧皮薄壁细胞组成后生韧皮部,共同组成初生韧皮部;原形成层分化出内始式发育的初生木质部,包括原生木质部和后生木质部,原生木质部外侧分化出导管和木薄壁细胞,组成后生木质部(图2-1),导管直径约(12.03±1.54)μm,管壁厚度约(2.52±0.67)μm。茎的中心为髓,薄壁细胞较大,呈圆形或多边形,排列疏松,胞间隙大。相邻维管束间由髓到皮层的3~5列薄壁细胞构成髓射线。
2.2.2 茎的次生结构 地锦的表皮、皮层、次生维管组织和髓构成茎的次生结构,次生维管组织中次生木质部包括导管、管胞、木薄壁组织和木纤维,次生韧皮部包括筛管、伴胞、韧皮纤维(图2-6)。次生部生长初期,初生木质部和初生韧皮部间的束中形成层和髓射线产生的束间形成层连接形成维管形成层环(图2-5)。维管形成层进行平周分裂,向内产生次生木质部、向外产生次生韧皮部,经连续切片观察,地锦茎维管形成层活动时间短,仅产生少量次生维管组织,不产生周皮。

1.茎初生结构,示初生木质部、初生韧皮部和维管形成层出现; 2.初生结构中的维管束,示乳汁管的分布;3.表皮中张开的气孔器;4.茎韧皮部外方乳汁管分布;5.维管形成层环和次生韧皮部附近乳汁管;6.茎的次生结构,示次生韧皮部和次生木质部
1.The primary structure of stem,showing the appearance of primary phloem,primary xylem and vascular cambium;2.Vascular bundle of the primary structure,showing the distribution of laticifers;3.An opening stomatal apparatus on the epidermis;4.The distribution of laticifers outside phloem;5.The ring of vascular cambium and the laticifers near secondary phloem;6.The secondary structure of stem,showing the secondary phloem and the secondary xylem
图2地锦茎的解剖结构
Fig.2TheanatomicalstructuresofstemofE.humifusa
2.3叶的结构
地锦叶无托叶,为不完全叶,单叶对生,矩圆形或肾状长圆形,长(4.2±0.56) mm,宽(3.54±0.3)mm,厚度(0.13±0.01)mm;叶尖钝圆、尖凹,基部偏斜,略呈渐圆形;主脉与叶缘红色,深红色叶柄较短,长(0.92±0.09)mm;叶面绿色,叶背淡红色,无表皮毛。地锦叶的横切结构由表皮、叶肉、叶脉组成(图3-1、图3-2),为异面型叶。
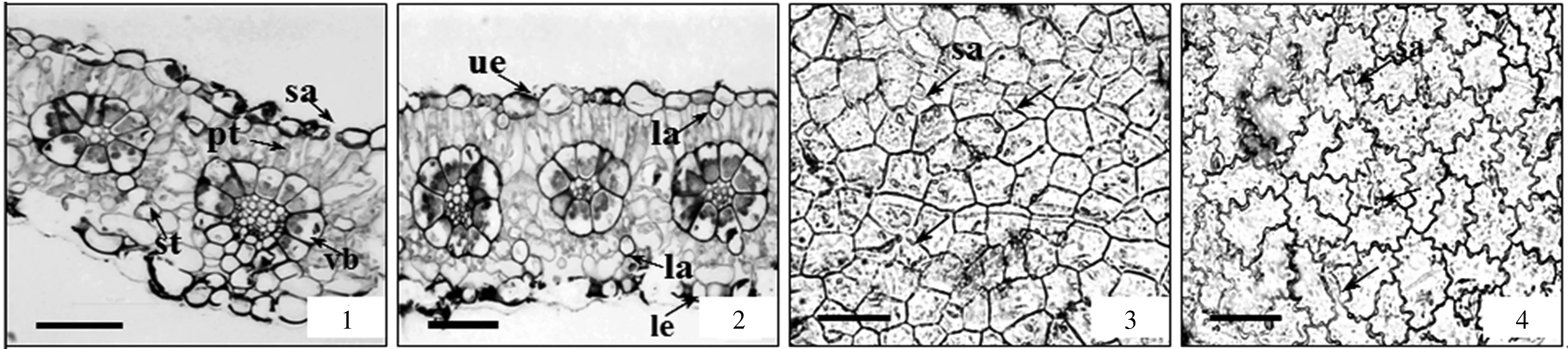
1.成熟叶的结构,示乳汁管的分布和花环状结构;2.异面叶,示栅栏组织和海绵组织;3.上表皮气孔的分布;4.下表皮气孔的分布
1.The structure of mature leaf,showing the distribution of laticifers and the kranz type bundle sheathes in mesophyll;2.Bifacial leaf,showing the palisade and spongy;3.The distribution of stomatal apparatus in the upper epidermis;4.The distribution of stomatal apparatus in the lower epidermis
图3地锦叶的解剖结构
Fig3TheanatomicalstructuresofleafofE.humifusa
上、下表皮细胞排列紧密,表面均有角质层,无胞间隙,不含叶绿体,厚度分别约为(13.15±4.83)μm和(13.5±2.43)μm。从叶表皮制片看,上、下表皮细胞的细胞壁相互嵌合,排列紧密,为不规则的多边形,上表皮垂周壁平直(图3-3),下表皮垂周壁深波状(图3-4)。上、下表皮单细胞面积分别约为(890.79±211.8)μm2和(1 355.85±326.03)μm2,上表皮细胞显著小于下表皮。气孔器由1对新月形的保卫细胞和气孔构成,属不规则型,上、下表皮气孔平均密度分别为344.14 个· mm-2和152.5 个· mm-2,单个气孔器周长分别约为38.79 μm和47.86 μm。
叶肉组织发达,由栅栏组织和海绵组织构成。栅栏组织细胞1层,呈长圆柱形,排列紧密,细胞间隙小,细胞长轴与表皮垂直,长度约(23.17±5.57)μm,有较多的叶绿体。海绵组织细胞形状不规则,排列疏松,细胞间隙较大,厚度约(41.67±19.72)μm。
叶脉为网状脉,叶脉维管束的木质部位于近轴面,韧皮部位于远轴面,为外韧维管束。主脉向下突起,直径约(125.8±6.05)μm;侧脉直径约(99.2±6.87)μm。主脉和侧脉周围均有由5~14个叶肉细胞构成的维管束鞘,形成碳四(C4)植物特有的花环式结构。
2.4乳汁管的结构
2.4.1 地锦乳汁管的分布 地锦主根的次生结构中,乳汁管主要分布在次生韧皮部的外侧(图1-5中la-2)、次生韧皮部的内部(图1-5中la-1)和皮层薄壁细胞中(图1-5中la-3),管腔形状不规则,直径大小差异较大。次生韧皮部外侧的乳汁管平均直径较小,约(17.2±2.59)μm。皮层中分布的乳汁管平均直径较大,约(20.09±2.38)μm。纵切面上(图1-9)可以看到乳汁管为无节乳汁管。侧根乳汁管分布位置与主根相同,形状相似,直径约(14.67±2.91)μm。不定根中未见乳汁管的分布。
在茎端分生组织中乳汁管已经开始分化。成熟的茎中,乳汁管主要分布在初生韧皮部(图2-2中la-1)和次生韧皮部的外侧(图2-6),大部分散生,也有少数聚生(图2-2中la-2),初生结构中乳汁管直径约(28.43±2.93)μm,次生结构中乳汁管直径约(61.54±8.8)μm。髓和髓射线中也偶有乳汁管分布(图2-2中la-3)。由于茎的增粗,乳汁管由多边近圆形挤压形成椭圆形(图2-6)。此外,茎中乳汁管内含有较多颗粒状晶体物质(图2-4、图4-2)。
叶中乳汁管主要集中在栅栏组织外侧和海绵组织内部(图3-2),在叶脉的韧皮部外侧也有分布(图3-1)。叶中的乳汁管直径大小差异不大,直径约为(11.17±0.82)μm。
2.4.2 地锦乳汁管的发育 地锦乳汁管为无节乳汁管,在种子萌发早期由简单的原始细胞发育而来,侵入式生长,其不同生长发育时期大致分为I期(图4-1),II期(图4-2)和III期(图4-3)。I期特点为:细胞皱缩,细胞壁破裂;II期特点:原生质体游离状态,乳汁颗粒明显;III期特点:管腔清晰,管壁较厚。
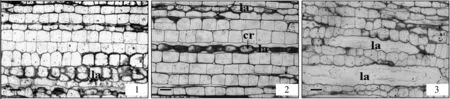
1.乳汁管发育I期;2.乳汁管发育II期;3.乳汁管发育III期1.Laticifer developmental stage I;2.Laticifer developmental stage II;3.Laticifer developmental stage III
3 结论
1)地锦根和茎的初生结构与次生结构的发育规律类似于双子叶植物根和茎的一般发育规律[6]。但是地锦茎的次生生长仅有维管形成层活动,木栓形成层则不活动,不产生周皮,这一结果与桔梗茎的次生生长特点相同[7]。
2)气孔是植物内、外气体交换的重要通道,地锦上表皮气孔密度大于下表皮,可能与下表皮细胞大上表皮细胞小有关。
3)碳四(C4)植物具有“花环”结构的维管束鞘细胞,是高光效的植物[8]。地锦叶片的维管束鞘发达,也具有“花环”结构,但地锦是否属于高光效的碳四(C4)植物,还需要从光合机理、酶的活性等方面开展进一步研究。
4)地锦各营养器官间乳汁管直径存在较大差异,根和茎中不同部位分布的乳汁管直径差异显著,而叶肉中的乳汁管直径则无较大差异,乳汁管直径大小依次为茎>主根>侧根>叶。和大戟属泽漆(EuphorbiahelioscopiaL.)的乳汁管直径大小根> 茎> 叶[9]的不同,和同属甘遂(EuphorbiakansuiLiou)的乳汁管直径大小茎>根>叶[10]相似。
5)大戟属植物多含有白色或黄色乳汁,主要贮存于乳汁管中[11]。地锦是全草入药[12],地锦乳汁管中含有乳汁及较多颗粒状晶体物质,有可能是其药用成分。地锦的药用成分是否主要集中在乳汁中,需要液相色谱技术做进一步检测。对地锦乳汁管的分布、发育与乳汁药用成分积累关系的研究,有利于确定其最佳采收时期和部位,提高药材产量和质量,是今后研究的主要方向。
[1] 中国科学院“中国植物志”编辑委员会.中国植物志:第44卷,第3册[M].北京:科学出版社,1997:49.
[2] 国家药典委员会.中华人民共和国药典[S].2010年版.北京:中国医药科技出版社,2010:118.
[3] 朱英.地锦草的研究进展[J].现代中药研究与实践,2003,17(5):62-63.
[4] 徐静文,廉文君,曹玉鑫,等.巴西橡胶树树皮环氧树脂组织切片及染色技术[J].热带生物学报,2014,5(2):161-165.
[5] 魏胜华,孟娜,柴瑞娟.地锦与斑地锦的显微鉴别研究[J].湖南农业科学,2012(1):42-43,49.
[6] 胡正海.植物解剖学[M].北京:高等教育出版社,2010,245-246.
[7] 谭玲玲,侯晓敏.桔梗营养器官的解剖学研究[J].青岛农业大学学报(自然科学版),2015,32(1):22-26.
[8] 巩玥,陈海苗,姜闯道,等.植物叶片解剖结构的量化及其在C4植物高粱中的应用[J].植物学报,2014,49(2):173-182.
[9] 魏博菲,蔡霞,胡正海.泽漆营养器官发育解剖学研究[J].西北植物学报,2011,31(8):1582- 1589.
[10] 李伟,蔡霞,王永建.甘遂乳汁管的解剖学及营养器官中二萜类化合物分布的研究[J].广西植物,2009,29(2):176-181.
[11] 李伟,蔡霞,胡正海,等.大戟属药用植物的研究进展[J].中国野生植物资源,2008,27(5):1-4,9.
[12] 柳润辉,王汉波,孔令义.地锦草化学成分的研究[J].中草药,2001,32(2):107-108.
(责任编辑:朱秀英)
AnatomicalstudyonvegetativeorgansandlaticifersofEuphorbiahumifusaWilld
LI Shuangjiang1,JIAO Zhujin2,XU Xiaomeng1,YAN Guanglan2,KAN Yunchao2,LI Chunqi1
(1.College of Life Science,Henan Agricultural University,Zhengzhou 450002,China; 2.Henan Provincial Key Laboratory of Funiu Mountain Insect Biology,Nanyang Normal University,Nanyang 473061,China)
The anatomic characteristics of vegetative organs,the distribution and development of laticifers ofEuphorbiahumifusaWilld were studied by using tissue sectioning and microscopic observation.The results showed that the primary xylem ofE.humifusaroot was triarch.The secondary structure of root consists of periderm,secondary phloem,secondary xylem and vascular cambium.The vascular bundle diameter(V)/root diameter(R)was 72.4%.The primary structure of stem consists of epidermis,cortex,vascular bundles and pith.No periderms existed in the secondary structure of stem.The leaf was bifacial leaf,and the stomata of upper epidermis were more than that of lower epidermis.There were kranz type bundle sheathes in mesophyll identical to that of the carbon 4 (C4) plant.The non-articulated laticifers ofE.humifusawere mainly distributed around the phloem of the taproot,lateral root,stem,and mesophyll.There were some granular crystal materials in the laticifers.The diameter of mature laticifers varied in vegetative organs,whose descending order lists as stem,taproot,lateral root and leaf.
EuphorbiahumifusaWilld;vegetative organs;anatomical structure;laticifers
Q 944.53
:A
2015-11-17
国家自然科学基金项目(31172158)
李双江(1988-),男,河南驻马店人,硕士研究生,主要从事植物解剖结构方面的研究。
李春奇(1963-),男,河南鲁山人,教授,博士。
1000-2340(2016)03-0383-06
