不同花椒品种光合特性比较研究
2016-09-23杨途熙魏安智李立新冯世静欧万发
陈 旅,杨途熙,魏安智,李立新,冯世静,欧万发,崔 丹
(西北农林科技大学 林学院,陕西 杨凌 712100)
不同花椒品种光合特性比较研究
陈旅,杨途熙,魏安智,李立新,冯世静,欧万发,崔丹
(西北农林科技大学 林学院,陕西 杨凌712100)
为筛选和推广高光效基因型花椒优良品种,寻求花椒品种最佳的光合效率管理模式,提高花椒产量和质量,在自然条件下,用Li-6400XT光合测定仪对8种不同产地4年生花椒树进行光合生理特性测定并分析。结果表明:8种花椒净光合速率日平均值大小顺序为党村无刺>狮子头>秦安一号>武都大红袍>无刺花椒>府谷花椒>韩城大红袍>凤县大红袍;8种花椒的净光合速率日变化曲线呈单峰型和双峰型2种类型;相关分析表明,8种花椒叶片的净光合速率与其蒸腾速率、气孔导度、水分利用率呈显著或极显著正相关,与胞间CO2浓度呈极显著负相关(P<0.01);对8种花椒进行光响应、CO2响应曲线拟合,得出8种花椒光的补偿点为36.30~102.76 μmol/(m2·s),饱和点为332.41~467.89 μmol/(m2·s);CO2补偿点为47.46~76.41 μmol/(m2·s),饱和点为698.887~1 509.000 μmol/(m2·s)。结果可为以后栽培管理、推广花椒提供参考依据。
花椒;光合作用;净光和速率;蒸腾速率;气孔导度
光合作用是植物合成有机物的过程,不同植物有着其独特的光适应特性,对不同植物进行光合作用方面的测定,能够分析不同环境因素对植物生长的影响并在植物规模栽培、引种推广时作为理论参考对植物进行合理的管理。近年来,国内外在不同植物光合作用上的研究报道很多,果树光合作用的日变化曲线表现为2种类型,其中一种为双峰曲线,有明显的 “午休”现象,一种为单峰[1]。但果树光合作用在具有相似性的同时,又因树种、品种、树龄、环境、试验条件和试验方法等因素的不同而呈现出一定的差异性。不同果树品种的补偿点和饱和点不同,光-光合响应曲线也不尽相同,同一树种在不同测定时期、不同树龄、不同季节的测定也存在明显差异。光是光合作用中光能的唯一来源,CO2则是光合作用的基本原料[2]。因此,了解植物光响应曲线和CO2响应曲线,可以得到植物的一系列光合参数。
花椒(Zanthoxylumbungeanum)为芸香科花椒属植物,全世界约250种,分类学上一般将花椒属分为花椒亚属和崖椒亚属,分布在亚洲、非洲、北美洲的热带和亚热带地区[3],是重要的食用调味品、中药材、香料及木本油料树种之一,也是一种经济效益和生态效益兼顾的优良树种[4];我国约有花椒属植物45种,主要品种有花椒及野花椒、竹叶花椒、青花椒[5]。目前,国内外对花椒的研究主要集中在有效成分提取、药用价值、栽培技术、分子遗传学等[6-15]方面,而对花椒光合作用研究较少。刘玲等[16]在自然条件下,对10个不同产地1年生花椒实生苗进行光合特性研究,结果表明,不同花椒的净光合速率日变化曲线和蒸腾速率日变化曲线均呈单峰型和双峰型2种类型。李明等[17]同时发现影响花椒净光合速率值的主要生态生理指标大小依次为:光合有效辐射>蒸腾速率>气孔导度>胞间CO2浓度。
但是,近年来我国在花椒的种植和生产中存在着品种混杂不清、管理技术粗放、优良易推广品种少、产品加工落后等问题,体现了我国在花椒研究工作上远远落后于其他经济树种[18]。因此,对花椒优良品种进行筛选、推广等工作是发展花椒产业的当务之急。本试验拟通过对不同种质资源花椒的光合特性、光响应曲线、CO2响应曲线进行研究比较,同时利用光响应曲线、CO2响应曲线各参数进行系统聚类,将当地8种花椒在光合作用的基础上进行大体归类;以期为筛选和利用高光效基因型的花椒优良品种、寻求最佳的光合效率管理模式、提高花椒产量和质量提供理论依据,能在引种、推广花椒种植过程达到最高经济效益。
1 材料和方法
1.1试验材料
选用凤县大红袍、韩城大红袍、武都大红袍、秦安1号、无刺花椒、府谷花椒、韩城狮子头、韩城党村花椒,水肥管理基本一致。
1.2试验地
试验设在陕西凤县白石铺村国家花椒工程研究中心4年生花椒种质试验园进行,花椒栽植行距为4 m×4 m,常规管理。
1.3光合指标测定
日变化:田间自然条件下,利用Li-6400分析仪闭路测定。选择晴天,从8:00-18:00(天微黑)每隔2 h测定一次光合生理指标,每株树取东南西北4片叶子,每个品种选3株,取平均值,作日变化曲线。
瞬时光合生理指标的测定方法:天气晴朗,光照条件充足,8:30-11:30光合有效辐射(PAR)设置为1 100 μmol/(m2·s),测定方法同日变化。
光响应曲线的测定:光合仪设置:PAR梯度设为0,50,100,200,400,600,800,1 000,1 200,1 400,1 600,1 800,2 000 μmol/(m2·s),CO2浓度为大气CO2浓度。
叶片选取及读数:每个品种选取长势相似3株,每种花椒树选取完全伸展的叶片3~4片,叶位、方向大体一致,在每个光强辐射梯度下读取数据3次。
CO2响应曲线的测定:光合仪设置:CO2浓度梯度设为0,20,50,100,200,300,400,600,800,1 000,1 200 μmol/mol,PAR设为1 100 μmol/(m2·s)。
叶片选取及读数:每个品种选取长势相似3株,每种花椒树选取完全伸展的叶片3~4片,叶位、方向大体一致,在每个光强辐射梯度下读取数据3次。
1.4数据处理
用Excel、SPSS 19.0统计分析软件对数据进行相关性分析,用Origin 8.0作图。水分利用率(WUE)的计算方法,WUE=净光合速率(Pn)/蒸腾速率(Tr)。表观量子效率(AQY)、光的补偿点(LCP)、光的饱和点(LSP)的计算方法,采用非直线双曲线方程[19]进行光响应曲线拟合,公式
其中,a为表观量子效率,Pnmax为最大净光合速率,k为光响应曲线曲角。再结合低PAR梯度下测得的Pn值建立直线回归方程,直线的斜率即为AQY,直线与横坐标轴的交点即为LCP,在Pn等于Pnmax处的PAR值就是LSP。
初始羧化效率(CE)、CO2的补偿点(CCP)、CO2的饱和点(CSP)的计算方法,采用直线双曲线方程[19]进行CO2响应曲线拟合,公式
Pn=CE·Amax·Ci/(CE·Ci+Amax)-Rp
其中,CE为初始羧化效率,Amax为饱和CO2浓度下光合速率,Ci为胞间CO2浓度,Rp为叶片的光呼吸速率。同理,用低Ci梯度下测得的Pn值建立直线回归方程,直线的斜率即为CE,直线与横坐标轴的交点即为CCP,在Pn等于Amax处的CO2浓度值就是CSP。
2 结果与分析
2.1瞬时(相同光强)8种花椒光合特性分析
光合速率(Pn)值的大小直接反映植物光合能力的强弱,二者呈正相关[20]。表1中,测定瞬时光合指标按瞬时光合速率均值由大到小排列,在相同光强下党村无刺花椒的瞬时净光合速率最大(均值为12.77 μmol/(m2·s)),凤县大红袍的净光合速率最小(均值为8.36 μmol/(m2·s))。党村无刺、狮子头与秦安一号之间没有显著性差异;但党村无刺、狮子头与其他品种间存在显著差异。气孔导度党村无刺最高,且与武都大红袍、府谷花椒、韩城大红袍、凤县大红袍存在极显著差异。胞间CO2浓度在P<0.01水平上没有明显差异;蒸腾速率党村无刺最大,它与秦安一号之间没有显著性差异,与其他品种存在极显著差异(P<0.01)。水分利用率上,武都大红袍与其他品种之间都没有极显著差异(P<0.01),且水分利用率均值最大。
2.2八种花椒的光合日变化
2.2.1净光合速率日变化净光合速率是植物光合作用能力最直接的体现。8种花椒的净光合速率日变化如图1所示,主要呈单峰型和双峰型。府谷花椒、狮子头、秦安一号、无刺花椒、武都大红袍、党村无刺6种花椒的净光合速率日变化曲线呈双峰曲线,第一个峰值出现在12:00左右,均值分别为12.82,14.82,13.22,13.87,12.53,16.23 μmol/(m2·s),第2个峰值出现在16:00左右,在14:00左右出现午休现象。凤县大红袍与韩城大红袍的净光合速率日变化呈单峰曲线,说明凤县大红袍与韩城大红袍没有午休现象,且净光合速率最大值出现在12:00左右,最大均值16.00,11.85 μmol/(m2·s),然后逐渐下降。
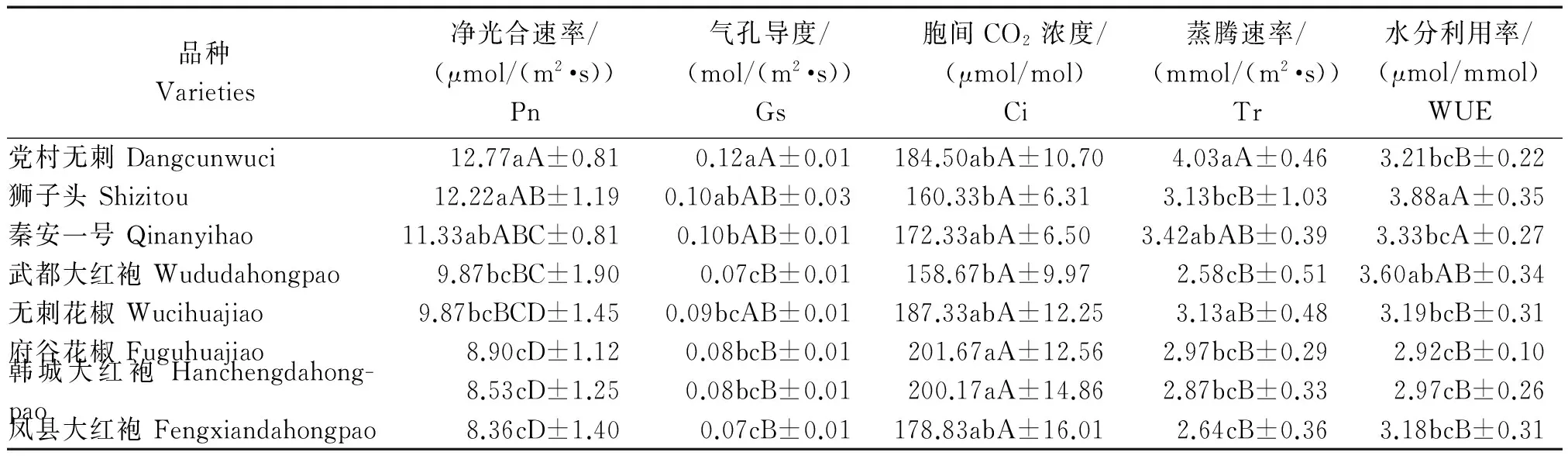
表1 不同花椒品种间瞬时光合参数的差异
注:小写英文字母表示在0.05水平上的显著性差异;大写英文字母则表示在0.01水平上的显著性差异。
Note:Lowercase English letters indicate significant difference at 0.05 level;Capital English letters indicate significant difference at 0.01 level.
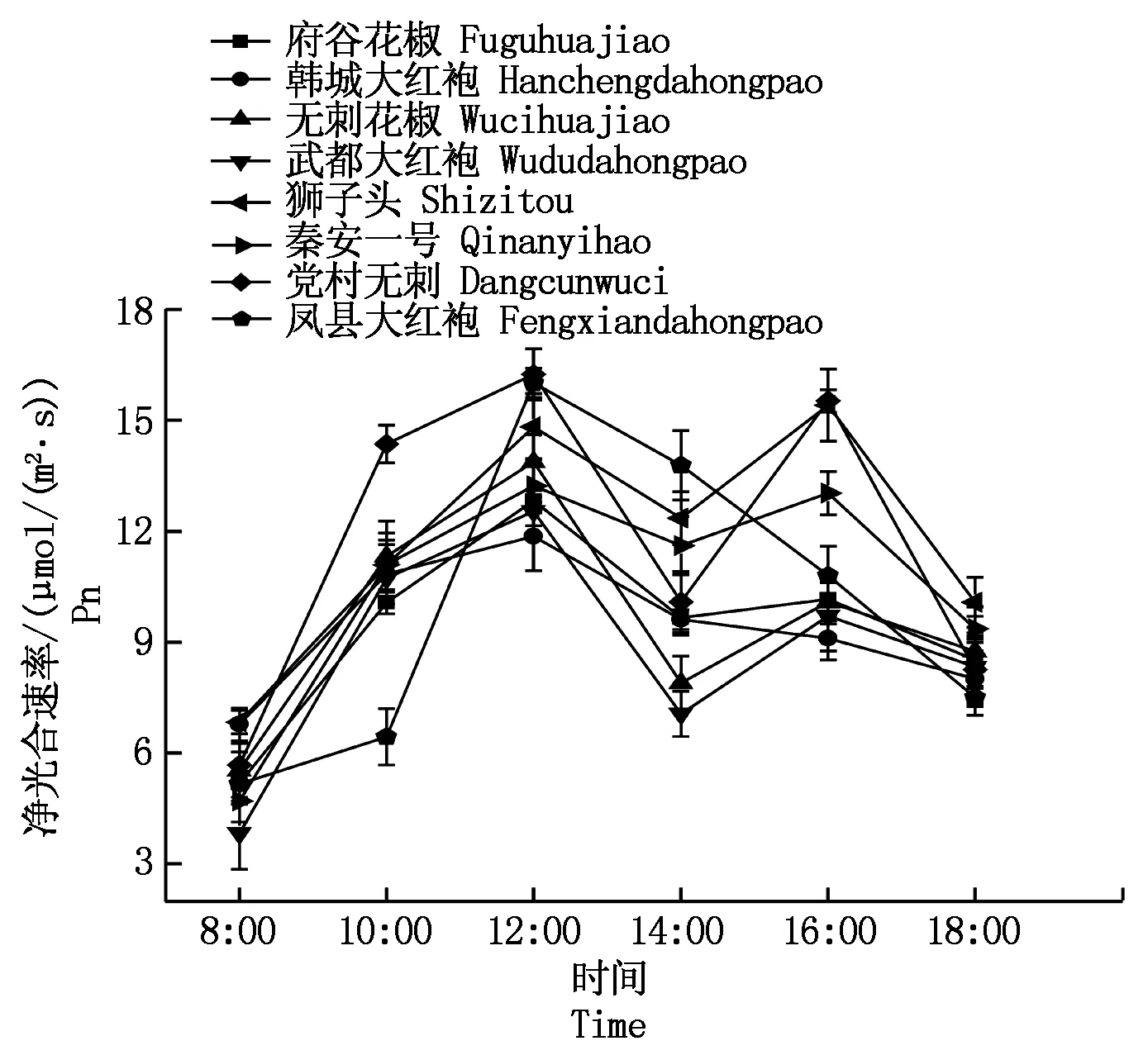
图1 不同品种花椒净光合速率日变化曲线
2.2.2气孔导度的日变化植物光合气孔导度表示植物叶片气孔的张开程度,气孔又是植物叶片与外界环境进行气体交换的主要通道。8种花椒气孔导度日变化如图2所示,气孔导度也存在单峰和双峰型。由图2可以看出,凤县大红袍、武都大红袍、狮子头、无刺花椒、韩城大红袍5种花椒为双峰型,且第1个峰值都大于第2个峰值。党村无刺、秦安一号、府谷花椒为单峰型,峰值出现在12:00左右。
2.2.3胞间CO2的日变化胞间CO2浓度(Ci)是光合生理生态研究中经常用到的一个参数。对8种花椒测得胞间CO2浓度如图3,在8:00时浓度最高,然后逐渐下降;其中韩城大红袍、府谷花椒、秦安一号最小值出现在12:00左右;无刺花椒、武都大红袍最小值出现在14:00左右;狮子头、党村无刺有2个谷,分别在12:00和16:00左右;而凤县大红袍最低值出现在16:00左右。总的来看,变化最大的为无刺花椒,变化趋于平稳的是狮子头。
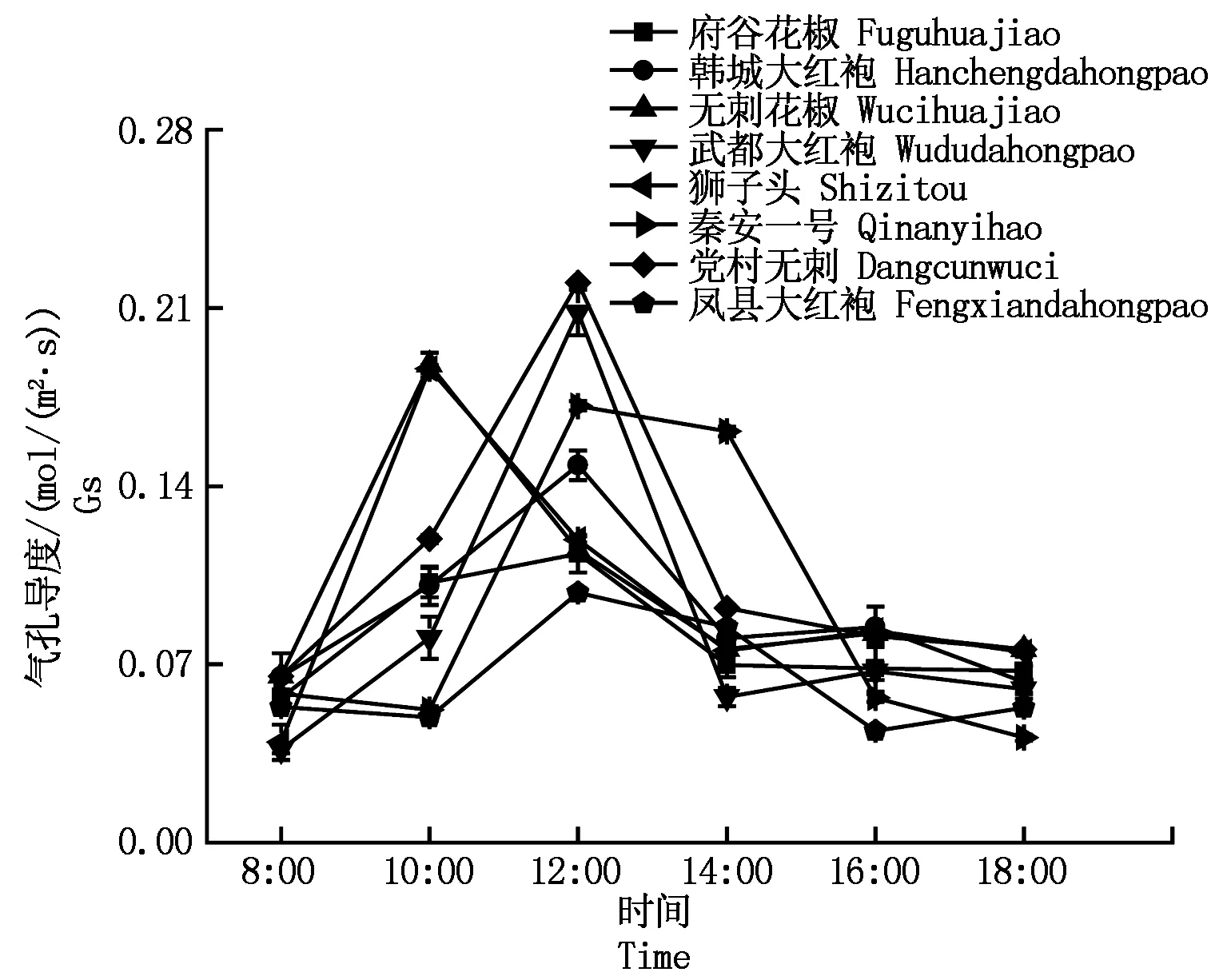
图2 不同品种花椒气孔导度日变化曲线
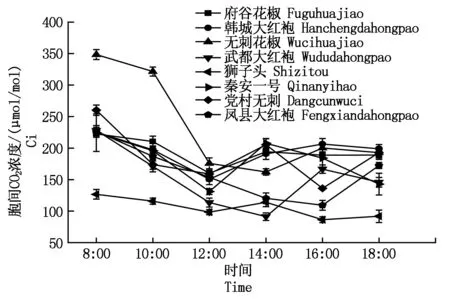
图3 不同品种花椒胞间CO2日变化曲线
2.2.4花椒蒸腾速率与水分利用率8种花椒的蒸腾速率和水分利用率日变化如图4-5所示。其中,韩城大红袍、无刺花椒、武都大红袍、狮子头、秦安一号5种花椒的蒸腾速率日变化趋势基本一致,呈单峰型,最大值出现在14:00左右;府谷花椒、党村无刺、凤县大红袍是双峰型,府谷花椒峰值出现在8:00和14:00;党村无刺峰值出现在10:00和14:00;凤县大红袍峰值出现在12:00和18:00。水分利用率的日变化除凤县大红袍外,其他与净光合速率日变化趋势一致。
2.3八种花椒光合作用的光响应曲线、CO2响应曲线比较
2.3.1八种花椒光响应曲线各参数间的相关性分析由表2可以看出,随着光合有效辐射(PAR)梯度的上升,8个不同产地花椒的净光合速率(Pn)值变化与PAR梯度之间存在显著的正相关,其中无刺花椒、韩城大红袍、凤县大红袍、秦安一号4种花椒为极显著的正相关;党村无刺的Gs与PAR梯度之间没有显著的相关性;Ci值的变化与PAR梯度之间存在显著的负相关,其中凤县大红袍、秦安一号为极显著负相关;Tr值的变化与PAR梯度之间存在极显著正相关;WUE值的变化除了凤县大红袍和党村无刺外,其他都与PAR梯度之间存在显著正相关。由表3可知,随着光梯度的上升,8个不同产地花椒的Gs值变化与Pn值变化之间存在显著正相关,其中无刺花椒、府谷花椒、凤县大红袍呈极显著正相关;Ci值的变化与Pn值变化之间存在极显著的负相关;WUE值的变化与Pn值变化之间除凤县大红袍外,均存在极显著的正相关,Tr值的变化与Pn值变化之间存在显著性正相关。
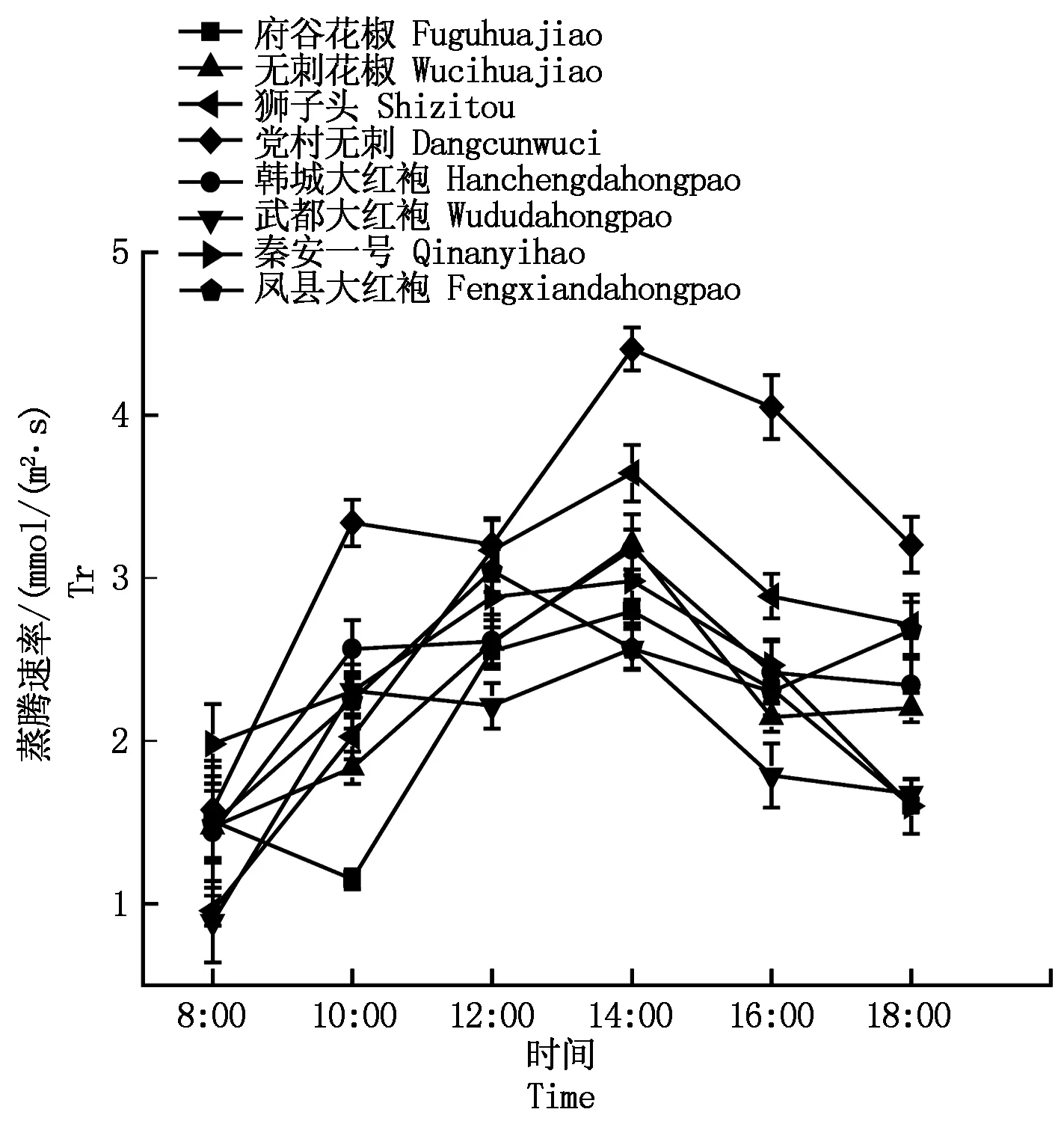
图4 不同品种花椒蒸腾速率日变化曲线
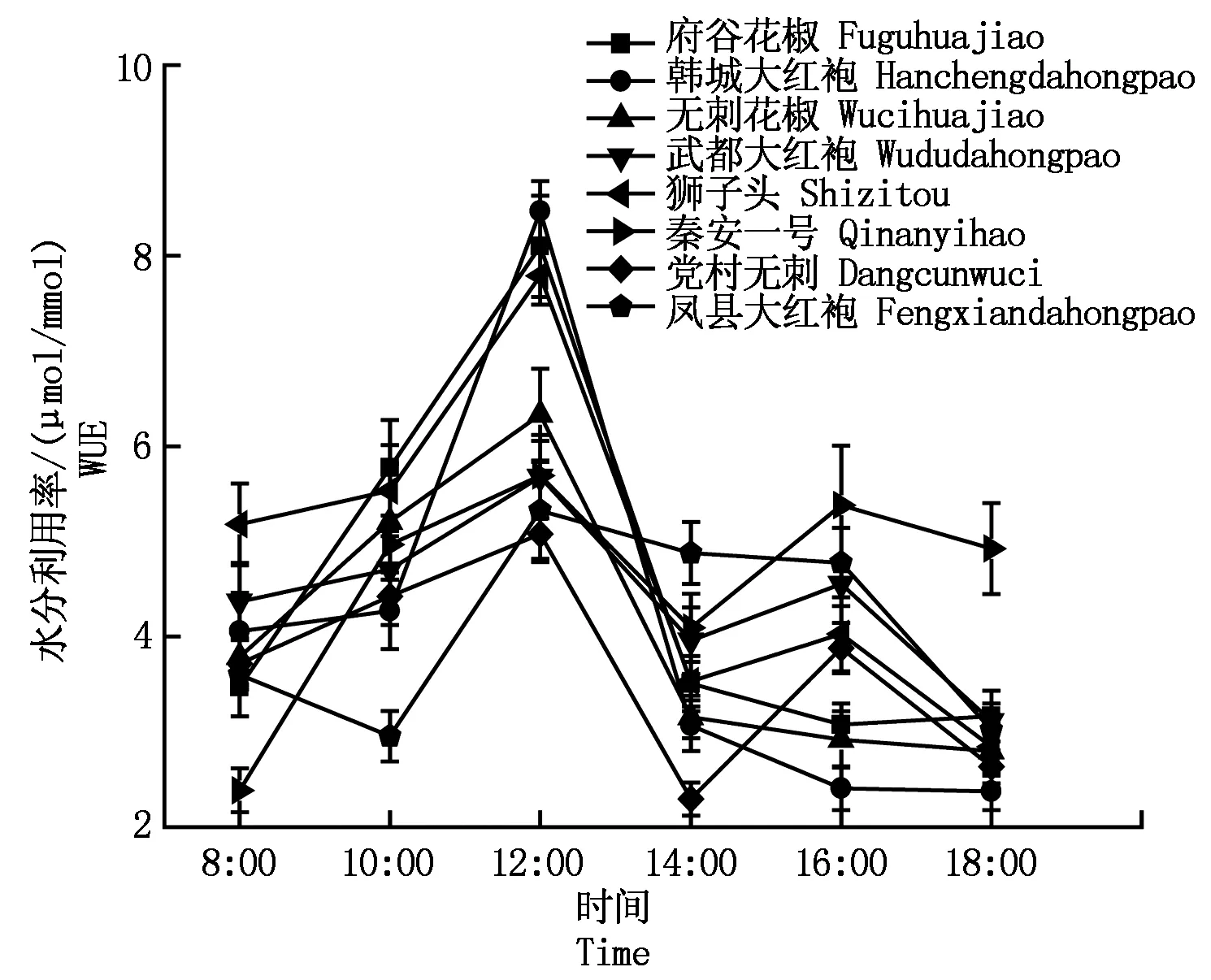
图5 不同品种花椒水分利用率日变化曲线
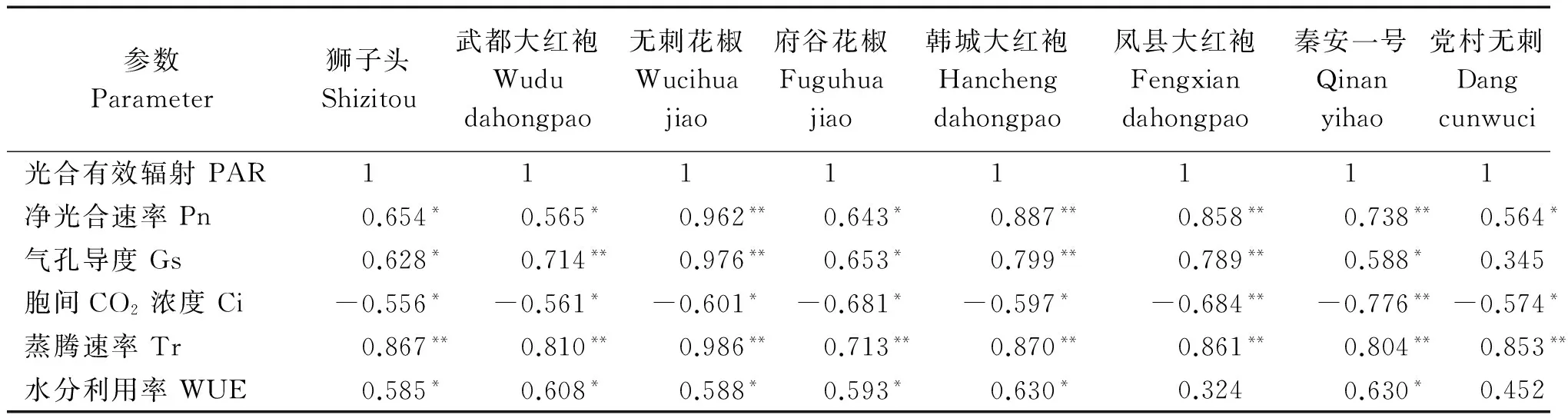
参数Parameter狮子头Shizitou武都大红袍Wududahongpao无刺花椒Wucihuajiao府谷花椒Fuguhuajiao韩城大红袍Hanchengdahongpao凤县大红袍Fengxiandahongpao秦安一号Qinanyihao党村无刺Dangcunwuci光合有效辐射PAR11111111净光合速率Pn0.654*0.565*0.962**0.643*0.887**0.858**0.738**0.564*气孔导度Gs0.628*0.714**0.976**0.653*0.799**0.789**0.588*0.345胞间CO2浓度Ci-0.556*-0.561*-0.601*-0.681*-0.597*-0.684**-0.776**-0.574*蒸腾速率Tr0.867**0.810**0.986**0.713**0.870**0.861**0.804**0.853**水分利用率WUE0.585*0.608*0.588*0.593*0.630*0.3240.630*0.452
注:*.在0.05水平上的显著性差异;**.在0.01水平上的极显著性差异。表3同。
Note:*.Correlation is significant at the 0.05 level;**.Correlation is significant at the 0.01 level.The same as Tab.3.
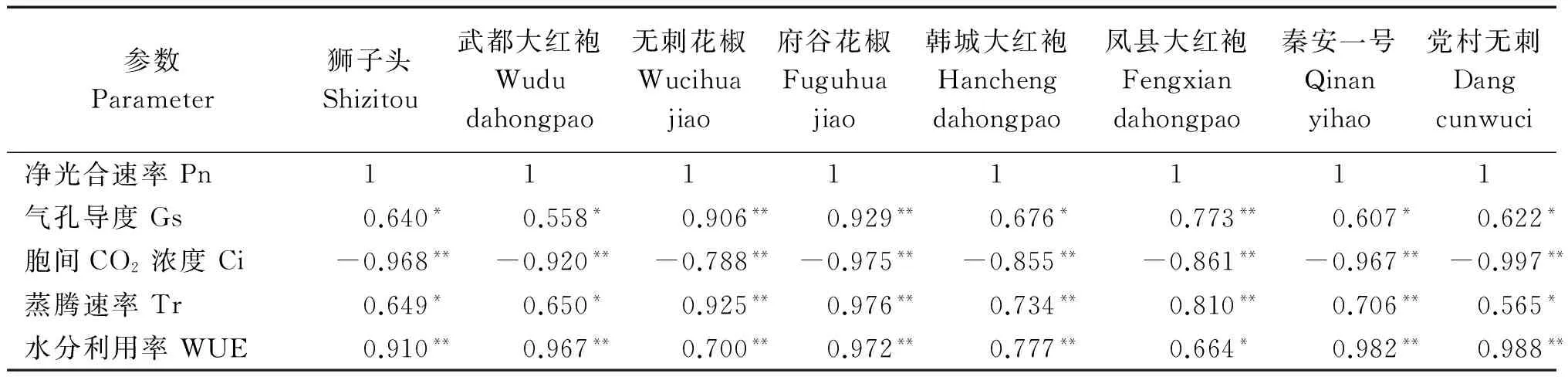
表3 不同PAR梯度下Pn与其他光合参数相关性
2.3.2八种花椒光响应曲线及AQY、LCP、LSP的比较由光合-光响应拟合曲线(图6)可知,8种花椒净光合速率(Pn)的大小随着光合有效辐射(PAR)的增大先大幅度增加,随后逐渐趋于平缓,增幅减小,达到一定值(光合饱和点)后Pn的大小不再随着PAR的增大而变化。其中无刺花椒对光的利用能力较强,党村无刺最低,其次是武都大红袍;其余各品种花椒对光的利用能力趋于相近。光是影响植物光合作用的重要环境因子,所以植物对光适应程度的强弱是其适应外界环境的重要表现[21]。
在自然条件下,植物的表观量子效率(AQY)为0.03~0.05[22];由表4可以看出,低PAR(<200 μmol/(m2·s))梯度下直线拟合得出的表观量子效率全部在上述范围之内。表观量子效率(AQY)能反映植物进行光合作用时对光的利用能力,尤其是植物对弱光的利用率,AQY值越高,说明此植物叶片对光能转化效率越高[23]。因此,表4可直观看出凤县大红袍、秦安一号表观量子效率最大,对光的利用率高;武都大红袍最低,对光的利用率低。通过非直角双曲线模型拟合结合低PAR梯度直线拟合得到光的补偿点(LCP)和光的饱和点(LSP),植物光补偿点(LCP)的高低能直接反映植物对弱光利用率的大小[24],是评价植物耐阴性的重要指标。由表4可以看出,凤县大红袍补偿点最低,说明凤县大红袍对弱光的利用率最强;党村无刺和府谷花椒的补偿点最高,说明它们对弱光的利用率最弱。LSP的高低表明其对高光强的忍受能力[25],韩城大红袍、无刺花椒和府谷花椒光的饱和点高,其中,韩城大红袍最高,说明韩城大红袍具有较强的耐高光强的能力。
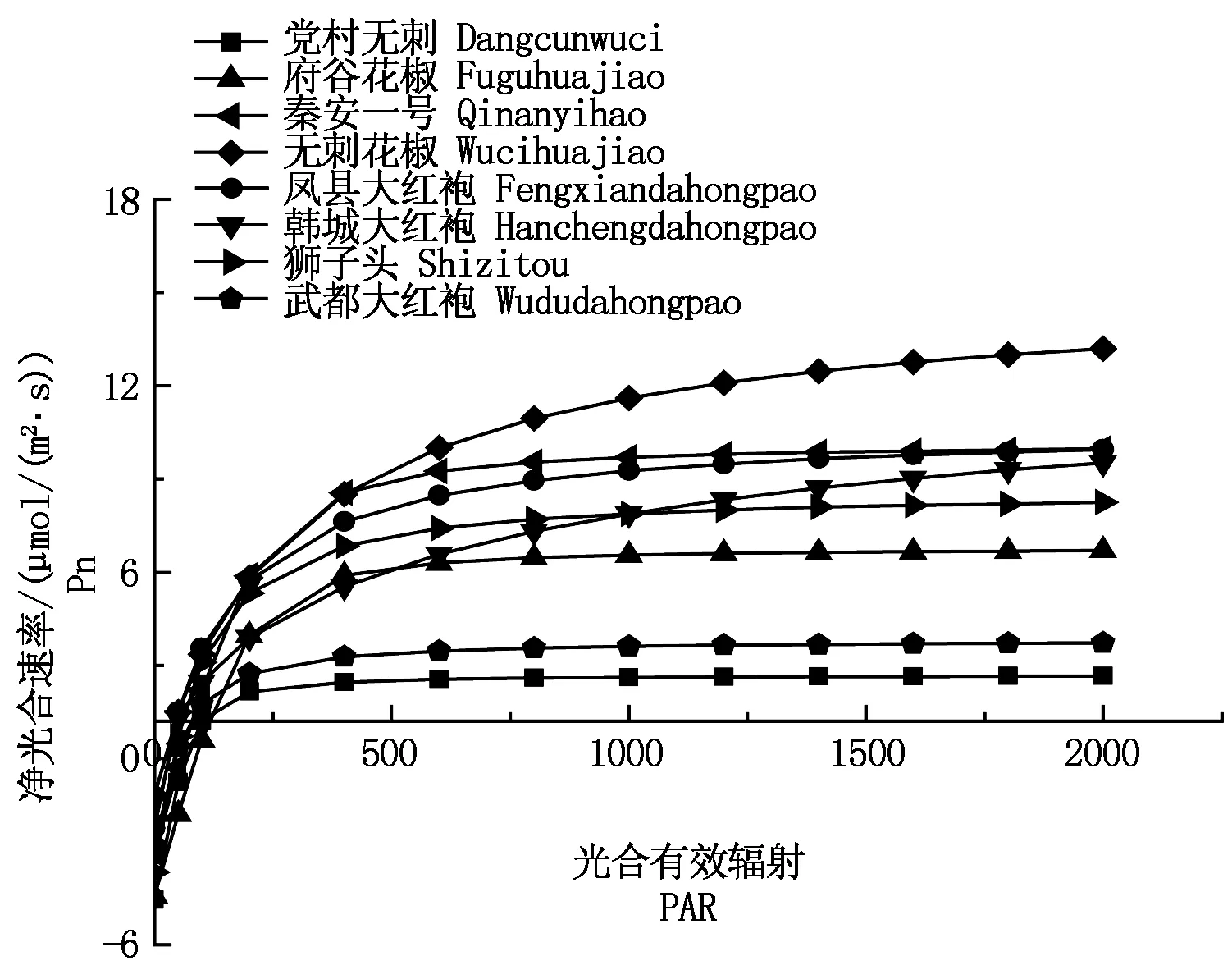
图6 不同品种花椒Pn的光响应曲线
2.3.3八种花椒CO2响应曲线及CE、CCP、CSP的比较8种花椒CO2响应曲线测定值如图7所示,可知净光合速率(Pn)对胞间CO2浓度(Ci)的响应趋势与光响应相似,即净光合速率(Pn)值随着胞间CO2浓度(Ci)的升高而增加。在低胞间CO2浓度(Ci<200 μmol/mol)时,各花椒品种Pn随Ci增加而迅速增加,之后Pn随Ci增加速率逐渐减缓。其中,党村无刺、凤县大红袍、秦安一号对CO2浓度的利用高于其他品种,府谷花椒、武都大红袍对CO2浓度的利用较低。
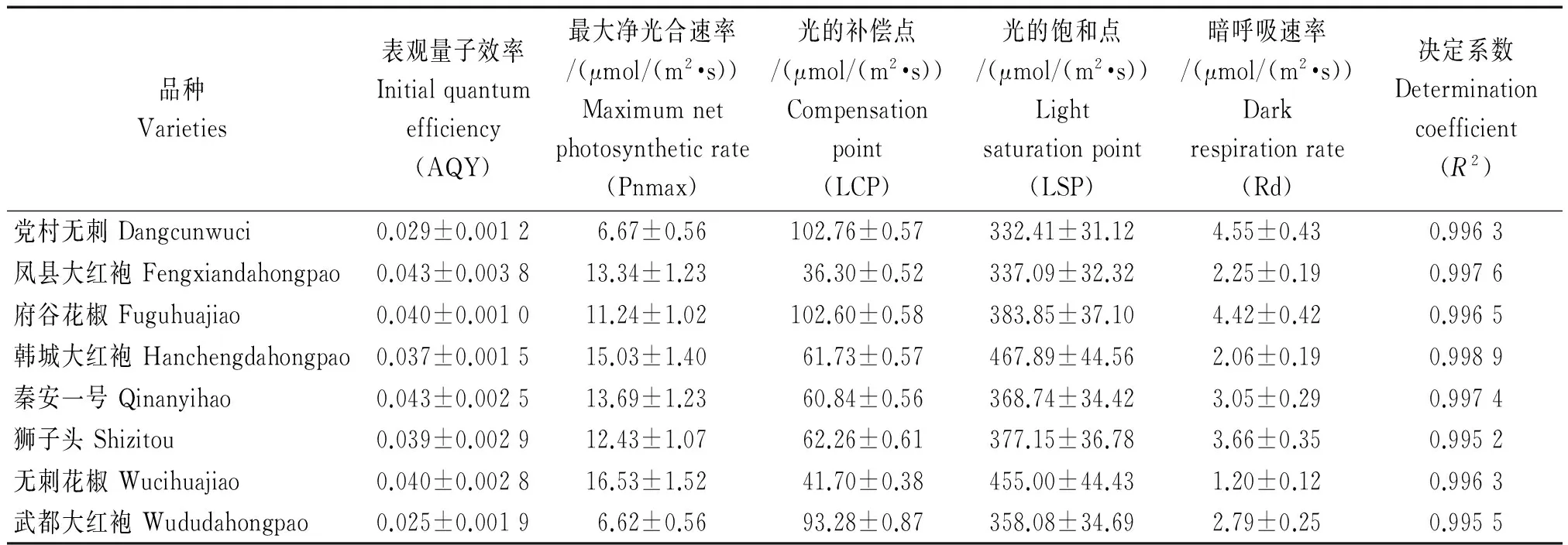
表4 不同产地花椒光响应曲线特征参数
利用直角双曲线模型拟合各花椒Pn-Ci响应曲线并计算出相应的特征参数(表5)。Amax 表示植物叶片的光合电子传递和磷酸化的活性[26],表5可知,党村无刺的Amax最高,说明其叶片光合电子传递和磷酸化的活性高于其他品种。植物光合作用初始羧化效率(CE)大小可以反映植物叶片中核酮糖-1,5-二磷酸羧化酶 (Rubisco)的活性大小及含量的高低[27],表中8种花椒凤县大红袍的CE最高,武都大红袍最低。CO2的补偿点(CCP)是植物光合同化作用与呼吸消耗相当时的CO2浓度,CO2的饱和点是植物利用高CO2浓度的能力。凤县大红袍和秦安一号的CCP最低,说明其容易适应低CO2浓度的外界环境;党村无刺的CSP最高,说明党村无刺对高CO2浓度的利用能力强。
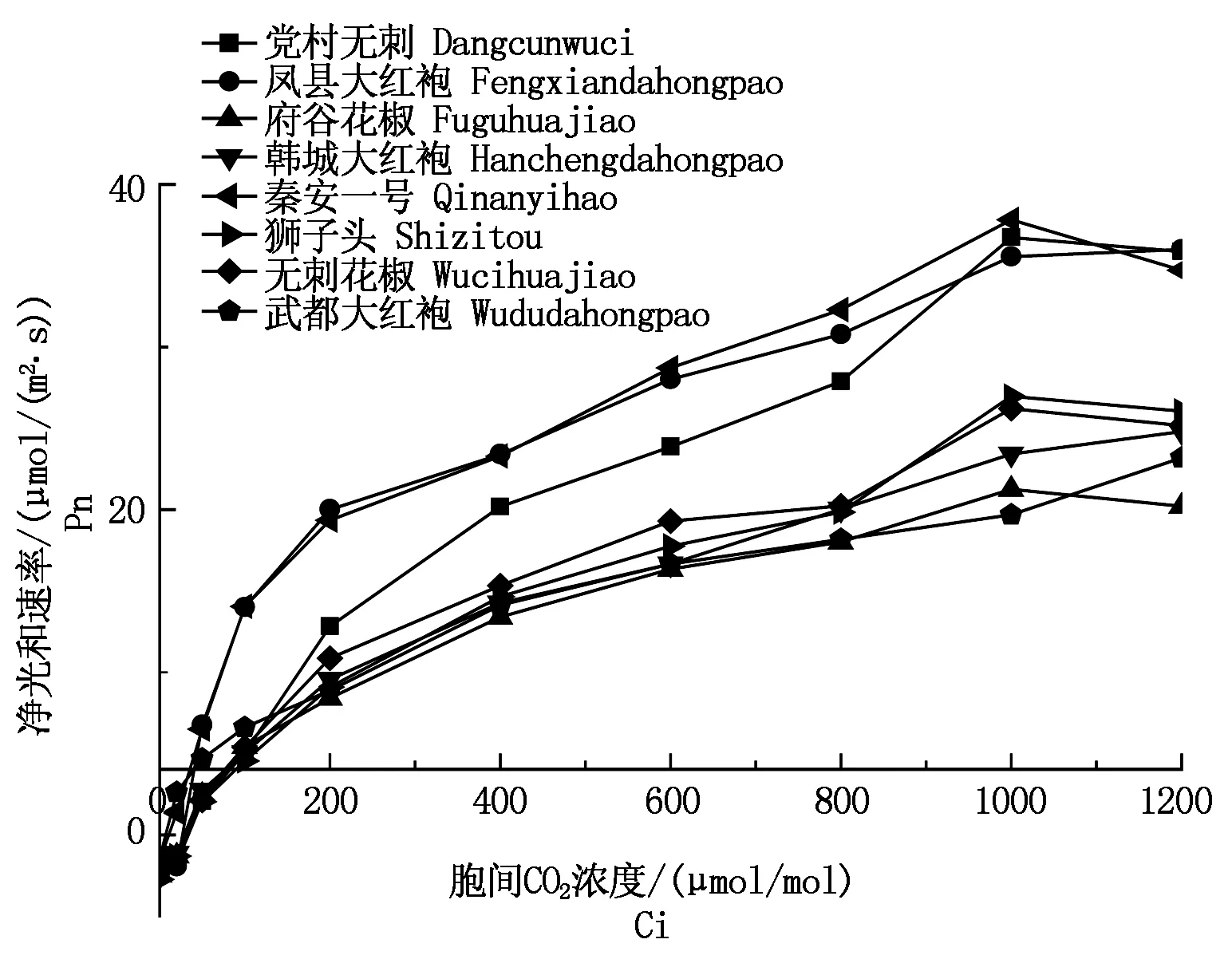
图7 不同品种花椒Pn-CO2响应曲线
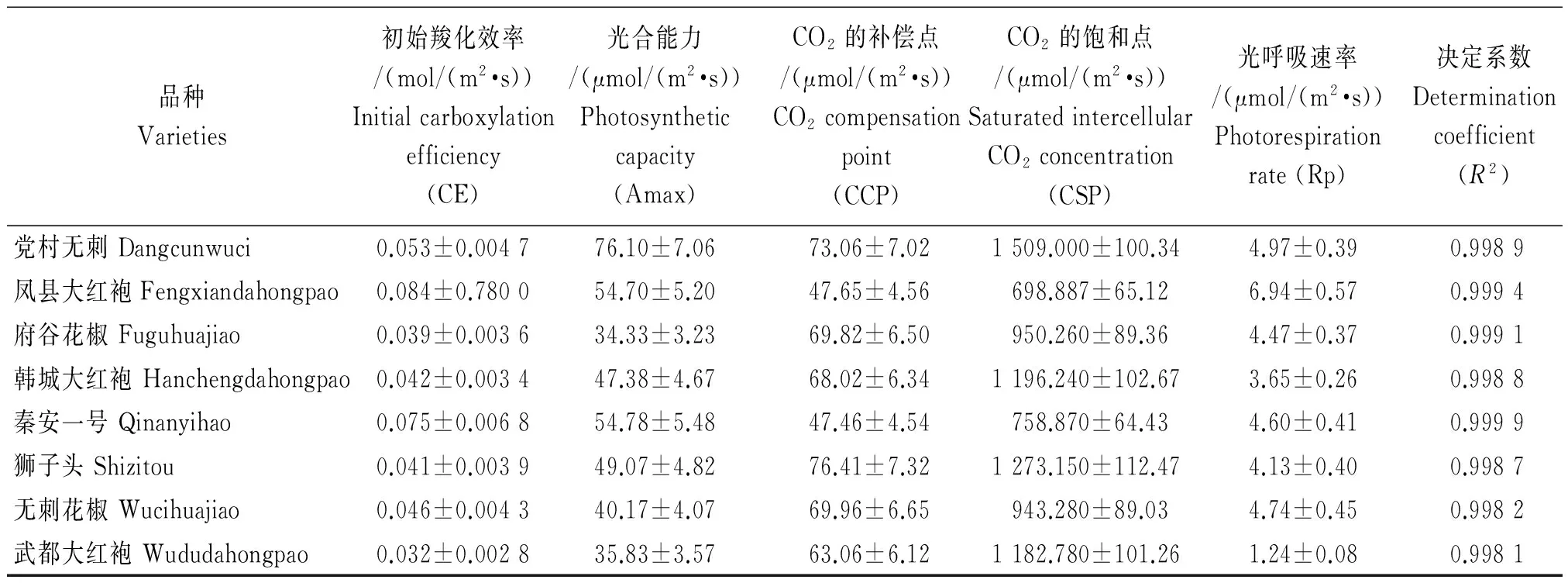
品种Varieties初始羧化效率/(mol/(m2·s))Initialcarboxylationefficiency(CE)光合能力/(μmol/(m2·s))Photosyntheticcapacity(Amax)CO2的补偿点/(μmol/(m2·s))CO2compensationpoint(CCP)CO2的饱和点/(μmol/(m2·s))SaturatedintercellularCO2concentration(CSP)光呼吸速率/(μmol/(m2·s))Photorespirationrate(Rp)决定系数Determinationcoefficient(R2)党村无刺Dangcunwuci0.053±0.004776.10±7.0673.06±7.021509.000±100.344.97±0.390.9989凤县大红袍Fengxiandahongpao0.084±0.780054.70±5.2047.65±4.56698.887±65.126.94±0.570.9994府谷花椒Fuguhuajiao0.039±0.003634.33±3.2369.82±6.50950.260±89.364.47±0.370.9991韩城大红袍Hanchengdahongpao0.042±0.003447.38±4.6768.02±6.341196.240±102.673.65±0.260.9988秦安一号Qinanyihao0.075±0.006854.78±5.4847.46±4.54758.870±64.434.60±0.410.9999狮子头Shizitou0.041±0.003949.07±4.8276.41±7.321273.150±112.474.13±0.400.9987无刺花椒Wucihuajiao0.046±0.004340.17±4.0769.96±6.65943.280±89.034.74±0.450.9982武都大红袍Wududahongpao0.032±0.002835.83±3.5763.06±6.121182.780±101.261.24±0.080.9981
2.3.4利用光响应、CO2响应各参数对8种花椒进行客观分类因近年来花椒种植品种混乱,且各种植区对花椒的叫法多样,孔繁玲[28]认为,用数学方法分析各指标间的相干性或不相干性的关系,可以得到客观划分类别的目的。因此采用系统聚类分析,以光响应、CO2响应各参数为因素聚类(图8),可以直观地将8种花椒分为2类,其中党村无刺、狮子头、韩城大红袍、武都大红袍为一类,其他4种为一类。
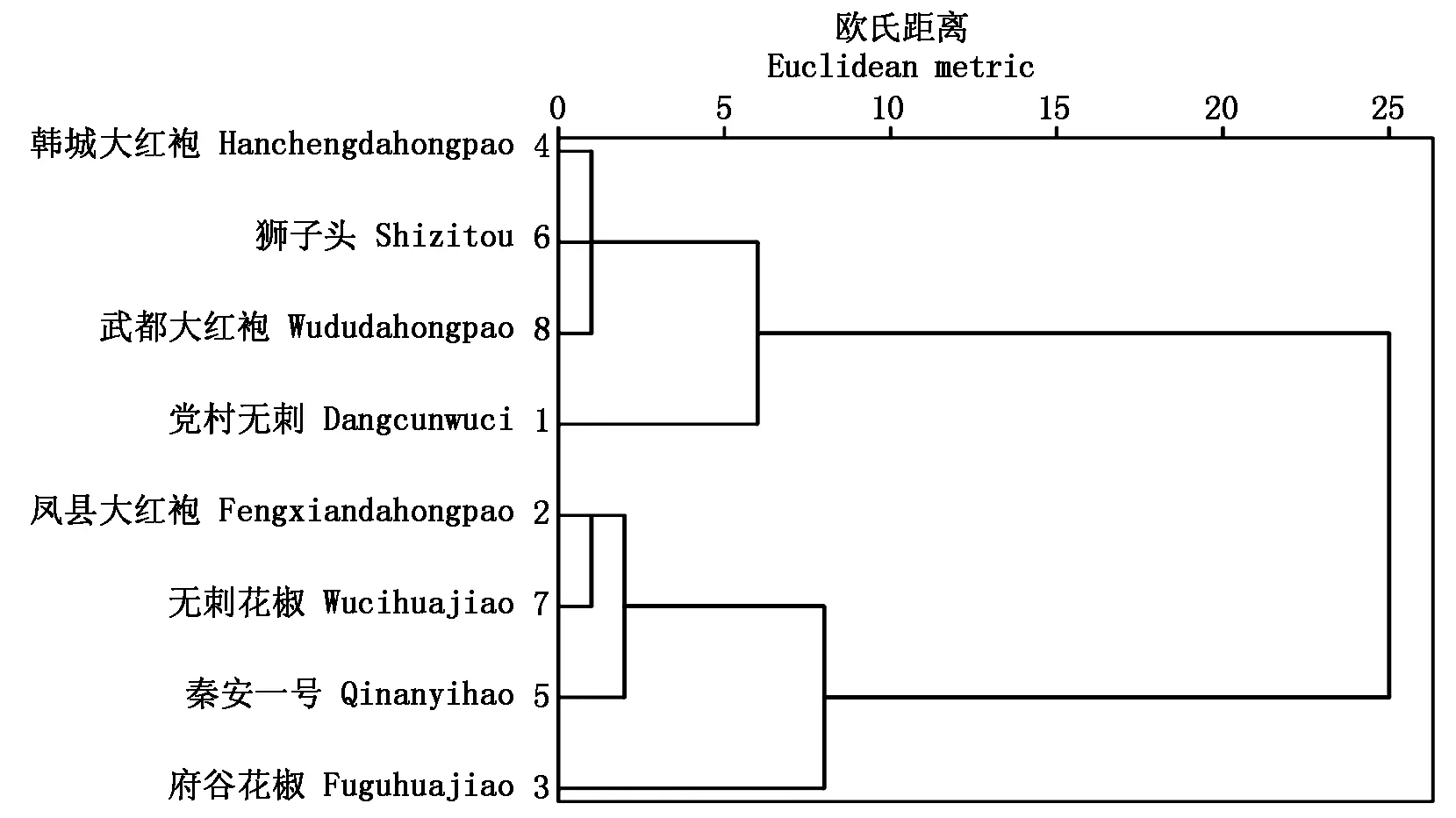
图8 不同品种花椒聚类图
3 讨论
瞬时光合速率(固定光强下)测得各参数,党村无刺Pn均值最大,Gs最大,Tr最大,说明在相同外部条件下,光强相同,党村无刺花椒的单位叶片进行光合作用的能力大于其他品种;凤县大红袍的净光合速率最低,说明当地环境和相同光强下其单位叶片的光合作用能力低;这可能与花椒品种长期对环境适应程度相关。
植物光合作用受很多外界环境因素的影响,如光照强度、环境中CO2浓度、外界温度、湿度和土壤水分等[29-32],且这些因素时刻都在变化。植物光合作用在一天中随着光强、温度、湿度等环境因子的变化称之为日变化。本试验测定了8种4年生花椒的日变化,结果表明,花椒光合作用、蒸腾过程都与光合有效辐射(PAR)变化相关。从早晨天亮开始,随着光合有效辐射(PAR)的增加,光合速率也增大,中午时达到一天中的最大值后再逐渐下降,天黑之前停止,形成单峰型日变化曲线,主要是凤县大红袍和韩城大红袍,这与刘玲等[16]测量结果一致。但其他6个品种在中午午休后会再达到一个峰值然后下降即为双峰型,即上午出现一个峰值,下午出现一个峰值,有“午休”现象。对于双峰曲线植物在午间降低(午休),可能受生理、生态、生化等多因素的影响,是一个相当复杂的难题[33]。8种花椒瞬时净光合速率平均值大小顺序为党村无刺>狮子头>秦安一号>武都大红袍>无刺花椒>府谷花椒>韩城大红袍>凤县大红袍。其中日变化呈双峰型的党村无刺和狮子头在“午休”时Pn和Tr平均值较高,说明党村无刺和狮子头对外界高温环境有一定的适应能力及主动调节能力,可以作为优良的高光效品种进行推广。
在本试验中,由不同PAR梯度下与其他光合参数的相关性分析,PAR与Pn之间存在显著性正相关(P<0.05);与Tr之间有极显著正相关(P<0.01);与Ci之间有显著负相关(P<0.05)。这说明,PAR的变化能引起光合作用的变化。不同PAR梯度下Pn与其他光合参数的相关性分析,Pn与Ci之间有极显著负相关(P<0.01);除凤县大红袍外,Pn与WUE之间存在极显著的正相关(P<0.01);Pn与Gs、Tr之间有显著性正相关(P<0.05)。这表明,8种花椒的净光合速率(Pn)的变化会影响到光合作用中其他因子的变化,他们相互作用共同影响着花椒的光合作用。
植物光合光响应曲线可以反映植物光合速率随外部光强变化而变化的规律。采用非直角双曲线拟合光响应曲线结合低PAR梯度(PAR<200 μmol/(m2·s))下直线拟合得到光响应各参数。光合光响应的各项指标能直接反映植物在逆境条件下的光合潜能、 光能利用率[34],AQY越低,说明植物光能利用率低,武都大红袍为8种花椒中利用光能最低品种,其次是党村无刺。花椒光响应曲线表明Pn 随PAR的增强而增大,当PAR超过一定数值(光的饱和点)后,Pn随PAR增加而降低或趋于平缓,即出现光合作用的光饱和现象,而生理上解释这种下降或趋于平缓的现象是因为光照过强而导致植物发生了光抑制作用。在相同的PAR下,无刺花椒的Pn均值大于其他品种,说明无刺花椒在高光条件下的光抑制程度较低,而党村无刺和武都大红袍最低,说明受高光条件的影响较大,光合效率低。LCP与LSP的大小反映植物对光照条件的要求和适应性强弱。8种花椒光的补偿点为36.30~102.76 μmol/(m2·s),饱和点为332.41~467.89 μmol/(m2·s)。一般来说,植物光的补偿点较低、光的饱和点较高,说明该植物对光环境的适应性较强;而补偿点较高、饱和点较低说明此植物对光的适应性较弱。本次试验研究中凤县大红袍的补偿点最低,但饱和点也低,韩城大红袍、无刺花椒补偿点低但饱和点高于其他品种,说明韩城大红袍、无刺花椒对环境的适应能力高于其他品种。
植物光合碳同化作用受植物体内Rubisco活性和1,5-二磷酸核酮糖(RuBP)再生速率的限制,一般认为植物CO2响应曲线的初始羧化效率(CE)与1,5-二磷酸核酮糖羧化酶(RuBPCase)的活性呈正相关[23]。8种花椒中凤县大红袍的CE最高,说明凤县大红袍叶片中RuBPCase活性高于其他品种。光响应拟合下的Pnmax均值都低于CO2响应拟合下的Amax,说明强光照条件下光合速率更大程度上受CO2供应的限制;其中党村无刺的CCP、CSP相对差值最大,说明其对CO2利用幅度最广。当植物进行光合作用时,外界CO2浓度达到植物CO2饱和点时,CO2浓度就不再是制约碳同化的主要因子,并且植物体内CO2受体的多少,即RuBP再生速率则成为限制光合作用的主要因素[35]。这可能就是当CO2浓度达到饱和点后,8种花椒光合速率趋于平缓或下降的原因。光合作用的光呼吸(Rp)一直以来被认为是一耗能过程,但郭连旺等[36]认为,光呼吸途径在消耗能量的同时,还能释放被3-磷酸甘油酸(PGA)结合的磷酸基团(Pi),这样就能缓解由于体内Pi暂时亏缺而引起的对电子传递和光合磷酸化过程的限制。8种花椒中凤县大红袍的光呼吸速率最大,但其饱和CO2浓度下Pn值相对较低;党村无刺的光呼吸速率居第二,且党村无刺饱和CO2浓度Pn值最大,说明党村无刺花椒在Pi亏缺时对电子传递和光合磷酸化过程的限制相对较小。
光合作用中CO2补偿点较低的植物,通常具有产量高、净光合速率高的特点;CO2补偿点越低的植物对外界环境中低CO2浓度有较高的利用能力,因此,育种学上将低CO2补偿点当作选育高产量品种的另一指标[23]。因此,对花椒光合各因子进行总体分析,若引种地区常年温度较高,需引进耐高温品种,党村无刺和狮子头相对于其他品种耐高温且对CO2的利用幅度大;若引种地区常年为干旱地区,可引进武都大红袍和凤县大红袍抗旱品种;韩城大红袍和无刺花椒对环境的适应力强,因此可以加以推广。
近年来,国内外对花椒品种分类的研究并不多见,且各种植区对花椒的命名也比较混乱,用系统聚类分析只能在数学方法上客观地将各种花椒区分开来,只能作为分类的一种参考,并不能准确地作为花椒分类的正确手段。因此,对于花椒的总体分类研究,希望能从分子生物学和遗传学角度来划分,各生理指标只能作为辅助参考依据。
[1]徐志防,罗广华,王爱国.光合作用的光抑制与光合器官的活性氧代谢[J].植物生理学通讯,1999,35(4):325.
[2]李宏梁,薛婷.花椒果皮的研究进展[J].中国调味品,2014(1):124-128,135.
[3]郭君难,田呈瑞.花椒开发利用的现状及前景分析[J].食品研究与开发,2008,29(8):167-170.
[4]周婷,蒲彪,姜欢笑.花椒麻味物质的研究进展[J].食品工业科技,2014(10):385-388,400.
[5]宋荣,曹亮,周佳民,等.花椒种质资源及其功能成分和生物学效应研究进展[J].湖南农业科学,2014(17):23-26,29.
[6]郑海星,李周岐,薛惠丹,等.花椒种质资源的RAPD分析[J].西北林学院学报,2011,26(2):96-100.
[7]朱晓慧,杨途熙,魏安智,等.无刺花椒嫁接愈合过程中相关生理指标的变化[J].西北林学院学报,2015,30(2):134-138.
[8]祝瑞雪,曾维才,赵志峰,等.汉源花椒精油的化学成分分析及其抑菌作用[J].食品科学,2011,32(17):85-88.
[9]姜太玲,吴红洋,董小华,等.花椒籽蛋白抗菌肽的起泡和乳化特性研究[J].核农学报,2015(12):2320-2326.
[10]吕小军.花椒花芽分化过程中的生理变化研究[M].杨凌:西北农林科技大学,2013.
[11]彭兴刚.汉源县花椒主要病虫害及防治措施[J].现代农业科技,2015(23):145-146.
[12]徐毅,阚建全.青花椒干燥过程叶绿素光降解原因初探[J].中国食品学报,2015(11):135-141.
[13]薛小辉.青花椒麻味成分的提取与分离[M].成都:四川农业大学,2013.
[14]杨立琛.花椒叶黄酮的微波提取及其成分分析[M].天津:天津商业大学,2013.
[15]张艳军.花椒黄酮和多酚含量及抗氧化活性研究[M].杨凌:西北农林科技大学,2013.
[16]刘玲,刘淑明,孙丙寅.不同产地花椒幼苗光合特性研究[J].西北农业学报,2009,18(3):160-165.
[17]李明,王有科,李捷,等.5个花椒品种光合特性的研究[J].甘肃农业大学学报,2010,45(6):128-132.
[18]毕君,王春荣,赵京献,等.北方花椒主产区种质资源考察报告[J].河北林果研究,2003,18(2):165-168.
[19]Thornley J H.Mathematical models in plant physiology[M].London:Academic Press,1976.
[20]刘怀年,邓晓建,李平.水稻品种资源光合速率研究[J].四川农业大学学报,2008,25(4):379-383.
[21]罗爱花,郭贤仕,柴守玺.水分胁迫对不同品种春小麦的水分利用率及生理指标的影响[J].干旱区研究,2008,25(5):683-689.
[22]李合生.现代植物生理学[M].北京:高等教育出版社,2002.
[23]董志新,韩清芳,贾志宽,等.不同苜蓿(MedicagosativaL.)品种光合速率对光和CO2浓度的响应特征[J].生态学报,2007,27(6):2272-2278.
[24]Sun C X,Qi H,Hao J J,et al.Single leaves photosynthetic characteristics of two insect-resistant transgenic cotton (GossypiumhirsutumL.) varieties in response to light[J].Photosynthetica,2009,47(3):399-408.
[25]Tartachnyk I I,Blanke M M.Effect of delayed fruit harvest on photosynthesis,transpiration and nutrient remobilization of Apple leaves[J].New Phytologist,2004,164(3):441-450.
[26]Coste S,Roggy J C,Imbert P,et al.Leaf photosynthetic traits of 14 tropical rain forest species in relation to leaf Nitrogen concentration and shade tolerance[J].Tree Physiology,2005,25(9):1127-1137.
[27]张彦敏,周广胜.植物叶片最大羧化速率及其对环境因子响应的研究进展[J].生态学报,2012,32(18):5907-5917.
[28]孔繁玲.植物数量遗传学[M].北京:中国农业大学出版社,2006.
[29]井艳丽,关德新,吴家兵,等.光合作用调控土壤呼吸研究进展[J].应用生态学报,2013,24(1):269-276.
[30]董伟欣,谢颍,刘明,等.短照诱导对小豆生长发育、光合和荧光参数的影响[J].华北农学报,2014,29(5):144-149.
[31]张德建,路战远,张向前,等.不同耕作条件下玉米光合特性的差异[J].华北农学报,2004,29(2):161-164.
[32]曹昌林,白文斌,史丽娟,等.不同培肥模式对小麦光合特性和干物质积累运转及产量的影响[J].山西农业科学,2014,42(7):663-666,671.
[33]赵黎明,李明,郑殿峰,等.水稻光合作用研究进展及其影响因素分析[J].北方水稻,2014,44(5):66-71.
[34]张力文,钟国成,张利,等.3种鼠尾草属植物光合作用-光响应特性研究[J].草业学报,2012,21(2):70-76.
[35]王海珍,韩路,徐雅丽,等.胡杨异形叶光合作用对光强与CO2浓度的响应[J].植物生态学报,2014(10):1099-1109.
[36]郭连旺,沈允钢.高等植物光合机构避免强光破坏的保护机制[J].植物生理学通讯,1996,32(1):1-8.
Research of the Photosynthetic Characteristics of Different Varieties of Chinese Prickly Ash
CHEN Lü,YANG Tuxi,WEI Anzhi,LI Lixin,FENG Shijing,OU Wanfa,CUI Dan
(College of Forestry,Northwest A&F University,Yangling712100,China)
In order to obtain for screening and utilization of high photosynthetic efficiency genotype of photosynthetic efficiency of Chinese prickly ash varieties,seeking the best management mode,improve the quality of Chinese prickly ash.In the fields,used the Li-6400XT photosynthetic apparatus to measure and analyzed eight different origin 4 years Chinese prickly ash trees for photosynthetic physiological characteristics.The results showed that the instantaneous net photosynthetic rateis as follows:Dangcunwuci>Shizitou>Qinanyihao>Wududahongpao>Wucihuajiao>Fuguhuajiao>Hanchengdahongpao>Fengxiandahongpao.The diurnal variation curves of net photosynthetic rate (Pn) of eight Chinese prickly ash in different producing areas were different,which showed two types,single peak curve and double peak curve.Correlation analysis showed that Chinese prickly ash leaf net photosynthetic rate and transpiration rate,cond,water use efficiency had a significant or highly significant positive correlation,leaf net photosynthetic rate and the Ci had a highly significant negative correlation (P<0.01).Though light response curves of Chinese prickly ash light compensation point of the range of 36.30-102.76 μmol/(m2·s),and light saturation point range from 332.41 to 467.89 μmol/(m2·s).and the CO2response curves of Chinese prickly ash CO2compensation point of the range of 47.46-76.41 μmol/(m2·s),and CO2saturation point range from 698.887 to 1 509.000 μmol/(m2·s).The experimental results can provide reference for later cultivation and management of Chinese prickly ash.
Chinese prickly ash;Photosynthesis;Net photosynthesis rate;Transpiration rate;Stomatal conductance
2016-04-12
国家林业局林业公益性行业科研专项(201304706)
陈旅(1989-),女,陕西旬阳人,在读硕士,主要从事林木遗传育种研究。
杨途熙(1963-),男,陕西杨凌人,副教授,硕士,主要从事林木遗传育种研究。
魏安智(1961-),男,陕西扶风人,教授,博士,博士生导师,主要从事林木遗传育种研究。
S573.01
A
1000-7091(2016)04-0153-09
10.7668/hbnxb.2016.04.025
